從C、N、P化學計量特征分析雷竹氮素克隆整合分株年齡效應
2020-04-21 03:59:38陳雙林時俊帥郭子武何奇江
林業科學研究 2020年2期
關鍵詞:植物
章 超,谷 瑞,陳雙林*,時俊帥,郭子武,劉 軍,何奇江
(1. 中國林業科學研究院亞熱帶林業研究所,浙江 杭州 311400;2. 南京林業大學,江蘇 南京 210037;3. 杭州市余杭區竹業協會,浙江 杭州 311100;4. 杭州市林業科學研究院,浙江 杭州 310016)
克隆植物(Clonal plant)主要指在自然生境條件下,通過營養繁殖的方式自發地產生多個與親代在遺傳基因上完全一致、在形態表現上相同、在生理結構上獨立或者潛在獨立的新個體的植物[1-3]。克隆整合是克隆植物為適應復雜生境而特有的補償機制,基于同一母株的不同分株通過匍匐莖、水平根、地上莖等橫生結構連接在一起,由維管系統沿源—匯梯度實現養分、水分、激素和光合產物等物質的傳輸,實現資源與信息的共享和重新分配[4-6]。植物通過克隆整合作用有助于增強處于弱勢斑塊的分株對異質生境的抗脅迫能力,從而提高整個克隆整合系統的適合度[7-9],較非克隆植物在逆境中的生存與生長、提高種間競爭能力、擴張生境和資源利用等方面有著明顯的優勢[10]。
氮素是植物生長發育必需的營養元素,在植物的生理代謝過程中起著重要作用,同時對植物C的固定和P的吸收產生影響[11]。氮素在生境中存在異質性分布,植物根系在空間上的延伸與拓展體現了其對異質氮素養分的獲取對策,克隆植物也存在著明顯的氮素克隆整合效應[12]。由于克隆植物具有特殊構件性特征,導致其對土壤中氮素的吸收位點呈現多樣性,當氮素一旦被某些分株吸收后,養分傳輸與資源分配的格局和過程呈現復雜化[13]。生態化學計量學是研究生態系統中能量、多種化學元素平衡的科學,主要強調活的有機體內主要元素特別是C、N、P等重要生命元素間的計量關系[14-16]。為了適應生境的變化,植物會主動地調整養分需求,改變體內各種元素的相對豐度[15],維持自身的營養水平從而保證相對較好的生長發育。因此,開展植物體內主要養分化學計量特征對氮素克隆整合的響應機制研究,有助于進一步揭示不同氮素養分條件下克隆植物的生態適應機制。
