貉源大腸桿菌四環素耐藥基因的檢測
2020-04-30 04:32:16劉竟雅侯曉昱吳金燕
野生動物學報 2020年2期
劉竟雅 侯曉昱 吳金燕 姜 楠 馮 濤 薛 原
(東北林業大學野生動物與自然保護地學院,哈爾濱,150040)
貉(Nyctereutesprocyonoides)是一種經濟動物,目前它的經濟價值主要體現在毛皮的用途上,除了能夠制成輕便、耐寒、美觀的衣物以外。還能制成書寫刷和化妝刷一類的產品。此外,根據近期研究發現貉的膽汁有望代替熊膽入藥[1]。
在貉的各種疾病中,細菌病占了很大比例,大腸桿菌(Escherichiacoli)病常對腸道和神經系統造成嚴重損害,四環素被廣泛用作治療和預防貉細菌病的廣譜抗生素,它作為一種選擇機制[2],讓具有四環素抗性基因的大腸桿菌存活下來。這逐漸增加了大腸桿菌抗性基因頻率,并改變了種群動態。同時擁有抗性基因的大腸桿菌不斷排入到生態系統,抗性基因在種群中不斷轉移,使得該病治療的難度很大。不同的四環素類藥物的使用,令不同地區、不同農場的大腸桿菌耐藥性存在一些差異[3]
四環素藥物作為一種廣譜抗菌藥,效果好,毒性低,廉價易得,因此被廣泛用于人畜疾病預防和治療中。然而,其長期的過度使用也導致其抗性菌株的出現和增長。到目前為止,世界上許多發達國家,仍然把含有大量廣譜和高效的四環素抗生素的生長促進劑用于畜牧業,同時四環素類藥物在治療人和動物的細菌感染等方面已經使用了很長時間,導致近幾年來耐藥菌株不斷出現在我們的生態環境中[4]。
在這項研究中,我們從不同地區不同飼養場健康貉體內分離大腸桿菌,并測試其四環素耐藥基因,為將來四環素的臨床使用提供理論依據,據報道,tetA、tetB、tetC和tetM是大腸桿菌對四環素的主要耐藥基因[5]。在這個結論的指導下,我們對它進行了驗證和探索,得出兩者之間的關系,對于今后科學、有效使用四環素具有很大的意義,還可以減少貉養殖場的經濟損失,并指導合理使用藥物。
1 材料
