低溫和限食對黑線倉鼠貯食行為的影響
2020-05-06 08:48:56李思懿劉振山何巧巧
野生動物學報 2020年2期
關鍵詞:差異
李思懿 劉振山 李 婧 何巧巧 楊 明
(沈陽師范大學生命科學學院,沈陽,110034)
自然界中許多動物在應對食物資源的波動和不可預期的變化中,進化出了貯食行為(food hoarding behavior),即在食物充足的時期將食物貯存起來,以備食物缺乏時利用。貯食行為是許多小型哺乳動物應對食物資源短缺以滿足自身能量需求的重要策略之一[1]。具有貯食行為的動物能在時間和空間上控制食物的可利用性[1-2],應對季節性的食物波動[3]。通過貯存食物可以減少食物短缺帶來的不良影響,同時依靠積累食物可以克服能量短缺的脅迫[4]。動物貯存食物行為的出現,使貯食動物在食物缺乏期的生存概率得到明顯的提高;貯藏的食物在食物短缺期,或為交配、繁殖后代等提供了能量[5],進而提高動物的繁殖成功率。因此,貯食行為的出現也提高了個體的適合度[6]。沒有貯食行為的動物在應對食物短缺時,通常需要遷移、動用身體能量儲備(如體脂),或者進入代謝抑制狀態,如冬眠、蟄眠等[1]。
貯食行為作為一種特殊的覓食策略,在嚙齒類動物和鳥類中普遍存在[1]。貯食行為會受到季節性環境信號、性別和外部食物資源等多種因素的影響。貯食行為會受到環境季節信號的影響,溫度、光周期的變化常被動物用作季節性環境信號來預測未來環境條件的變化,并做好應對的準備。如秋季的低溫和短光照可以誘導小型哺乳動物產熱器官褐色脂肪組織增生,產熱能力增加[7],為增加冬季產熱能力維持恒定體溫做好準備。雜色山雀(Sittiparusvarius)冬季僅啄取和貯存松子,春季取食和貯存少量葵花籽[8]。長爪沙鼠(Merionesunguiculatus)的貯食行為表現出明顯的季節性,主要在秋季貯存食物[9]。性別差異對嚙齒動物的食物貯藏行為存在一定影響[10],大多數嚙齒類動物中,雌性的貯食量大于雄性。已有研究報道,雌激素能夠顯著影響實驗大鼠的食物貯藏行為,睪酮也能顯著抑制雄性長爪沙鼠對食物的貯藏行為[11]。另如對金色中倉鼠(Mesocricetusauratus)[12]、長爪沙鼠[13]、刺囊鼠(Liomyssalvini)[14]的研究發現,雌性個體貯存食物的行為比雄性個體更為強烈。外部食物資源也是影響動物貯食行為的一個重要因素。在食物資源缺乏時,動物攝食不足,會面臨饑餓風險,一般認為,與動物的食物貯藏行為密切相關的攝食行為的調節分為短期和長期兩種水平[1]。而攝食和貯藏之間可能至少會形成3種關系:第一種關系是當動物在饑餓的情況下尋覓到食物時,選擇先吃掉一部分食物,來解決目前的饑餓問題,因此貯藏活動被推遲;第二種關系是饑餓對食物貯藏活動有一定的刺激或促進作用;第三種關系是饑餓感受對貯食行為沒有影響[6]。