竹類植物是典型的克隆植物,顯著的克隆整合效應使其能夠提高對異質性生境資源的傳輸效率,加強對外界干擾的抵抗能力,從而實現快速繁殖。目前,對竹類植物分株間克隆整合效應的研究已有一些報道。毛竹(Phyllostachys edulis(Carrière) J.Houz.)林施肥研究表明,當竹鞭穿越養分分布不均的環境時,竹筍有選擇地生長于養分豐富的地段而避開養分貧乏的地段[17]。筇竹(Chimonobambusa tumidissinodaJ. R. Xue et T. P. Yi ex Ohrnb.)無性系分株間通過地下莖的傳輸實現養分資源的共享[18]。美麗箬竹(Indocalamus decorusQ. H. Dai)克隆分株間氮素養分與水分在克隆整合上存在明顯的交互作用[19]。雷竹(Phyllostachys violascens(Carr.) A. et C. Riv. 'Prevernalis')地下鞭根系統盤根錯節,單位面積竹林上有若干個克隆整合單位組成,能夠依靠克隆整合單位在資源、信息等方面進行共享,最大化地利用資源[20]等。已有的研究多涉及水分、養分克隆整合對竹類植物分株間表型可塑性、整合動態、資源分配與養分運輸格局的影響,而不同年齡分株間的養分克隆整合特征差異尚不清楚。為此,本研究以1年生和2年生雷竹組成的克隆整合單元為研究對象,通過竹腔注射硝酸銨方法給不同年齡雷竹添加不同濃度水平的氮素處理,分析各處理的氮素注射與未注射分株葉片C、N、P含量和化學計量比間的差異,試圖回答兩個問題:(1)氮素養分在雷竹分株間是否會發生傳導?如有傳導,對雷竹克隆分株間的養分含量及其化學計量比有無明顯影響?(2)雷竹分株間的氮素克隆整合是否存在分株年齡效應?如果存在,氮素濃度和處理后時間是否會影響這種年齡效應?旨在為雷竹林精準化、減量化施肥提供參考。
1 材料與方法
1.1 試驗地概況
試驗地位于浙江省杭州市余杭區(119°40′~120°23′ E,30°09′~30°34′ N)瓶窯鎮牛頭山,地處天目山系的末端,屬北亞熱帶南緣季風氣候,溫暖濕潤,四季分明,光照充足,雨量充沛,年平均降雨日130~145 d,年平均降水量1 150~1 550 mm,相對濕度74%~85%,年平均氣溫15.3~16.2℃,年平均日照時數1 970 h,年平均無霜期244 d。試驗雷竹林2016年2月新造,坡度15度以下,生長良好,尚未成林,可以明顯區分出由同一母竹萌發而成的克隆整合單元。