1.1 菌株
試驗用大腸桿菌56株,分離自黑龍江省和吉林省貉養殖場。
1.2 主要試劑
10×Buffer、Dntp、Taq酶、DL-2000 Marker、6×Loading Buffer,均由TaKaRa公司提供。
2 實驗方法
2.1 PCR擴增模板的制備
將56株貉源大腸桿菌接種于LB瓊脂培養基過夜。挑取單個菌落置于LB肉湯中,37 ℃搖床培育3 h獲得一定濃度的菌液。
2.2 四環素基因的檢測
根據文獻[6-7],由吉林省庫美生物科技有限公司合成TetA、TetB、TetC、TetD、TetE、TetM這6種四環素基因的引物,其序列見表1。PCR產物在1%瓊脂凝膠中電泳。電泳結束后用凝膠成像儀成像分析。
表1 引物序列信息

Tab.1 Primer sequences information
3 結果
電泳出樣獲得了與預期長度一致的目標條帶,結果顯示本批貉大腸桿菌樣品中,四環素耐藥基因tetA、tetB、tetC、tetM均有檢出,其中以tetM檢出率最高為82.14%,tetA、tetB、tetC的檢出率依次為16.07%、28.57%、5.35%,tetE檢出率為零。
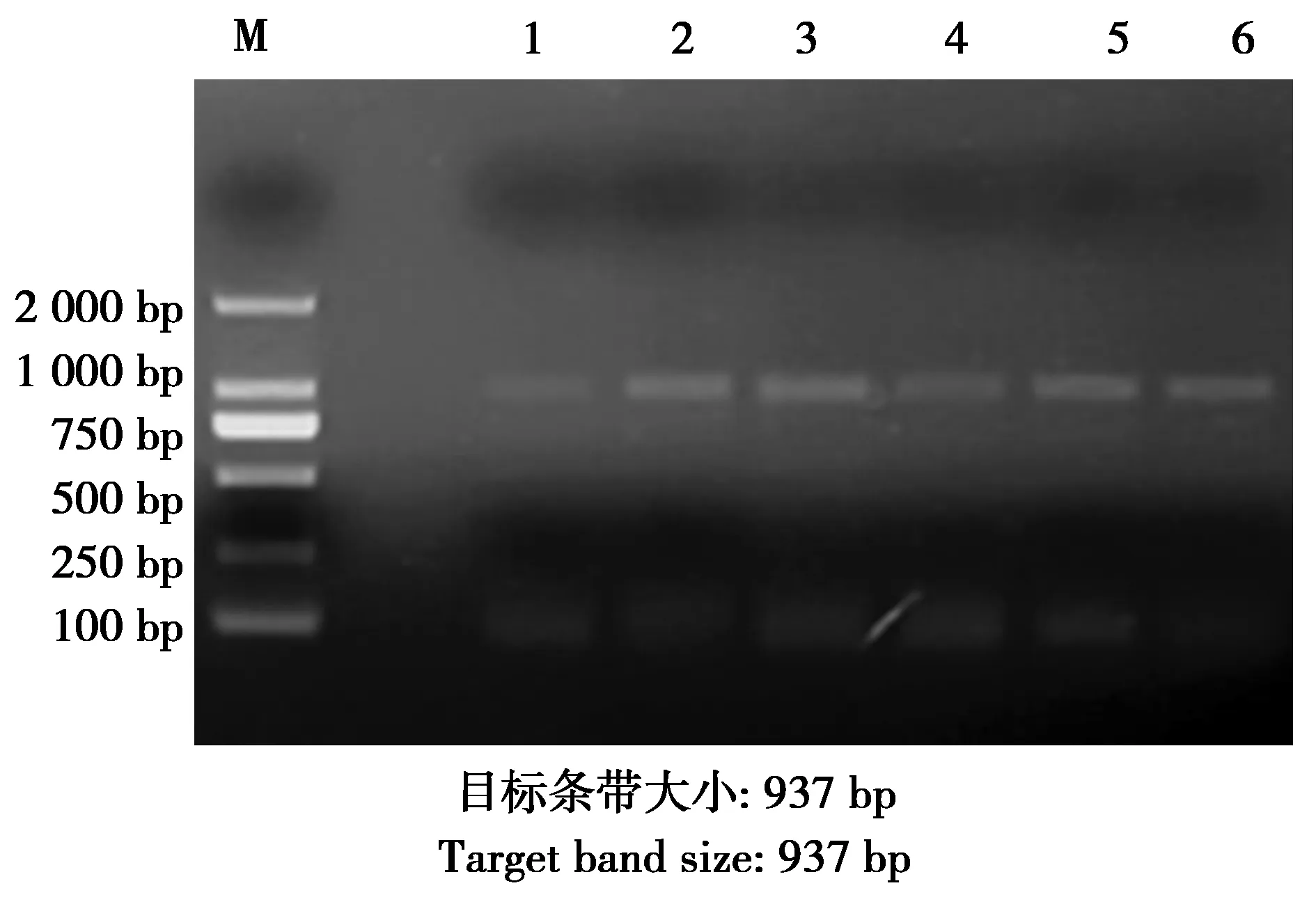
圖1 tetA的PCR擴增結果Fig.1 PCR amplified results of tetA gene 注:M:DNA marker;2—6:tetA耐藥基因的PCR擴增結果 Note:M:DNA marker.2-6:PCR products of tetA gene
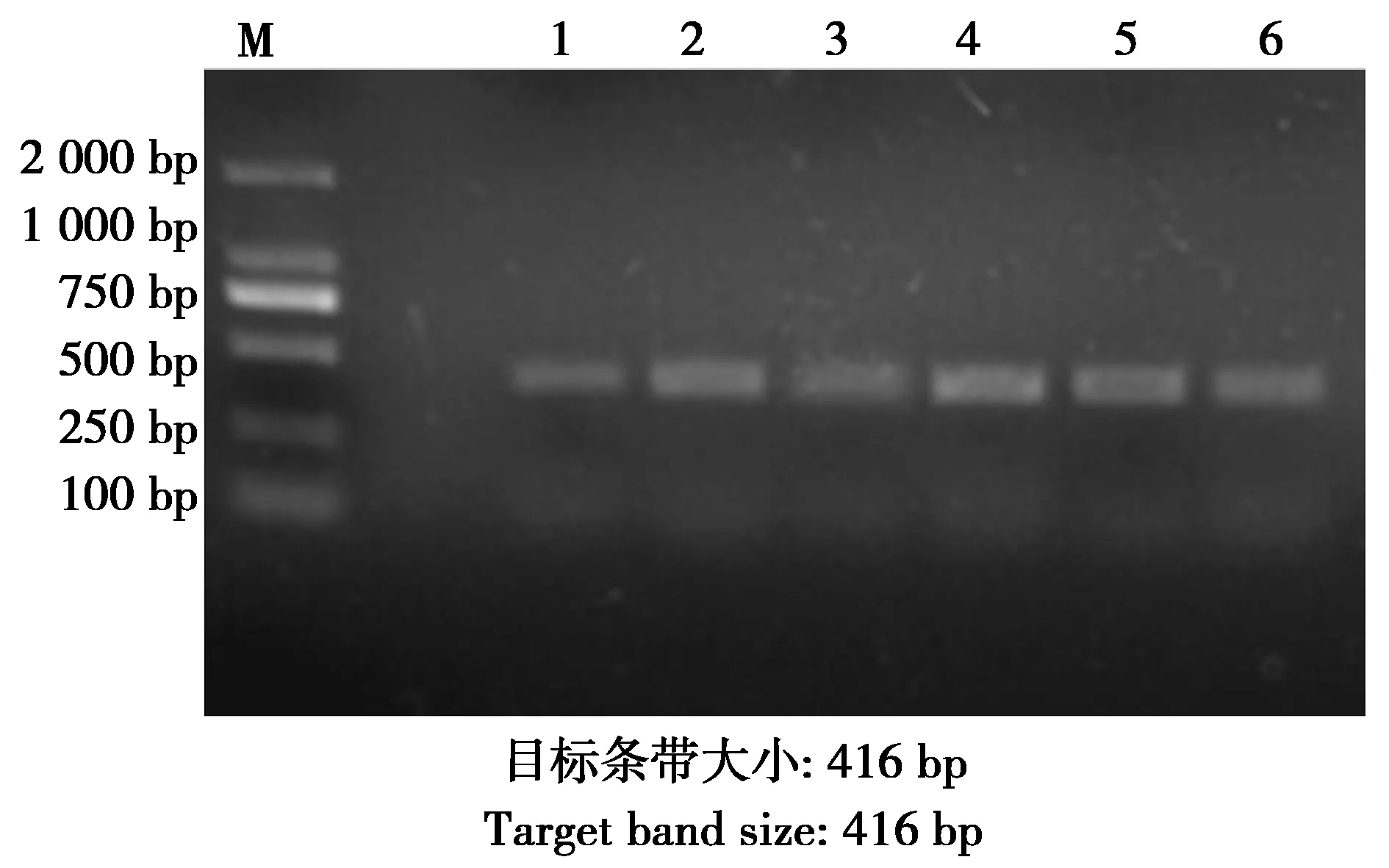
圖2 tetB的PCR擴增結果Fig.2 PCR amplified results of tetB genes 注:M:DNA marker;1—6:tetB耐藥基因的PCR擴增結果 Note:M:DNA marker.1-6:PCR products of tetB gene
