一株綠鷺源H9N2亞型禽流感病毒全基因組序列分析
2020-05-06 08:49:32段博芳曾邦權肖望成曾維歡黃翠琴代飛燕
野生動物學報 2020年2期
楊 勇 段博芳 曾邦權 肖望成 曾維歡 黃翠琴 代飛燕*
(1.福建省家畜傳染病防治與生物技術重點實驗室,龍巖學院生命科學學院,龍巖,360012;2.云南省高校獸醫公共衛生重點實驗室,云南農業大學動物醫學院,昆明,650201;3.云南省動物疫病預防控制中心,昆明,650201)
綠鷺(Butoridesstriata)為鷺科(Ardeidae)綠鷺屬,體形較小的涉禽,主要以魚為食。綠鷺部分種群遷徙,部分為留鳥;在中國長江以北繁殖的種群多為遷徙。通常在4月遷來北方繁殖地,9月離開繁殖地遷往南方越冬地[1]。野生水禽,被認為是低致病性禽流感病毒(LPAIVs)的主要宿主,它們攜帶的病毒可能會進化,成為高致病性的家禽或人畜共患病病原[2]可通過糞-口途徑或污染的水,在野生水禽之間或野生水禽與家禽之間傳播[3-4],通常,野鳥感染低致病性 AIVs 呈隱性感染,無明顯臨床癥狀[2]并隨其每年的遷徙而在不同國家地區間傳播。云南因其得天獨厚的自然環境與氣候條件成為許多野生候鳥的越冬地,大大增加了野生動物間疾病傳播的風險。
禽流感病毒屬正粘病毒科(Orthomyxoviridae)甲型流感病毒屬(InfluenzavirusA),流感病毒的基因組由8個單獨的負鏈片段組成,依次為PB2、PB1、PA、HA、NP、NA、M和NS,共編碼10種蛋白[5]。根據表面糖蛋白血凝素(HA)基因和神經氨酸酶(NA)基因,AIV 可以分為 16種 HA 亞型和 9種 NA 亞型幾乎所有的亞型組合都可以感染鳥類[6]。