試驗土壤為紅壤,基本理化性質為:pH值4.46,有機質16.97 g·kg-1,全氮 1.14 g·kg-1,全磷 0.74 g·kg-1,全 鉀 8.47 g·kg-1, 水 解 氮 91.2 mg·kg-1,有 效 磷91.7 mg·kg-1,速效鉀 135.7 mg·kg-1。
1.2 試驗方法
2018年9月,在試驗雷竹林中選取邊界清晰、生長良好、由同一母竹萌發的含2株以上立竹的竹叢為試驗克隆整合單元,采用選擇性伐竹方式,控制整合單元立竹的年齡結構、胸徑和分株距離,構建立竹胸徑基本一致(2.87±0.51)cm、分株距離大致相近(0.73±0.07)m、1年生和2年生立竹各1株的克隆整合單元共18個。
雷竹試驗克隆整合單元構建后,在竹林生理和生長恢復15 d后開展氮素添加試驗。氮素為10.09%豐度的雙標記穩定性同位素15NH415NO3(上海化工研究院生產),分別稱取35、70、105 g15NH415NO3溶解于去離子水中,各配制成容積為210 mL的硝酸銨溶液。用電鉆在雷竹分株距地面5 cm的基部打孔,采用竹腔注射方法注射配制成的不同濃度硝酸銨溶液30 mL·株-1,注射后用膠泥封閉注射孔。氮素添加濃度設置為3個水平,分別為低濃度 N( 4.07 mol·L-1) 、 中 濃 度 N( 8.13 mol·L-1)、高濃度 N(12.20 mol·L-1),各重復3次,1年生和2年生雷竹分株各處理9株。
試驗處理后的第3、6、9天對注射和未注射雷竹分株葉片分別取樣,在樣竹冠層中按照上、中、下部混合取樣方法,采集生長狀況良好的葉片樣品約100 g裝于取樣袋,放入冰盒帶回實驗室。將葉片樣品清洗干凈裝進信封袋,105℃殺青30 min,75℃烘至恒質量,用植物樣品粉碎機粉碎,再用球磨儀(MM400Ball Mill, Retsch, Germany)研磨成細粉狀(<0.5 mm),裝袋儲于真空干燥器中備測。葉片N含量用元素分析儀(Thermo Flash EA1112 Elemental Analyzer)測定,C、P含量分別采用重鉻酸鉀容量法、鉬銻抗比色法測定[21]。
1.3 數據處理與統計分析