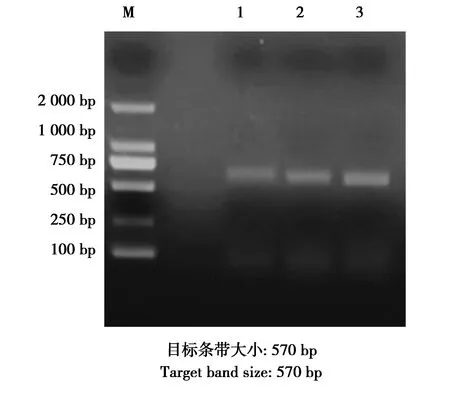
圖3 tetC的PCR擴增結果Fig.3 PCR amplified results of tetC genes 注:M:DNA marker;1—3:tetC耐藥基因的PCR擴增結果 Note:M:DNA marker.1-3:PCR products of tetC gene
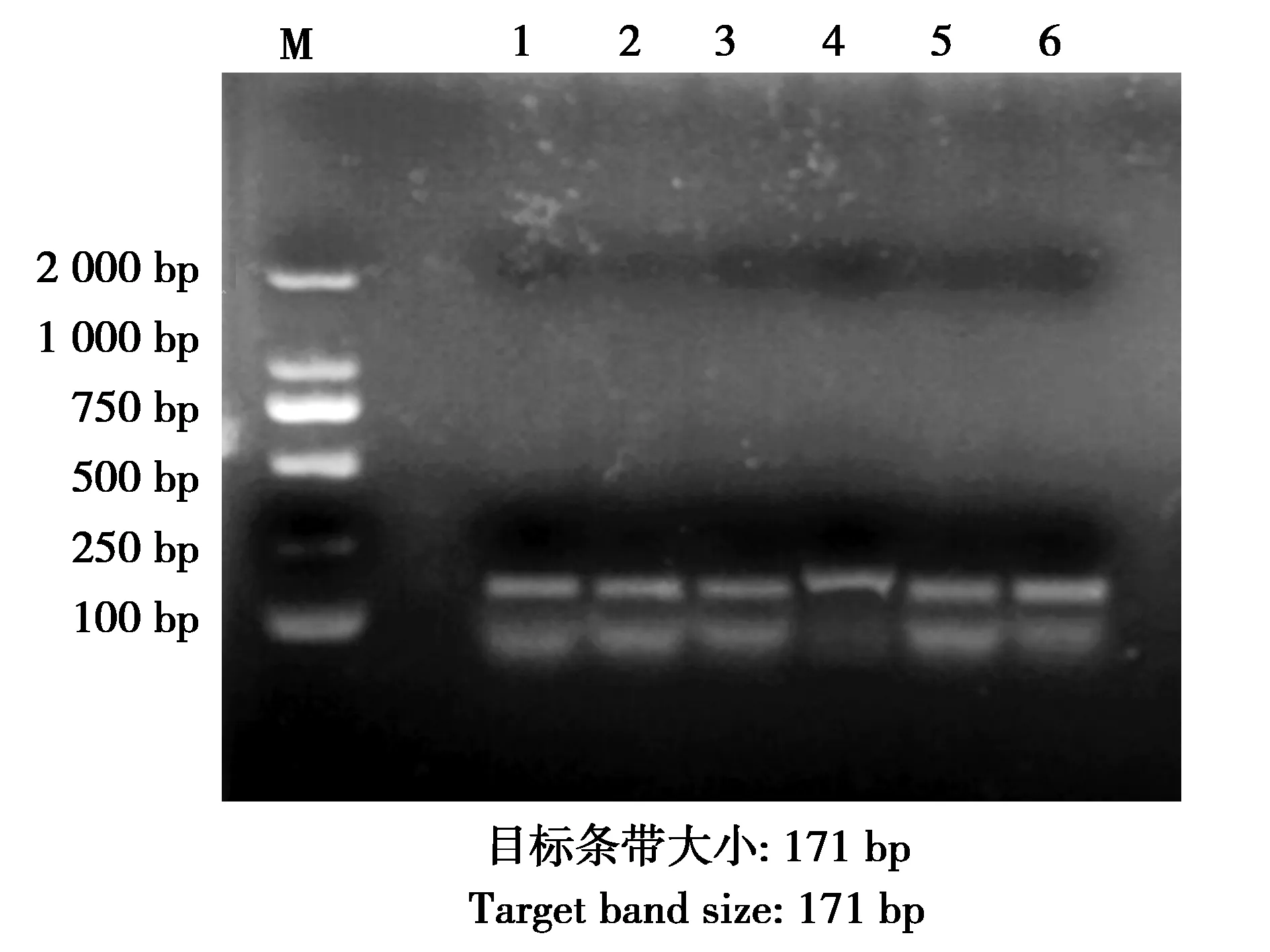
圖4 tetM的PCR擴增結果Fig.4 PCR amplified results of tetM genes 注:M:DNA marker;1—6:tetM耐藥基因的PCR擴增結果 Note:M:DNA marker.1-6:PCR products of tetM gene
4 討論
細菌耐藥性的出現與其耐藥基因的存在及數量密切相關。根據先前黑龍江省和吉林省貉源大腸桿菌耐藥性的檢測,可以得知貉源大腸桿菌對四環素表現出較高的耐藥性,其耐藥率超過60%[8]。因此,基于該結論,本實驗進一步在分子水平檢測了大腸桿菌四環素抗性基因。本實驗結果表明,四環素耐藥性的確與四環素耐藥基因的存在密切相關,尤其是tetM。
一般認為,細菌對四環素產生耐藥性的機制包括4種:外排泵機制,核糖體保護機制,鈍化酶機制,核糖體靶位修飾機制[9]。Tet基因廣泛存在于人源、動物源、共生菌和和環境條件致病菌的不同菌種[9]。大腸桿菌的耐藥性主要基于主動外排機制,也伴有其他耐藥機制,例如tetM基因則是本實驗中核糖體保護機制(四環素類藥物作用的主要作用機制是抑制細菌蛋白質合成,而許多對四環素耐藥的微生物能夠合成一種核蛋白體保護蛋白[9]。從而保證細菌中蛋白質的正常合成,臨床表現為耐藥性。而tetM基因正是這種蛋白的一種編碼基因)。被研究較多的主動外排基因主要是tetA、tetB、tetC等[10]。