H9N2亞型AIV 通常表現為低致病性,很少導致家禽的直接死亡[7],H9N2禽流感病毒的宿主范圍已經非常廣泛,家禽、野鳥中都有報道,且宿主范圍還在不斷擴大,有研究從豬等低等哺乳動物體內成功分離到了該亞型病毒[8]。H9N2 亞型 AIV 廣泛存在變異重組現象,可能為形成新型跨種傳播感染人的流感病毒株提供條件。許多研究表明H9N2 AIV 可能是其他亞型AIV內部基因的供體[9-10],1997年,香港人感染H5N1經研究發現,分離到的H5N1病毒與H9N2 的內部基因相似性在98%—99%,2013年爆發的感染人H7N9亞型及2014年報道的感染人H10N8 亞型AIV其內部基因與H9N2亞型內部基因高度同源[11-12],對H9N2 亞型 AIV 進行流行病學監測,了解其生物學特性具有重要的公共衛生意義。
本研究對從送檢死亡的綠鷺分離的1株H9N2 病毒(A/striated heron/Yunnan/2018)進行生物信息學分析,以期了解該亞型病毒在野生水禽綠鷺中的進化和生物學特性,為今后科學防控禽流感提供依據。
1 材料與方法
1.1 樣品來源
云南省某地區林業部門送檢死亡的綠鷺,經實驗室檢測H9N2感染。

圖1 送檢死亡的綠鷺Fig.1 Submitted the dead striated heron

圖2 綠鷺喉氣管剖檢Fig.2 Laryngeal trachea of striated heron
1.2 主要試劑
核酸提取試劑盒(磁珠法)購自西安天隆科技有限公司,One Step RT-PCR、膠回收試劑盒、質粒提取純化試劑盒、pMD18-T 載體連接試劑盒和DH5α感受態細胞均購自寶生物工程(大連)有限公司,H9N2檢測引物及SPF雞胚由云南省動物疫病預防控制中心提供,其他試劑均為國產分析純。
1.3 病毒分離擴增