自然條件下的食物資源短缺狀態可以在實驗室利用剝奪食物(禁食)或者減少食物供應(限食)的方式來模擬[15]。一些嚙齒動物在經歷禁食后重新獲得食物時,會表現出短暫的攝食量急劇增加的現象,如褐家鼠(Rattusnorvegicus)[16-17];也有些動物則表現出攝食量不增加但貯食量增加[4],如黑線毛足鼠(Phodopussungorus)[18-19]等。增加貯食量可能比增加攝食量具有更多優勢,如動物能夠留在比較安全的洞穴環境中,通過食用貯存的食物來維持其每日能量攝入[15]。
黑線倉鼠(Cricetulusbarabensis)屬于嚙齒目(Rodentia),倉鼠科(Cricetidae),倉鼠屬的一種小型哺乳動物,是我國黃河以北一些省份的優勢鼠種。其體型較小,多以玉米、豆類等的種子為食,偶爾也取食少量的昆蟲和植物的根、莖、葉等,屬于一種雜食性鼠類。該物種分布地冬季嚴寒,食物資源短缺,一般會通過蟄眠和貯食以越過寒冷冬季。本實驗室前期的研究發現,該物種有貯食行為,且該物種中體重小的雌性個體會在短光低溫的條件下發生日蟄眠[20],其余的個體沒有發生日蟄眠。但是其貯食的影響因素及其與蟄眠的關系尚不清楚。本研究的目的是探究低溫和限食與黑線倉鼠貯食行為之間的關系,分析該物種應對冬季低溫和食物短缺的能量策略。
1 研究方法
1.1 實驗動物
實驗所用黑線倉鼠選自遼寧省鐵嶺市的實驗室內繁殖種群。在沈陽師范大學動物房內單籠(29 cm×18 cm×16 cm)飼養,籠內用碎刨花和少量棉花作為巢材。動物房內環境溫度為(20±2)℃,光照條件設定為16L∶8D,標準鼠飼料采購于沈陽市于洪區前民實驗動物飼料廠,自由取食、飲水。
1.2 貯食行為的測定裝置和方法
動物貯食行為的測定裝置由1根半透明塑料管連接2個飼養籠組成,半透明塑料管內徑為5 cm、長約90 cm,2個籠子分別作為“家巢籠”(home cage)和“食物籠”(food cage)[4],籠的大小均為29 cm×18 cm×16 cm的清潔塑料籠(圖1)。其中,家巢籠內放入碎刨花和少量棉花作為巢材,類似于實驗前的飼養籠;食物籠內無巢材和棉花,籠內放置稱量好的食物[4]。在實驗測定前,動物在貯食行為測定裝置中適應2周,動物在適應期間不進行任何測量[4,21]。