試驗數據在Microsoft Excel 2010軟件中進行整理和圖表制作,在SPSS 23.0統計軟件中對不同施N處理和不同處理時間的雷竹葉片各指標進行單因素方差分析(One-way ANOVA),當統計數據方差齊性時選擇Duncan法進行多重比較;當統計數據方差不齊時選擇Dunnett’s T3法進行比較。采用獨立樣本t檢驗判斷1年生和2年生立竹間葉片各指標的差異。相關性分析采用Two-tailed的Pearson相關系數進行分析。
2 結果與分析
2.1 試驗雷竹葉片C、N、P含量
從表1分析可知:1年生立竹不同氮素濃度處理后,葉片C、N含量隨著氮素處理濃度的增大總體呈升高趨勢,且低濃度N、高濃度N處理間差異顯著,葉片P含量則無明顯變化;相連的2年生立竹葉片C、P含量總體上影響不顯著,但對葉片N含量有明顯的影響,且有時間效應,處理后3 d時差異不顯著,處理后6、9 d時均在高濃度N處理下顯著增加。隨著處理時間的延長,不同氮素濃度處理的1年生、2年生立竹葉片C、N、P含量均無明顯變化,1年生立竹葉片N含量在不同氮素濃度處理后3、6、9 d時均顯著高于2年生立竹。由此可見,1年生立竹氮素處理,對葉片N含量有明顯影響,且存在一定的時間效應和氮素濃度效應,1年生立竹會將部分氮素傳輸至2年生分株,但主要是滿足1年生分株自身需要。
從表2分析可知:2年生立竹不同氮素濃度處理后,葉片C、N含量除處理后3 d時變化不明顯外,處理后6、9 d時均在高濃度N處理下顯著升高,葉片P含量隨著氮素處理濃度的增大總體呈“V”形變化趨勢;相連的1年生立竹葉片C、N、P含量總體無顯著變化。隨著處理時間的延長,不同氮素濃度處理的1年生、2年生立竹葉片C、N、P含量總體上均無明顯變化。2年生立竹葉片N含量在低濃度N、中濃度N處理下與1年生立竹無顯著差異,至高濃度N處理時顯著高于1年生立竹。由此可見,2年生立竹氮素處理后,將氮素大量傳輸至1年生立竹,且也存在明顯的時間效應和氮素濃度效應,當氮素處理濃度過高或隨著處理后時間的延長,分株間氮素傳輸能力減弱,試驗的中濃度N是顯著變化的“拐點”。
2.2 試驗雷竹葉片C/N、C/P和N/P