無菌采取死亡綠鷺喉氣管加入含2%雙抗的PBS勻漿,12 000 r/min 離心 10 min,取上清200 μL接種 9—10日齡 SPF雞胚,37 ℃孵化,72 h后收集雞胚尿囊液。
1.4 病毒RNA提取和RT-PCR鑒定
按照核酸提取試劑盒(磁珠法)說明書提取尿囊液病毒總RNA;采用一步法RT-PCR使用H9N2檢測引物進行擴增。PCR產物經1.0%瓊脂糖凝膠電泳檢測分析。
1.5 全基因組序列擴增
根據文獻[7]報道對8個病毒基因片段(PB2、PB1、PA、HA、NP、NA、M和NS)全長擴增,引物由生工生物工程(上海)股份有限公司合成;產物經1.0%瓊脂糖凝膠電泳檢測,陽性產物按膠回收試劑盒說明書回收,與pMD18-T 載體連接并轉入DH5α感受態細胞,挑取陽性克隆質粒送生工生物工程(上海)股份有限公司測序。
1.6 生物信息學分析
應用 Lasergene Seqman、Lasergene MegAlign、Mega 7等軟件對測序結果進行序列拼接、進行同源性及關鍵位點分析、構建進化樹;登陸NCBI進行BLAST比對分析;通過NetNGlyc 1.0 Server在線分析毒株HA、NA蛋白上的潛在糖基化位點
2 結果
2.1 病毒PCR鑒定結果
提取滅活后尿囊液病毒核酸,使用H9N2檢測引物進行RT-PCR擴增,產物經1.0%瓊脂糖凝膠電泳后,出現特異性目的條帶(圖3),大小與預期目的片段相符。
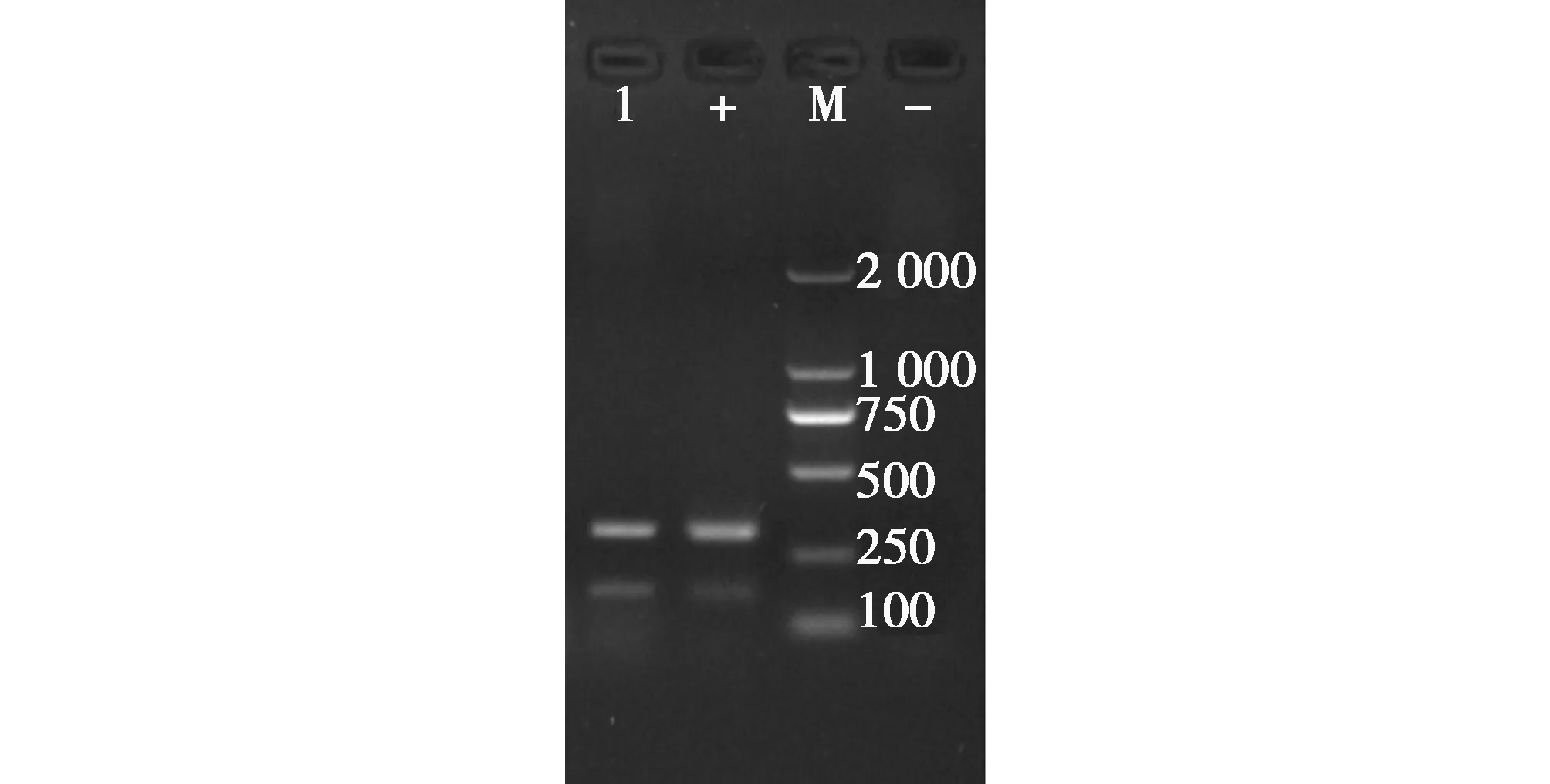
圖3 H9N2特異性引物擴增結果(M:DL2000Maker,1:樣品Sample,+:陽性對照Positive control)Fig.3 Results of amplification of H9N2-specific primers figure
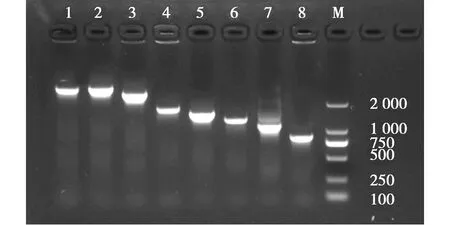