測定動物的貯食行為時,將搬離食物籠的料塊全部記為貯存的食物,沒有被搬出食物籠的食物記為剩余食物。用24 h貯存食物的重量衡量貯食行為的強度。24 h后分別將剩余的食物以及貯存的食物取出稱重,放入的總食物量減去剩余食物量以及貯存食物量為動物1 d的攝食量。
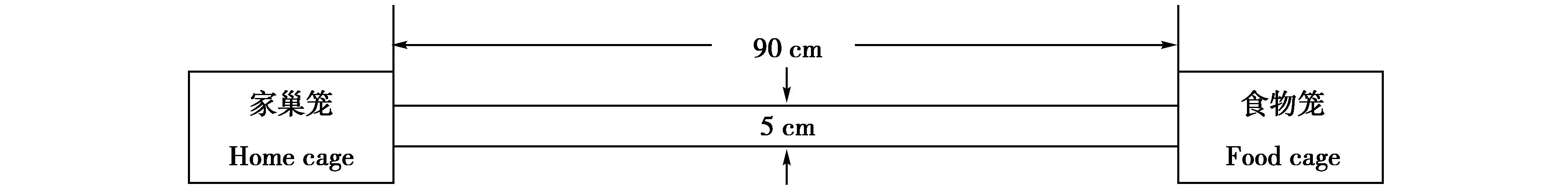
圖1 動物貯食行為的測定裝置示意圖Fig.1 Scheme of food hoarding apparatus
1.3 溫度和光周期對兩性黑線倉鼠貯食行為的影響
隨機選取在實驗室內繁殖的黑線倉鼠20只,雌雄各10只。分成4組,每組5只動物,長光暖溫雄性組(LW-♂)、長光暖溫雌性組(LW-♀),這兩組置于環境溫度為(20±2)℃,光照∶黑暗=16∶8的環境中;等光低溫雄性組(EC-♂)、等光低溫雌性組(EC-♀),這兩組置于環境溫度為(10±2)℃,光照∶黑暗=12∶12的環境中。喂食經過稱量的超量的鼠飼料,自由取食、飲水,每日14:00測定動物體重、攝食量和貯食量,連續測定1周。體重、食物重量測定使用精確到0.01 g的電子天平(UTP-313,上海花潮電器有限公司)。
1.4 限食對黑線倉鼠貯食行為的影響
隨機選取實驗室內繁殖的動物8只,均為雌性。實驗在溫度為(20±2)℃,光照∶黑暗=16∶8的環境中進行。喂食經過稱量的超量的鼠飼料及兔飼料,自由取食、飲水,每日測定1次動物體重、攝食量和貯食量,連續測定1周,此為非限食階段;2周之后開始限食階段,每日9:00—17:00去除食物進行限食處理,每日17:00至次日9:00給食經過稱量的超量的鼠飼料及兔飼料,水自取,每日17:00測定1次動物體重、攝食量和貯食量,再連續測定1周。體重、食物重量測定使用精確到0.01 g的電子天平(UTP-313,上海花潮電器有限公司)。
1.5 食物的主要成分
實驗中使用的食物分別為實驗用標準鼠飼料和實驗用標準兔飼料,食物的主要成分為水分、粗蛋白、粗脂肪、粗纖維、粗灰分、鈣、磷、賴氨酸、蛋氨酸+胱氨酸,鼠飼料和兔飼料的營養配比如表 1所示。
1.6 統計分析