從表3分析可知:1年生立竹不同氮素濃度處理后,葉片C/N、C/P、N/P在處理后3、6、9 d時不同濃度處理間差異均不顯著;相連的2年生立竹葉片C/N、C/P總體上低濃度N處理顯著高于中濃度N、高濃度N處理,葉片N/P隨著氮素處理濃度的增大無顯著變化。隨著處理時間的延長,不同氮素濃度處理的1年生、2年生立竹葉片C/N、C/P、N/P總體上變化均不顯著。隨著氮素處理濃度的增大,2年生立竹葉片C/N在處理后3、6 d時均顯著高于1年生立竹;C/P在低濃度N處理時總體高于1年生立竹,至高濃度N處理時1年生、2年生立竹間差異不顯著;N/P與1年生立竹無顯著差異。由此可見,1年生立竹不同氮素濃度處理對2年生立竹葉片C/N、C/P的提高均有促進作用,但會隨著氮素處理濃度的增大及處理后時間的延長而減弱。
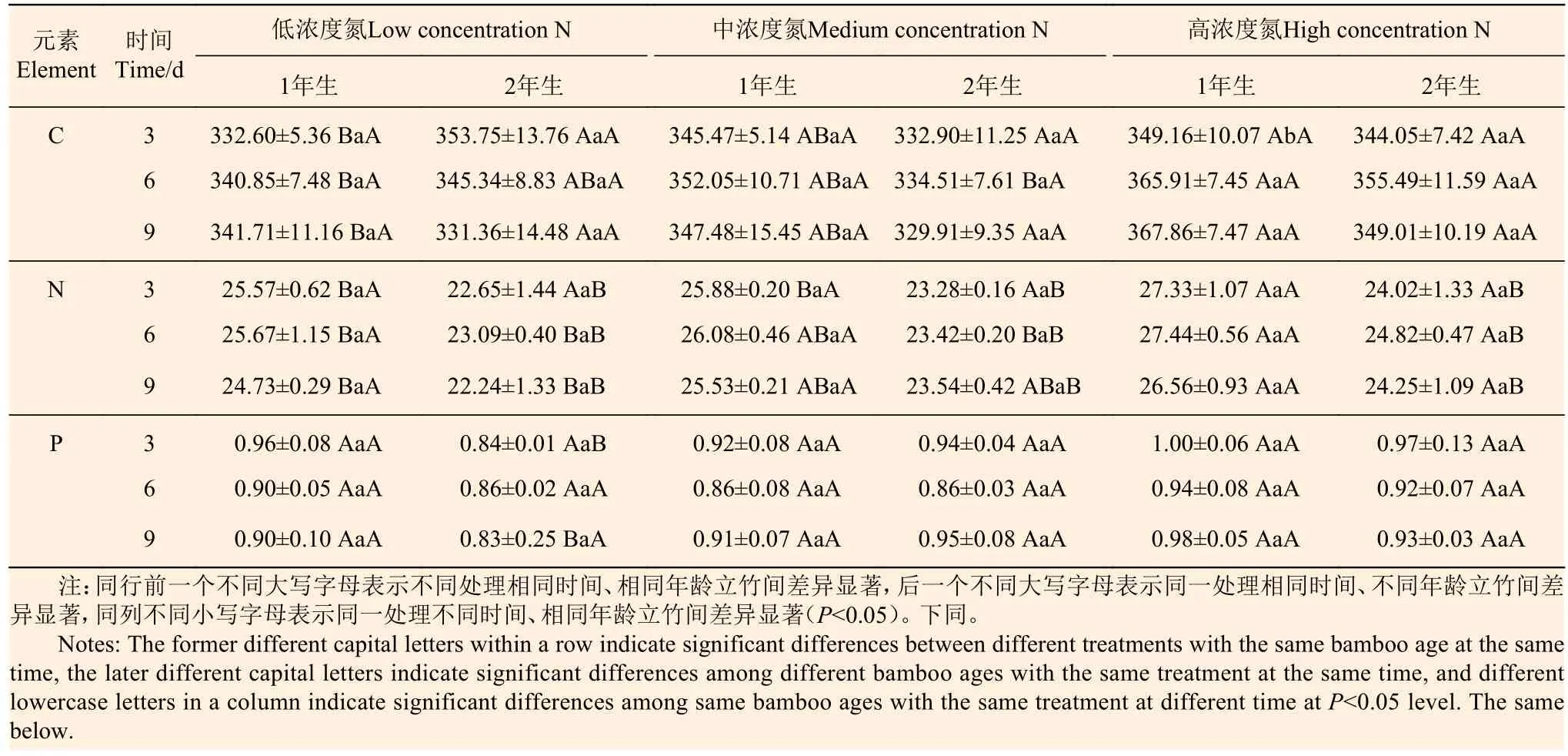
表 1 不同氮素濃度注射處理1年生雷竹立竹葉片C、N、P含量Table 1 Contents of C, N and P in leaves of 1-year-old Ph. violascens with different nitrogen concentrations g·kg-1
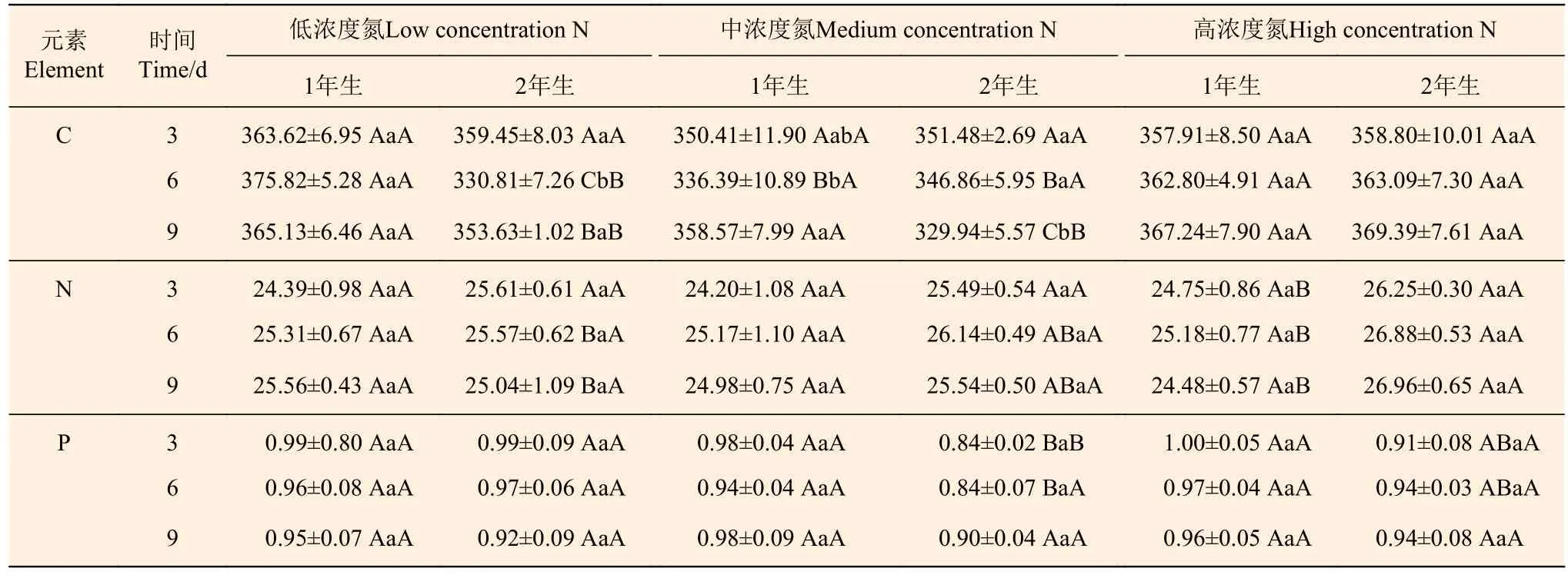
表 2 不同氮素濃度注射處理2年生雷竹立竹葉片C、N、P含量Table 2 Contents of C, N and P in leaves of 2-year-old Ph. violascens with different nitrogen concentrations g·kg-1
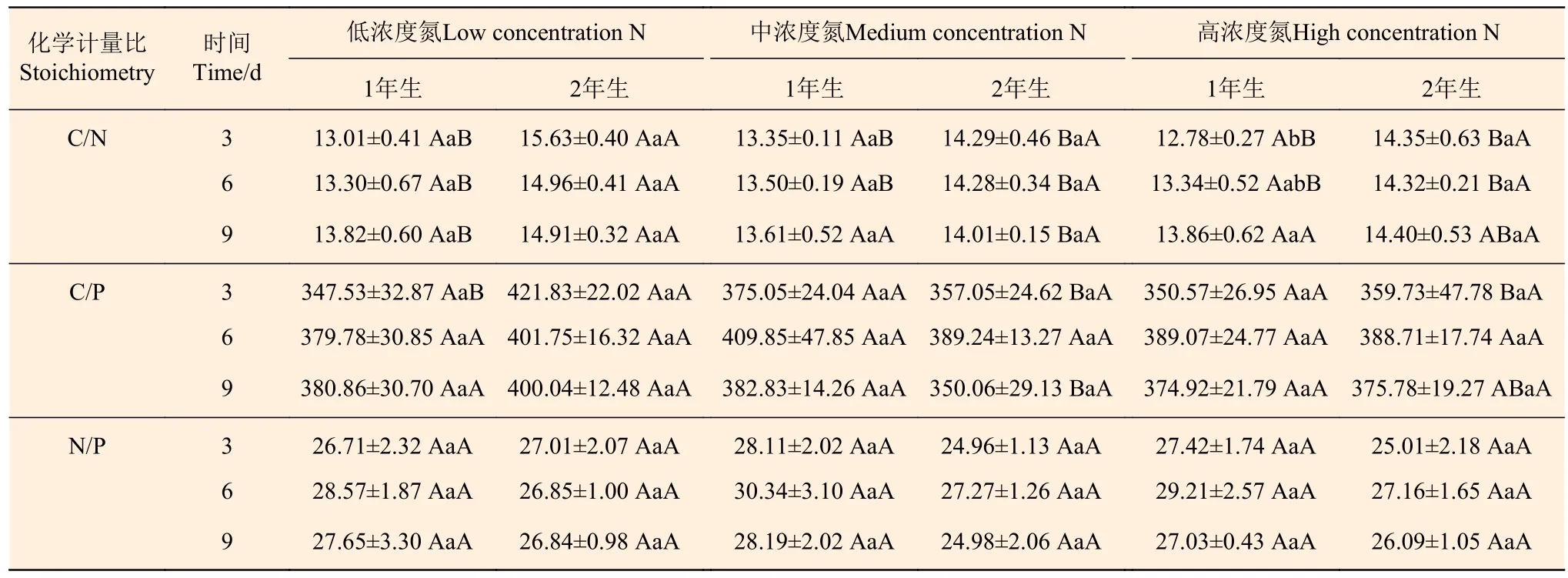
表 3 不同氮素濃度注射處理1年生雷竹立竹葉片C、N、P化學計量比Table 3 C, N and P stoichiometric in leaves of 1-year-old Ph. violascens with different nitrogen concentrations
從表4分析可知:2年生立竹不同氮素濃度處理后,1年生、2年生立竹葉片C/N總體上變化不顯著,中濃度N、高濃度N處理的2年生立竹葉片C/N在處理后9 d時顯著低于1年生立竹;不同氮素濃度處理后3、6、9 d時,1年生、2年生立竹葉片C/P均差異不顯著,且這種變化不隨處理后時間的延長而明顯變化,中濃度N處理的2年生立竹總體顯著高于1年生立竹,其它氮素濃度處理的1年生、2年生立竹間均無顯著差異;不同氮素處理濃度和不同處理時間后1年生立竹葉片N/P差異不顯著,2年生立竹處理后3、6 d時中濃度N處理顯著高于低濃度N處理,而至處理9 d時不同氮素濃度處理間又無顯著差異。