圖4 毒株全基因組序列擴增結果(M:DL2000 Maker,1:PB2,2:PB1,3:PA,4:HA,5:NP,6:NA,7:M,8:NS)Fig.4 Virus genome whole genome sequence amplification results
2.2 病毒全基因組PCR擴增結果
PCR擴增分離株各個基因片段,瓊脂糖凝膠電泳圖顯示,相應基因PCR產物與目的大小一致(圖4)。
2.3 基因組遺傳進化分析
將測序后的全基因序列經Lasergene Seqman軟件拼接校正后上傳GenBank(GenBank登錄號MN396247—MN396254),通過 Blast 比對顯示(表1),各基因片段分別與多株來自雞源、鴨源、鴿子源、野鳥源的 H9、H7、H10等多種亞型AIV高度同源,其中PB1與多株H10N8亞型有較高同源性、PA與1株人源H7N9亞型和多株雞源H7N9亞型有較高同源性;遺傳進化分析結果(圖5—12)顯示,PB2、M基因位于G1-like分支[13],NA、HA基因位于Y280-like分支,PA、PB1、NP、NS基因位于F98-like分支。提示該分離株基因來源復雜,可能為不同亞型間病毒基因發生重組所造成。
表1 分離株各基因片段 Blast分析結果

Tab.1 Blast analysis of gene fragments of virus isolates
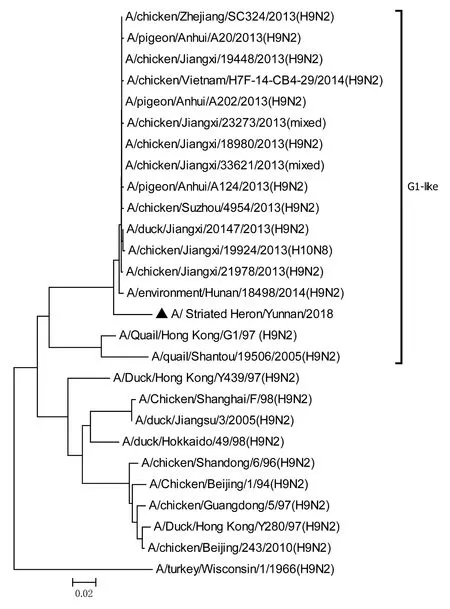
圖5 PB2基因進化樹Fig.5 Phylogenetic tree of PB2 genes
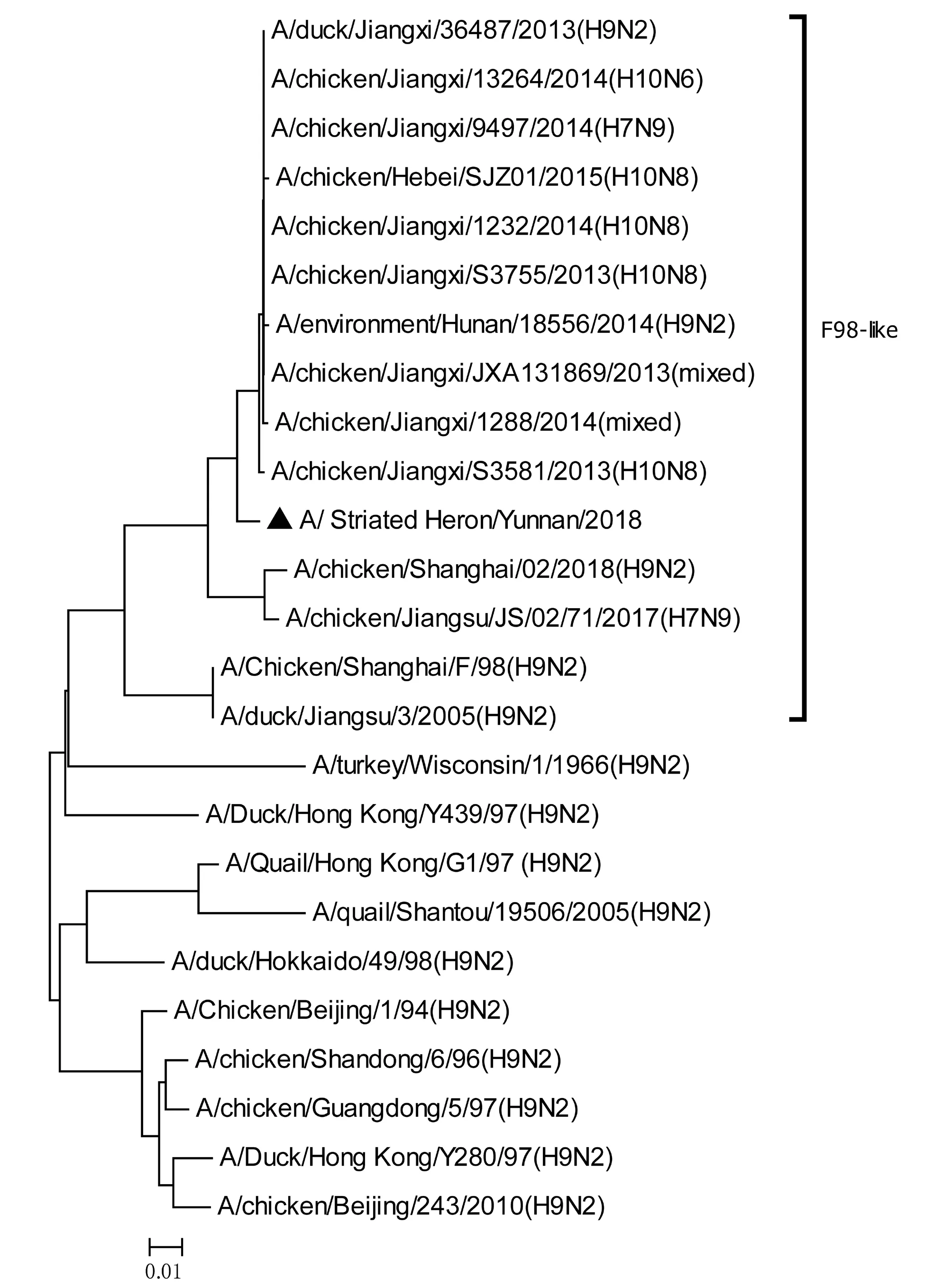
圖6 PB1基因進化樹Fig.6 Phylogenetic tree of PB1 genes