采用SPSS 20.0軟件進行數據分析。在統計分析前,用Kolmogorov-Smirnov test檢驗數據的正態性,用Levene test檢驗齊次性。不同溫度光周期與性別對黑線倉鼠體重、攝食量、貯食量的影響采用雙因素方差分析的方法;相同溫度光周期兩性黑線倉鼠及不同溫度光周期同性黑線倉鼠的初始體重、結束體重、體重變化量、體重變化率、攝食量、貯食量比較采用獨立樣本t檢驗的分析方法;動物實驗前后體重差異、非限食階段與限食階段黑線倉鼠初始體重、結束體重、體重變化量、體重變化率、攝食量、貯食量的比較均采用配對樣本t檢驗的分析方法。文中結果均以平均值±標準誤呈現。P≤0.05為差異顯著,P≤0.01為差異極顯著。
表1 鼠飼料和兔飼料主要成分的含量對比
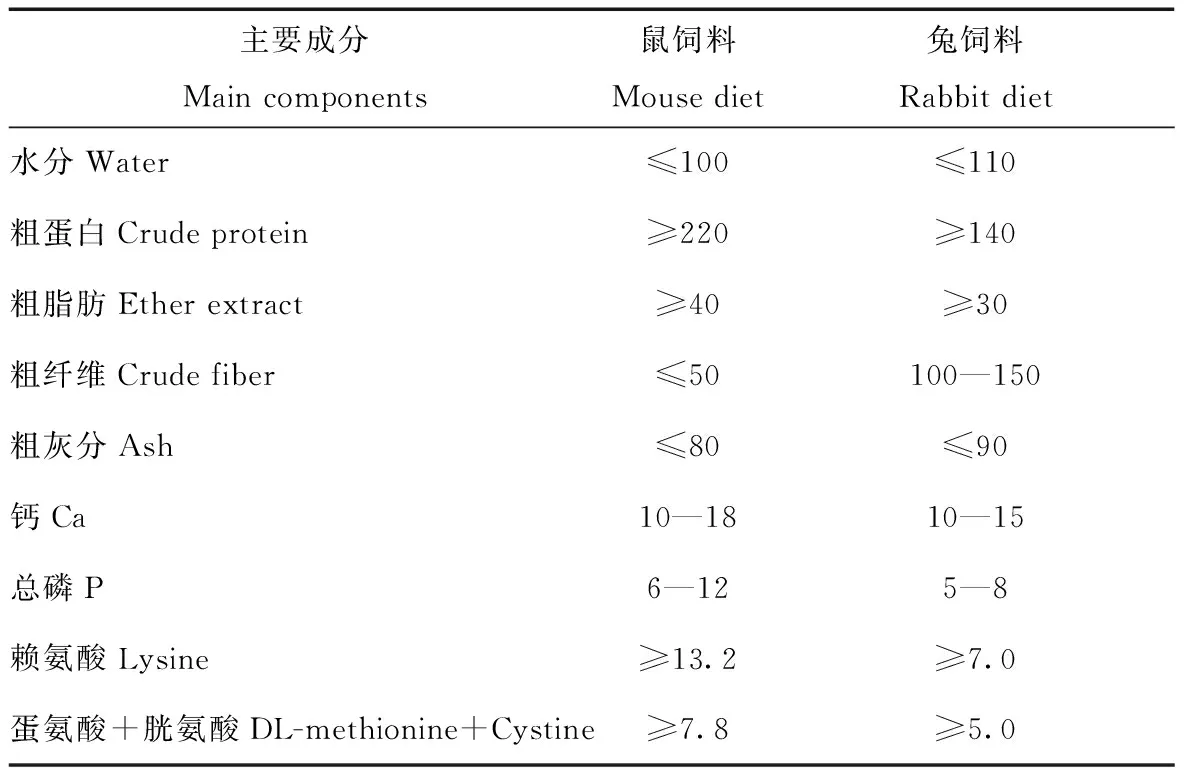
Tab.1 Comparison of main components between mouse diet and rabbit diet g/kg
2 結果
2.1 溫度光周期對兩性黑線倉鼠的影響
2.1.1 溫度光周期對兩性黑線倉鼠體重的影響
雙因素方差分析結果顯示:溫度光周期對體重變化量的影響極顯著(F=11.488,P=0.004);性別對體重變化量的影響不顯著(F=0.286,P=0.600);溫度光周期與性別對體重變化量的影響無顯著的交互作用(性別*溫度光周期F=0.370,P=0.551)。
4組黑線倉鼠的實驗初始體重分別為:LW-♂為(37.27±2.50)g(n=5),LW-♀為(26.69±1.04)g(n=5),EC-♂為(33.55±1.94)g(n=5),EC-♀為(27.03±0.75)g(n=5)。LW兩性別之間存在極顯著差異(F=0.893,P=0.003);SC兩性別之間存在極顯著差異(F=1.481,P=0.008);雄性兩組之間無顯著差異(F=0.094,P=0.391);雌性兩組之間無顯著差異(F=0.582,P=0.590)。實驗結束時4組動物體重分別為:LW-♂為(37.27±2.50)g(n=5),LW-♀為(26.69±1.04)g(n=5),EC-♂為(33.55±1.94)g(n=5),EC-♀為(27.03±0.75)g(n=5)。LW兩性別之間存在極顯著差異(F=0.292,P=0.005);SC兩性別之間存在顯著差異(F=0.245,P=0.014);雄性兩組之間無顯著差異(F=0.582,P=0.590);雌性兩組之間無顯著差異(F=0.267,P=0.579)。實驗前后動物體重:LW-♂存在極顯著差異(t=8.045,P=0.001);LW-♀無顯著差異(t=2.445,P=0.071);EC-♂存在極顯著差異(t=6.445,P=0.003);EC-♀無顯著差異(t=1.550,P=0.196)。