其中,tetA、tetB、tetC3種基因在本實驗中均有較高的檢出率。但本實驗貉源大腸桿菌中以攜帶tetM基因為主,檢出率高達82.14%。在表明不同來源大腸桿菌中各種抗性基因攜帶狀態有較大的差異。此外,貉的四環素耐藥性可能由tetM基因支配。
研究表明,禽大腸桿菌中,tetA、tetB、tetC、tetM基因也廣泛存在[10],與本實驗大腸桿菌中抗性基因的攜帶狀態一致。此外,在相關的魚類和家禽育種的調查中發現,環境標本中tetM基因檢出率極高,幾乎接近100%。
tetM基因在對四環素耐藥的革蘭氏陽性菌中比較常見。而革蘭氏陰性菌中tetA和tetB較多[11-12](一般認為主動外排機制和核糖體保護機制為四環素產生抗性的主要機制,其中革蘭氏陰性菌主要是主動外排作用,革蘭氏陽性菌以核糖體保護機制為主[13])。2004年Bryan等[14]從雞和豬體內分離的大腸桿菌中克隆得到了tetM基因。而食物源標本的tetM檢出率中,魚類高于豬肉類,豬肉類高于禽類[14]。這也表明了tetM基因宿主具有多種類型和廣泛的范圍。過往研究認為,核糖體保護類抗性基因具有較強的遷移轉化能力,能和可移動元件結合,從而在不同的微生物之間傳播[15]。由于其廣泛的的宿主菌群范圍和強大的遷移轉化能力,tetM基因可以通過宿主細菌為食物鏈和畜群中輕松運輸。含有tetM的基因可以殘存在飼料和食物中,從而進入動物體和人體,因此含有tetM抗性基因的菌群廣泛存在于動物體和人體中。可以推測,吉林省和黑龍江省健康貉中大腸桿菌中檢出tetM最高的原因可能是其飼料中存在著含有tetM的微生物。此外,tetM基因能夠通轉導接合作用在某些病原菌的群體中轉移。該機制的研究對于防止病原菌抗藥性的出現以及擴散具有重要的意義。