隨著氮素處理時間的延長,1年生、2年生立竹葉片N/P變化均不明顯。由此可見,2年生立竹氮素處理對1年生立竹N、P資源利用效率的提高有促進作用,但這種促進作用也隨氮素濃度的增大及處理后時間的延長而減弱,同時保持1年生、2年生立竹葉片N/P的相對穩定。
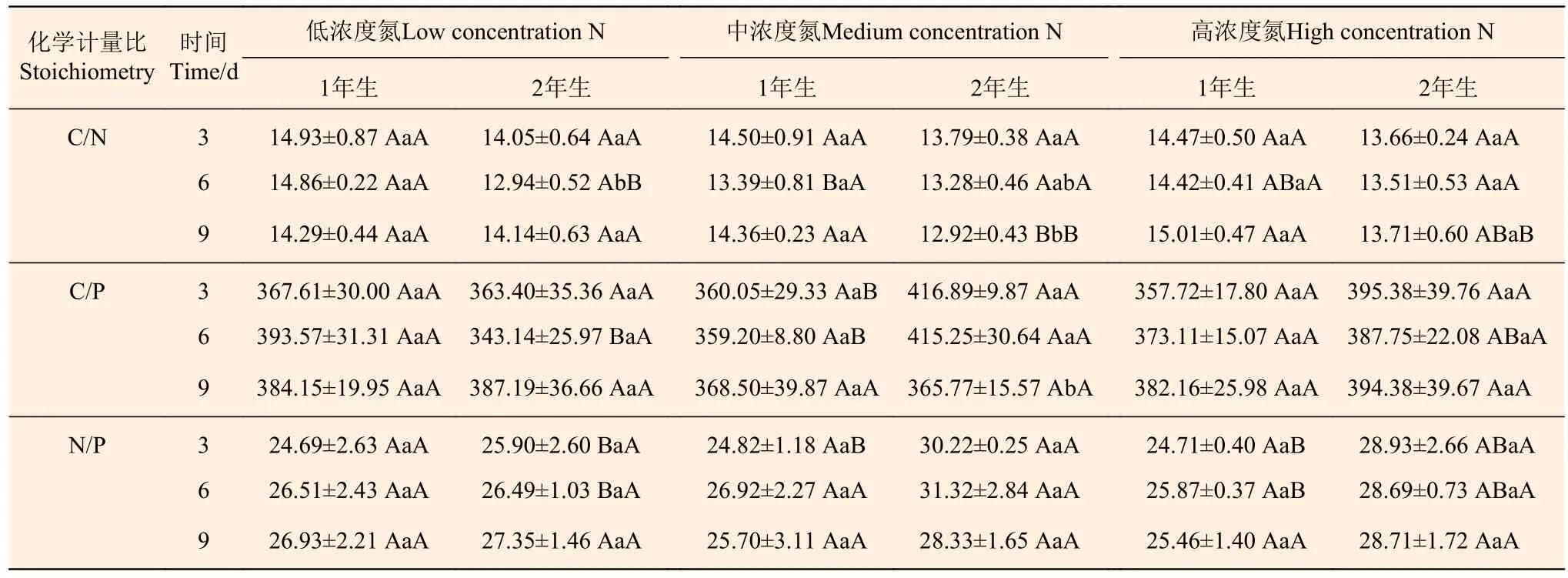
表 4 不同氮素濃度注射處理2年生雷竹立竹葉片C、N、P化學計量比Table 4 C, N and P stoichiometric in leaves of 2-year-old Ph. violascens with different nitrogen concentrations
2.3 試驗雷竹葉片C、N、P含量及其化學計量比間關系
從表5可以看出:1年生雷竹氮素處理后,1年生立竹葉片C含量與N含量呈極顯著正相關(P<0.01),與相連的2年生立竹葉片N含量也呈極顯著正相關(P<0.01),而與P含量、N/P以及2年生立竹葉片C、P含量和C/N、C/P、N/P相關性不顯著;1年生立竹葉片N含量與2年生立竹葉片N含量呈顯著正相關(P<0.05),而與P含量、C/P以及2年生立竹葉片C、P含量和C/N、C/P、N/P相關性不顯著;1年生立竹葉片P含量與C/N以及2年生立竹葉片C、N、P含量和C/N、C/P、N/P相關性均不顯著。2年生立竹葉片C含量與N含量呈極顯著正相關(P<0.01),而與P含量、N/P相關性不顯著,N含量與P含量呈極顯著正相關(P<0.01),而與C/P相關性不顯著,P含量與C/N呈極顯著負相關(P<0.01),C/N與C/P、C/P與N/P均呈極顯著正相關(P<0.01)。
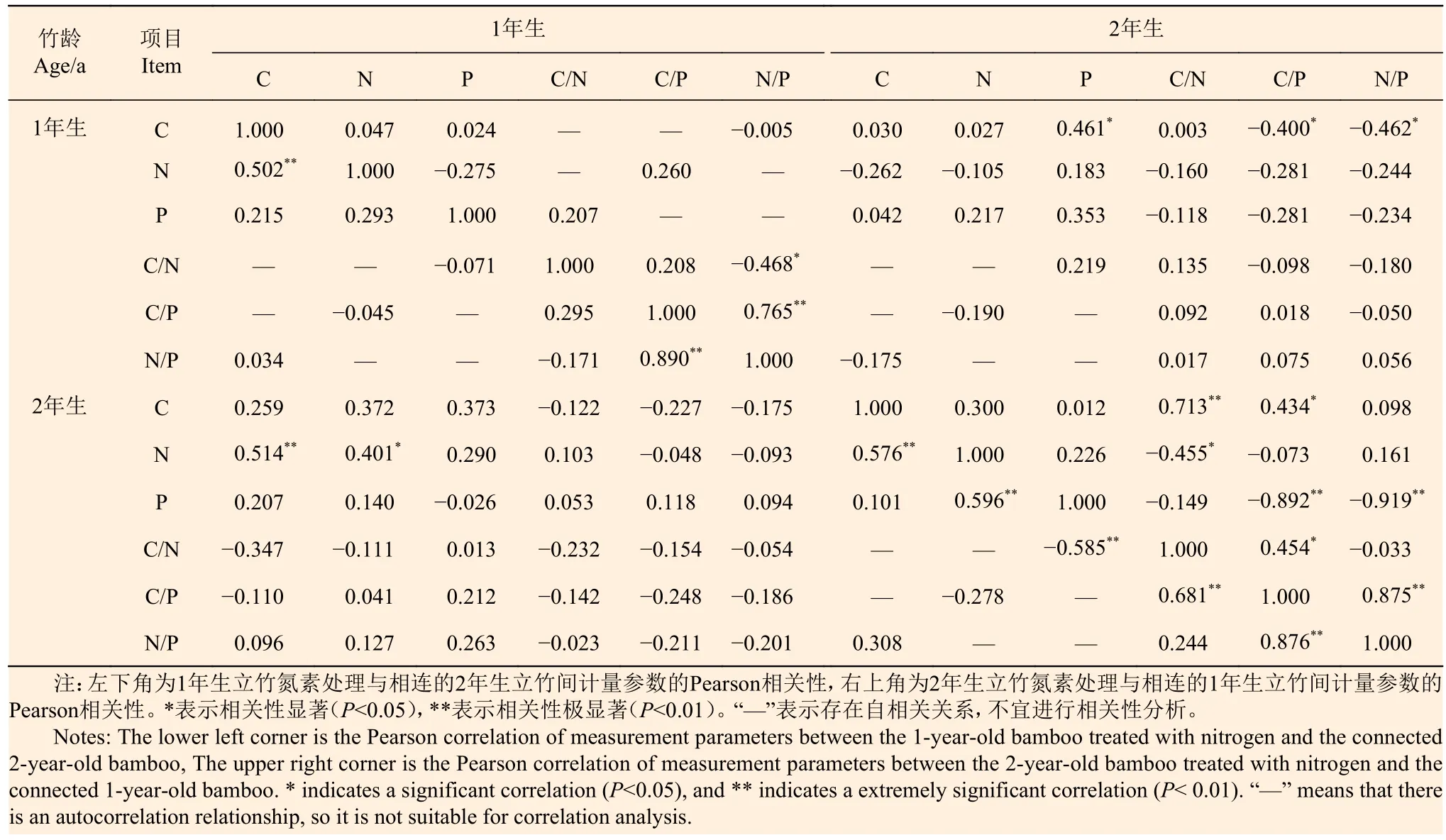
表 5 不同年齡雷竹氮素添加處理立竹葉片C、N、P含量及化學計量比的相關性Table 5 Correlations of C, N, P contents and stoichiometric in leaves of Ph. violascens treated with nitrogen addition at different ages