實驗前后體重變化量:LW兩性別之間存在顯著差異(F=3.052,P=0.612);SC兩性別之間存在顯著差異(F=2.480,P=0.583);雄性兩組之間無顯著差異(F=1.217,P=0.209);雌性兩組之間無顯著差異(F=0.102,P=0.696)。體重變化率:LW兩性別之間存在顯著差異(F=2.987,P=0.305);SC兩性別之間存在顯著差異(F=3.855,P=0.932);雄性兩組之間無顯著差異(F=0.563,P=0.113);雌性兩組之間無顯著差異(F=0.098,P=0.655)(表2)。經1周的低溫等光處理雌性動物體重未改變,而雄性動物體重顯著降低。
2.1.2 溫度光周期對兩性黑線倉鼠的攝食量的影響
雙因素方差分析結果顯示:溫度光周期對攝食量的影響極顯著(F=8.518,P=0.010);性別對攝食量的影響極顯著(F=18.433,P=0.001);性別與溫度光周期對攝食量的影響無顯著的交互作用(性別*溫度光周期F=2.036,P=0.173)。
4組黑線倉鼠實驗期間的攝食量分別為:LW-♂為(5.43±0.14)g/d(n=5),LW-♀為(3.97±0.28)g/d(n=5),EC-♂為(5.76±0.15)g/d(n=5),EC-♀為(4.92±0.14)g/d(n=5)。LW兩性別之間存在極顯著差異(F=3.143,P=0.002);SC兩性別之間存在極顯著差異(F=0.118,P=0.004);雄性兩組之間無顯著差異(F=0.162,P=0.153);雌性兩組之間存在顯著差異(F=3.214,P=0.017)(表2)。經1周的低溫等光處理,雄性動物的攝食量未改變,而雌性動物的攝食量顯著增加。
2.1.2 溫度光周期對兩性黑線倉鼠貯食量的影響
實驗中所有動物均出現貯食行為,平均每只動物每天貯存食物量為(15.37±11.77)g,說明黑線倉鼠有貯食行為。
表2 4組黑線倉鼠體重及攝食量結果比較
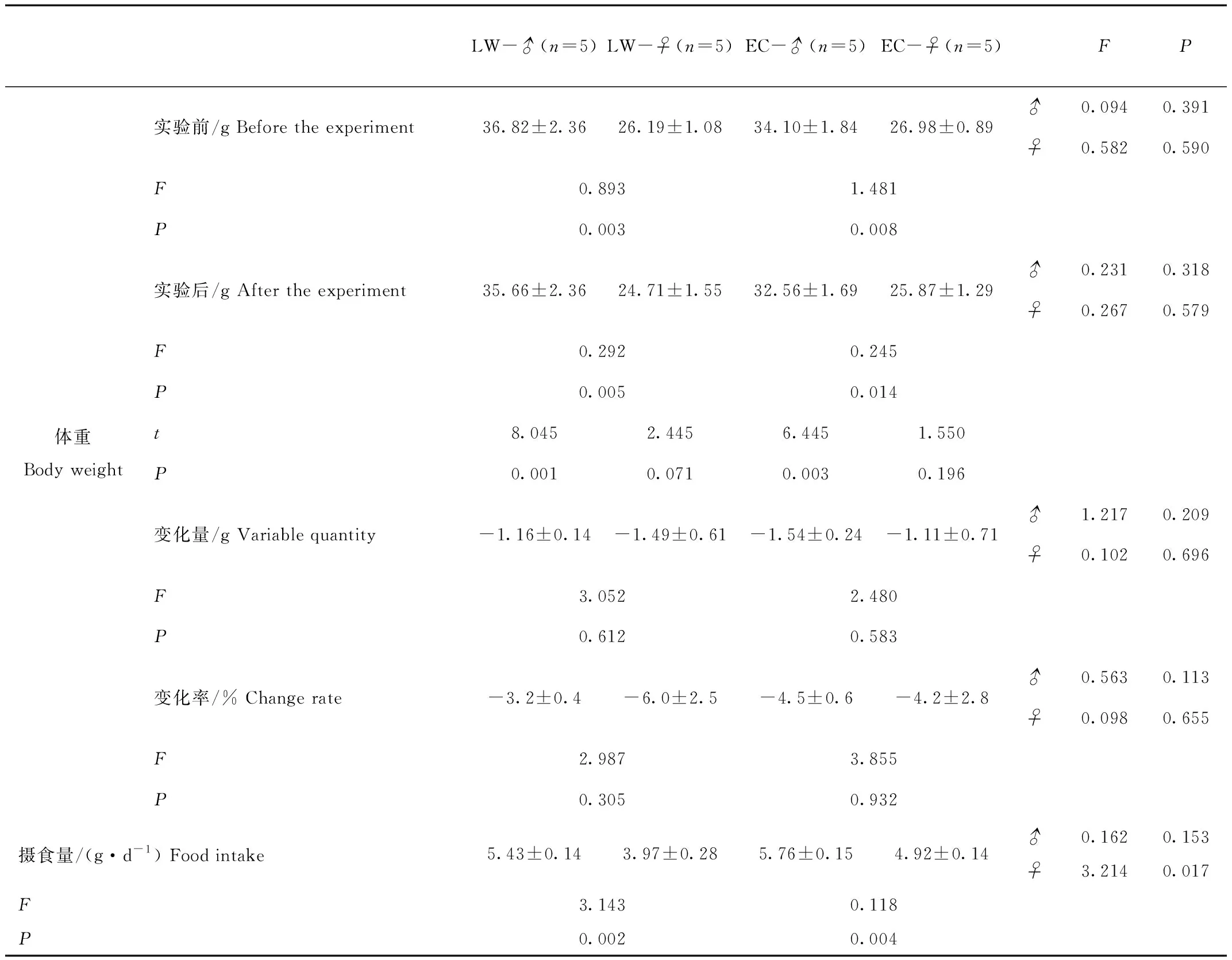
Tab.2 Different of body weights and food intake between four groups striped hamster