同時本試驗檢測出的tetA和tetB基因主要通過主動外排機制介導四環素的抗性[10]。根據先前實驗,在不同地域、物種等檢測出的主動外排基因中,主要以tetA和tetB基因檢出最多,尤其是tetB,并且多重耐藥菌株中tetB基因的檢出率更高。這表明tetB基因在介導多重耐藥有重要的作用[10],也符合本實驗中健康貉大腸桿菌四環素多重耐藥的事實。近年來,國內外大量的報道顯示,tetA、tetB等的遺傳環境中含有許多可自由移動的轉座子[11]。抗性基因可以通過轉座子與其他的一些基因連接。而這些基因可以通過質粒接合而傳遞,在該過程中傳遞抗藥性。可移動元件質粒、整合子[9]、轉座子等有效的介導了耐藥基因的存在和轉移。因此這些流動成分的研究對于四環素乃至很多其他抗生素有著深遠意義,此外,對中國大量細菌耐藥基因的研究也僅止步于基因表型的測定,對于轉座子等移動成分的研究和報道很少。在這方面仍有很多未知的知識需要探索。一旦實現了新的突破,預防耐藥性的出現和預防轉移就可以發揮重要的作用,降低臨床用藥治療和預防的難度。
貉作為經濟動物,其毛皮和膽汁均有著十分廣泛的用途。為了更好地發展養殖業,及時準確地進行流行病學調查非常重要。目前為止,最有效的方法仍然是減少抗生素的使用以避免耐藥的出現。另外飼養環境的衛生檢查也應該符合相應標準,避免由于飼料及畜舍的衛生問題而導致耐藥菌群的擴散。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
保健醫苑(2022年5期)2022-06-10 07:46:38
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
文苑(2018年21期)2018-11-09 01:23:06
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
中國衛生(2015年9期)2015-11-10 03:11:12
中國當代醫藥(2015年17期)2015-03-01 02:03:58
中國衛生(2014年3期)2014-11-12 13:18:12