2年生雷竹氮素處理后,1年生立竹葉片C含量與N、P含量和N/P以及2年生立竹葉片C、N含量和C/N相關性不顯著,而與2年生立竹葉片P含量呈顯著正相關(P<0.05),與2年生立竹葉片C/P、N/P呈顯著負相關(P<0.05);1年生立竹葉片N含量與P含量、C/P以及2年生立竹葉片C、N、P含量和C/N、C/P、N/P相關性不顯著;1年生立竹葉片P含量與C/N以及2年生立竹葉片C、N、P含量和C/N、C/P、N/P相關性均不顯著。2年生立竹葉片C含量與N、P含量和N/P相關性不顯著,N含量與P含量、C/P相關性不顯著,P含量與C/N相關性不顯著,C/N與C/P、C/P與N/P分別呈顯著和極顯著正相關。由此可見,對于雷竹1年生、2年生立竹氮素的克隆傳輸,克隆系統分株會發生一系列的葉片C、N、P含量及其化學計量比關系的適應性調節。
3 討論
從多數克隆植物的克隆整合格局研究中發現,分株年齡是決定克隆分株間源—匯關系的重要因素[22-25]。在本研究中,雷竹1年生立竹氮素濃度處理后,通過克隆整合單元的傳輸,相連的2年生立竹葉片N含量隨著氮素處理濃度的增大而升高,但1年生立竹葉片N含量在3個氮素處理濃度上均顯著高于2年生立竹,而2年生立竹氮素濃度處理后,1年生、2年生立竹葉片N含量在低濃度N、中濃度N處理時無顯著差異,至高濃度N處理時2年生立竹顯著高于1年生立竹。這說明雷竹克隆整合系統存在明顯的分株年齡效應,2年生立竹會將更多的氮素養分傳輸給1年生立竹,而1年生立竹僅將小部分氮素養分傳輸給2年生立竹。分析原因,一方面可能是1年生立竹獲取氮素養分后更多是滿足自身體積和質量生長的需求,存在明顯的“利已”行為,而2年生立竹在滿足自身養分需求的同時,會將氮素大量傳輸至幼齡分株,從而提高雷竹克隆整合單元的適合度,存在著明顯的“利他”行為;另一方面也可能與1年生植株對氮的響應較敏感[26-27]等有關。