雙因素方差分析結果顯示:光照溫度對貯食量無顯著影響(F=2.107,P=0.167);性別對貯食量的影響極顯著(F=11.623,P=0.004);且性別與光照溫度對貯食量的影響無顯著的交互作用(性別*光照溫度F=0.315,P=0.583)。4組黑線倉鼠實驗期間貯食量分別為:EC-♂為(23.17±3.67)g/d(n=5),EC-♀為(10.9±2.89)g/d(n=5),兩性別之間差異顯著(F=0.590,P=0.030);LW-♂為(18.49±2.52)g/d(n=5),LW-♀為(9.23±2.71)g/d(n=5),兩性別之間差異顯著(F=0.066,P=0.027);LW-♂與EC-♂之間無顯著差異(F=0.643,P=0.303);LW-♀與EC-♀之間無顯著差異(F=0.102,P=0.489)(圖2)。
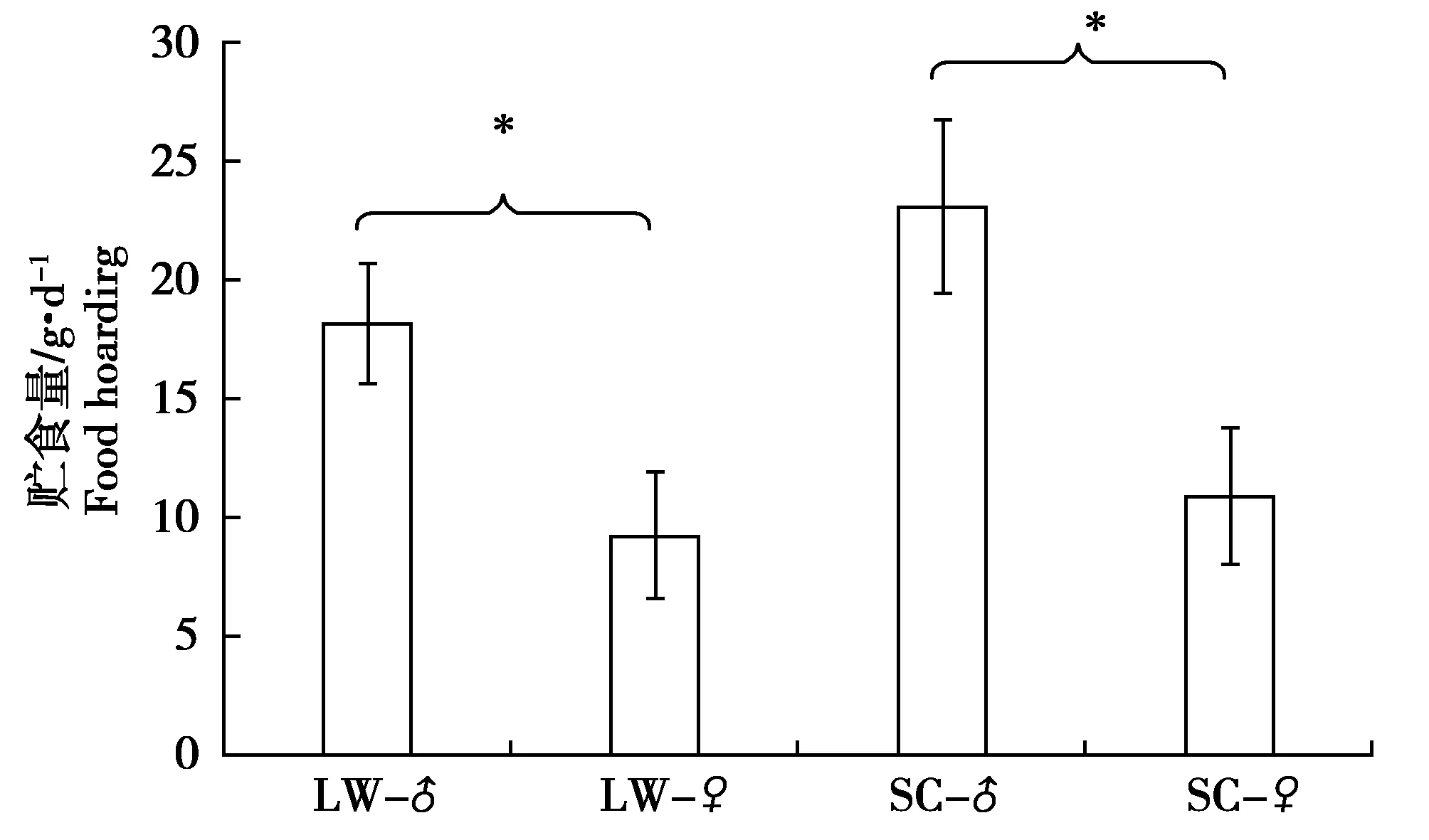
圖2 4組黑線倉鼠貯食量比較Fig.2 Different of hoard size for four groups striped hamster 注:同一環境條件黑線倉鼠雌雄間差異顯著性差異用*表示,P<0.050 Note:*Indicate the significant difference between male and female striped hamsters in the same environment,P<0.050
2.2 限食對黑線倉鼠的影響
2.2.1 限食對黑線倉鼠體重的影響
非限食階段黑線倉鼠初始體重為(24.75±0.77)g(n=8),結束體重為(24.23±0.81)g(n=8),體重變化量為(-0.53±0.18)g,體重變化率為(-2.2±0.8)%;限食階段初始體重為(24.25±0.65)g(n=8),結束體重為(21.94±0.56)g(n=8),體重變化量為(-2.31±0.33)g,體重變化率為(-9.5±1.3)%。其中非限食階段初始體重與結束體重之間存在顯著差異(t=2.885,P=0.023);限食階段初始體重與結束體重之間存在極顯著差異(t=6.460,P=0.000);兩階段初始體重沒有顯著差異(t=0.192,P=0.853);結束體重存在極顯著差異(t=4.368,P=0.003);兩階段體重變化量存在極顯著差異(t=4.832,P=0.002);兩階段體重變化率存在極顯著差異(t=5.340,P=0.001)。
2.2.2 限食對黑線倉鼠攝食量的影響