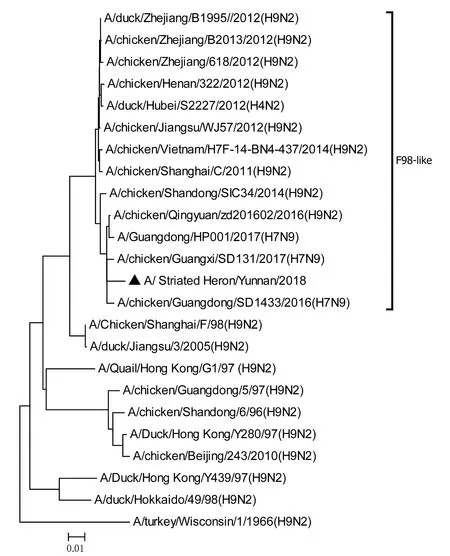
圖7 PA基因進化樹Fig.7 Phylogenetic tree of PA genes
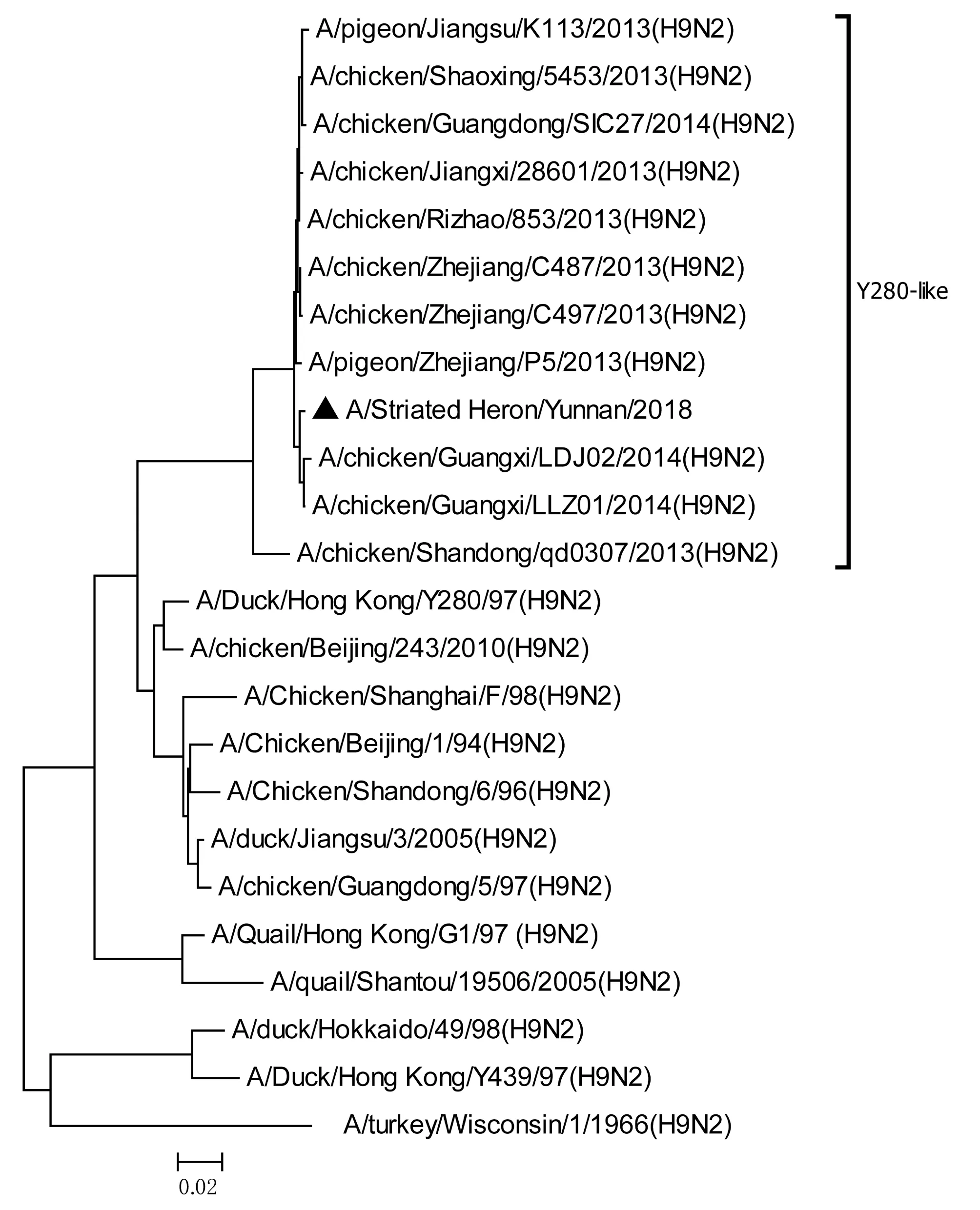
圖8 HA基因進化樹Fig.8 Phylogenetic tree of HA genes

圖9 NP基因進化樹Fig.9 Phylogenetic tree of NP genes
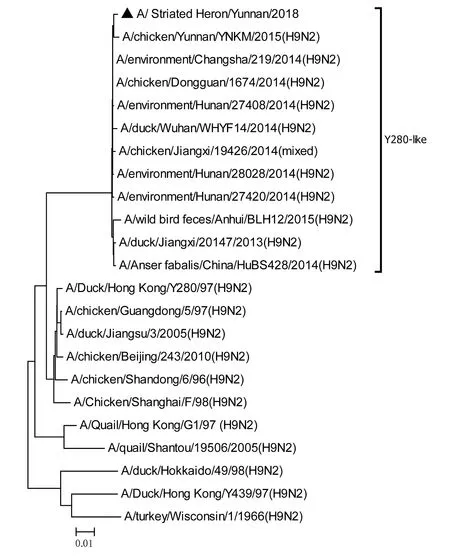
圖10 NA基因進化樹Fig.10 Phylogenetic tree of NA genes
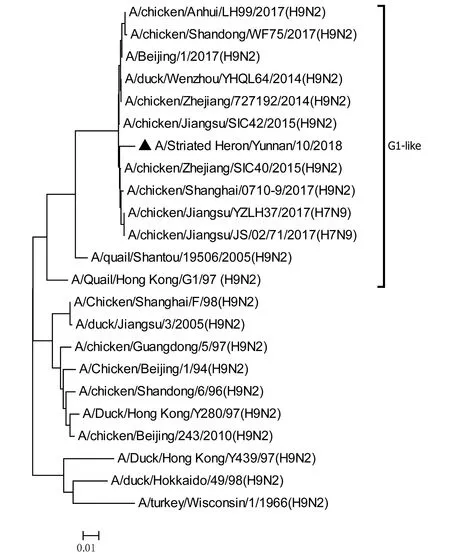
圖11 M基因進化樹Fig.11 Phylogenetic tree of M genes

圖12 NS基因進化樹Fig.12 Phylogenetic tree of NS genes
2.4 基因組序列關鍵位點分析
分離株的HA核苷酸序列推導氨基酸序列分析顯示,HA裂解位點第334位氨基酸發生A334S突變,其序列為333PSRSSR↓GL340,為非連續堿性氨基酸,符合 LPAIV 氨基酸序列特征。分離株受體結合位點109、161、163、191、198、202、203氨基酸序列為YWTNTLY,第198位氨基酸為T與 A/Duck/Hongkong/Y280/1997毒株一致。第 234位氨基酸(相當于H3 第226位氨基酸)發生Q234L突變。