黑線倉鼠非限食階段攝入鼠飼料(10.31±1.94)g/d(n=8),兔飼料(20.14±0.99)g/d(n=8),合計(30.45±1.73)g/d;限食階段攝入鼠飼料(10.38±1.63)g/d(n=8),兔飼料(17.95±1.24)g/d(n=8),合計(28.33±1.82)g/d。其中非限食階段鼠飼料與兔飼料的攝食量存在極顯著差異(t=-3.860,P=0.006);限食組鼠飼料與兔飼料的攝食量存在顯著差異(t=-3.349,P=0.012);非限食階段與限食階段之間鼠飼料攝食量無顯著差異(t=-1.119,P=0.300);兔飼料攝食量無顯著差異(t=-0.336,P=0.747);攝入食物總量無顯著差異(t=-0.944,P=0.376)(表3)。
2.2.3 限食對黑線倉鼠貯食量的影響
黑線倉鼠非限食階段貯存鼠飼料(10.25±2.88)g/d(n=8),兔飼料(57.80±39.96)g/d(n=8),合計(68.05±40.95)g/d(n=8);限食組階段貯存鼠飼料(47.91±22.58)g/d(n=8),兔飼料(41.75±19.53)g/d(n=8),合計(89.66±38.95)g/d(n=8)。其中非限食階段鼠飼料與兔飼料的貯食量無顯著差異(t=-1.213,P=0.264);限食階段鼠飼料與兔飼料的貯食量無顯著差異(t=0.378,P=0.717);非限食階段與限食階段之間鼠飼料貯食量無顯著差異(t=-1.801,P=0.115);兔飼料貯食量無顯著差異(t=0.301,P=0.772);貯存食物總量無顯著差異(t=-0.657,P=0.532)(表3)。
表3 非限食階段與限食階段體重、攝食量、貯食量結果比較

Tab.3 Different of body weights,food intake and hoard sizes in food non-restricted and restricted stage
3 討論
3.1 體重和攝食量
實驗過程中所有實驗動物體重均存有下降趨勢,這可能與每日對其稱重有關,但對于每只動物稱重的裝置及手法相同,因此可以認為本研究中人為干擾對每只動物的影響是恒定的,不會隨著組別或個體而改變。黑線倉鼠具有雄性個體大于雌性個體的特性,低溫等光誘導后,雄性黑線倉鼠體重顯著降低,而攝食量并沒有顯著增加;雌性個體體重保持穩定,攝食量顯著增加,但是依然顯著低于同一環境中的雄性動物。說明在面對寒冷時,雌雄個體表現出不同的應對策略,即,雄性個體優先選擇消耗自身能量來維持正常生命活動,雌性黑線倉鼠會通過增大攝食量,來保持恒定的體重對抗惡劣環境。在白天時段進行限食處理后,雌性黑線倉鼠體重也顯著降低,而攝食量沒有變化。前期研究發現,黑線倉鼠是一種夜行性動物[20],這可以解釋為何在白天時段對黑線倉鼠進行限食,其食物攝入量未能改變。通過對限食階段與非限食階段組內鼠飼料和兔飼料的攝食量進行比較,發現兩組中兔飼料的攝食量均顯著高于鼠飼料,說明黑線倉鼠更愿意食用兔飼料。
3.2 貯食量
實驗中所有動物均具有貯食現象,說明黑線倉鼠具有貯食行為,但其貯食量存在個體差異。縮短光照降低溫度沒有影響黑線倉鼠的貯食,這一點與長爪沙鼠的研究不同[9];雄性黑線倉鼠的平均貯食量明顯高于雌性黑線倉鼠,與金色中倉鼠、長爪沙鼠、刺囊鼠的研究結果相反[12-13]。前期研究發現黑線倉鼠有50%的雌性個體會發生蟄眠的行為,而雄性均不發生蟄眠[20],這可能與雄性動物體型較大且貯存食物量多于雌性有關。在溫度降低光照縮短(類似于秋季)的環境中,雌性黑線倉鼠會增大攝食量維持恒定體溫,但并沒有貯存更多食物,由于雌性個體自身體型較小,消化器官的最大消化吸收能力也相對較低,若進一步降低溫度縮短光照,外界環境更加惡劣,雌性黑線倉鼠的攝食量將達到最大閾值,即使貯存足夠多的食物,也無法全部攝入以維持恒定體溫所需的能量,因此會選擇蟄眠這一策略來抵抗惡劣環境;而雄性黑線倉鼠,相對體型較大,攝食量及其閾值也相對較大,因此可以選擇貯存更多食物,當面對寒冷環境時,一方面可以利用貯存的大量食物,另一方面也可以通過消耗自身能量來維持自身恒定溫度,滿足自身能量需求,因此不需要采取蟄眠的生存策略。此外,在長光暖溫的白天時段對雌性黑線倉鼠進行限食,未改變其貯食量,可能也與黑線倉鼠是一種夜行性動物,且長光暖溫的環境不需要動物過多消耗能量有關。
綜合上述,黑線倉鼠是一種具有貯食行為的小型哺乳動物,該物種雌性個體體型相對較小,在類似于晚秋的低溫等光照條件下,其攝食量增加,但貯食量無顯著變化;而在類似于夏季的暖溫長光條件下,進行階段性限食會導致身體能量消耗,即體重降低,但貯食量也無顯著變化。這可能是受其消化器官處理食物的限制,面臨冬季的極度寒冷時,即使有足夠的食物,也無法滿足維持恒定體溫的需求,則放松對高體溫的維持而進行日蟄眠來減少能量的消耗;雄性個體具有相對較大的體型,且貯食量明顯高于雌性,在冬季低溫能量脅迫時可以通過適量增加攝食維持恒定體溫。作為雌雄體型大小不同的物種,不同性別的黑線倉鼠應對低溫脅迫及食物缺乏表現出了不同的能量策略。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