存在8個潛在糖基化位點(表 2)依次為29NST31、141NVS143、145NGT148、298NTT300、305NVS307、313NCS315、492NGT494、551NGS553;可見 29、141、298、305、492、551位點相對保守,與參考毒株比較,缺失218位點氨基酸,增加了145、313 位點氨基酸,第145位氨基酸發生S145N突變增加了一個糖基化位點(表3)。
表2 HA 裂解位點和受體結合位點的比較分析
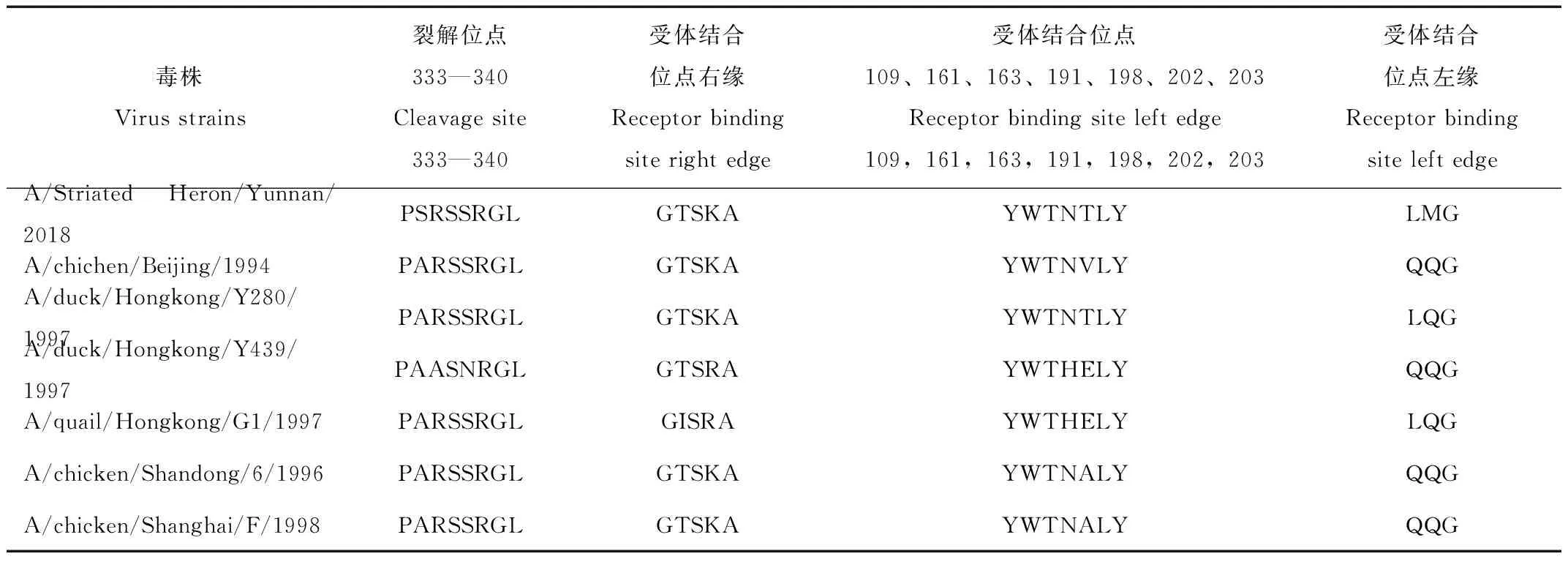
Tab.2 Comparative analysis of HA cleavage site and receptor binding site
表3 HA 潛在糖基化位點的比較分析
表4 NA潛在糖基化位點和紅細胞結合位點分析

Tab.4 Analysis of NA glycosylation sites and HB site
分離株NA推導氨基酸序列分析顯示,頸部處缺失了63TEI65 3個氨基酸,與經典株及疫苗株比較在368位增加一個位點,在402位減少一個位點,共存在6個潛在的糖基化位點69NTS71、86NWS88、146NGT148、200NAT202、234NGT236、368NGS370(表4)。NA蛋白紅細胞結合位點分析發現,367—372位氨基酸序列為 KNGSRS,399—404氨基酸序列為SDDW,431—433位點則相對保守均為PQE氨基酸基序列(表4)。
內部基因序列及推導氨基酸序列分析析顯示:該分離株 PB2未出現與哺乳動物適應相關的E627K和D701N突變;PB1-F2全長54 aa,未出現N66S突變;PA未出現T97I、K615N S224P和N383D突變,NP未出現K184A、N319K突變;M基因中M1蛋白出現N30D、T215A 突變;M2蛋白出現S31N突變;NS 80—84處頸部氨基酸未見缺失。
3 討論
綠鷺作為候鳥是AIV的自然儲存庫,病毒可隨其遷徙長距離傳播,可通過糞-口途徑或污染的水,在野生水禽之間或野生水禽與家禽之間傳播。H9N2亞型AIV 通常表現為低致病性,很少導致家禽的直接死亡[7],該死亡綠鷺經實驗室診斷為細菌及寄生蟲感染導致的死亡,但不排除重組后造成毒力增強造成其免疫力下降,從而導致的后續感染,有待進一步探究。
Sun等[14]的研究顯示1994—2008年中國主要流行CK/BJ-like分支的病毒,1998—2004年CK/BJ-like和F98-like分支的病毒共同流行但F98-like分支占主導,2004—2008年F98-like分支成為優勢毒株。2006年后Y280-like分支逐漸成為主要流行毒株[15]。但近年來 H9N2 亞型 AIV 不斷發生重組,國內流行的H9N2 株出現越來越多的重組基因型,本研究對野生水禽綠鷺中分離到的1株 H9N2 亞型 AIV分離株HA、NA基因位于Y280-like分支,PB2、M基因位于G1-like分支,PA、PB1、NP、NS基因位于F98-like分支。遺傳演化分析表明分離株可能來自家禽雞、鴨或其他野鳥,該分離株可能在不同宿主間交互傳播。內部基因與H7、H9、H10等多種亞型的 AIV 同源性較高,其中PB1基因與多株H10N8亞型有較高同源性、PA基因與一株人源H7N9亞型和多株雞源H7N9亞型有較高同源性,表明了內部基因來源復雜,可能為不同亞型間病毒基因發生重組所造成。
HA受體結合位點發生Q234L突變,提示該分離株具有和人流感受體唾液酸-α2,6-半乳糖受體結合的特性[16],存在人獸共患風險,裂解位點為333PSRSSR↓GL 340,符合低致病性禽流感毒株特征,S145N突變增加了一個糖基化位點,提示該位點的出現可能會使毒株致病性提高,免疫原性發生改變[17],此外增加的313NCS315糖基化位點靠近HA裂解位點,新增糖基化位點有可能影響蛋白酶對裂解位點的識別作用,從而對病毒毒力造成影響[18]。
NA蛋白頸部處缺失了63TEI65 3個氨基酸,該缺失可能導致 NA 酶活性的增加,在結合位點發生突變可能與毒力變化有關;增加了一個潛在糖基化位點368NGS370。
內部基因PB2氨基酸序列相對保守,毒力相關位點未見改變;PB1-F2全長54 aa未出現N66S突變,研究表明,PB1-F2全長大于 78 aa 能夠增強病毒的致病力,同時N66S單個氨基酸殘基替換使病毒致病力增加[19];PA蛋白上的T97I突變可以提高流感病毒對哺乳動物的致病性[20],K615N突變可以提高流感病毒對小鼠的致病性,S224P突變和N383D突變能提高流感病毒對鴨的致病性[21];NP蛋白的K184A突變與病毒毒力和增殖能力相關[22],N319K會增加病毒聚合酶的活性來增強病毒對小鼠的致病性[23];本次分離株PA、NP均未出現突變,仍是低致病性病毒的分子特征。M1蛋白出現N30D、T215A 突變可能使毒株對小鼠致病性產生影響[15]。M2 蛋白S31N突變,可導致流感病毒對金剛烷胺類藥物產生耐藥性[24]。NS蛋白 80—84處頸部氨基酸未見缺失,有研究表明80—84位氨基酸殘基的缺失,能增強病毒毒力或提高對細胞因子的抗性,是對哺乳動物高致病性的一個明顯特征[25]。
該分離株與毒力、宿主特異性、耐藥性相關的位點存在不同程度的突變,內部基因與H7、H10亞型流感病毒間存在基因重組風險對公共衛生安全產生潛在的威脅。野生水禽作為重要的自然宿主在疾病傳播過程起著關鍵作用,本研究中從野生水禽綠鷺中分離到的H9N2亞型基因重組復雜,具有感染人類的潛在風險。加強對野生禽類禽流感病毒生物學監測具有重要意義。