本研究中,雷竹1年生立竹氮素濃度處理后,1年生立竹葉片C含量隨氮素處理濃度的增大呈升高趨勢,且在高濃度N處理下顯著增加,相連的2年生立竹葉片C含量總體呈先降低后升高的趨勢,說明氮素添加在一定程度時會明顯促進雷竹1年生立竹C固定能力,這與吳福忠等對白刺花(Sophora davidii(Franch.) Skeels)施氮研究結果相一致[28]。2年生立竹氮素濃度處理后,2年生立竹葉片C含量僅在高濃度N處理下顯著增加,相連的1年生立竹葉片C含量在低濃度N處理下顯著高于2年生立竹,其原因可能是2年生立竹在低濃度N處理下“利他”行為更為明顯,優先將氮素養分傳輸至1年生分株,從而強化1年生立竹葉片C固定能力。本研究中,氮素添加對雷竹1年生和2年生立竹葉片P含量均無明顯影響,和前人的研究結果基本一致[29-31],說明氮施加量對雷竹P的吸收影響不大。植物葉片的C/N、C/P意味著植物吸收營養所能同化C的能力,在一定程度上反映植物的養分利用效率,因而,具有重要的生態學意義[15]。本研究發現,雷竹1年生立竹氮素濃度處理后,相連的2年生立竹葉片C/N均顯著高于1年生立竹,葉片C/P在低濃度N處理下總體高于1年生立竹,葉片N/P在不同氮素濃度和不同處理時間均無明顯變化,且隨著1年生立竹氮素濃度的增大以及處理時間的延長,2年生立竹葉片C/N、C/P呈減小的趨勢;2年生立竹氮素濃度處理后,相連的1年生立竹葉片C/N總體均高于2年生立竹,葉片C/P在低濃度N處理總體高于2年生立竹,葉片N/P在高濃度N處理下總體顯著低于2年生立竹,隨著2年生立竹氮素濃度的增大,1年生立竹葉片C/N、C/P總體也呈減小的趨勢。說明1年生、2年生立竹氮素處理均能提高相連分株葉片N、P的養分利用效率,且隨氮素處理濃度的增大及處理時間的延長而減弱,同時維持相連立竹葉片N/P的相對穩定。但2年生立竹氮素濃度處理后,隨氮素處理濃度的升高,2年生立竹葉片N/P明顯增大,受P限制作用可能會進一步增強[32]。對1年生、2年生立竹氮素處理與相連不同年齡立竹間計量參數的相關性分析表明,1年生立竹氮素濃度處理后,1年生立竹葉片C含量與N含量呈極顯著正相關,與相連的2年生立竹葉片N含量也呈極顯著正相關,說明氮素添加在一定程度時會明顯促進雷竹1年生立竹C固定能力。另外,1年生立竹氮素添加處理后,1年生立竹葉片N含量與相連的2年生立竹葉片N含量呈顯著正相關,而2年生氮素添加處理后,2年生立竹葉片N含量與相連的1年生立竹葉片N含量呈負相關,進一步說明2年生立竹會將更多的氮素養分傳輸給相連的1年生立竹,表現為“利他”行為,且在氮素養分充足的情況下,這種氮素傳輸能力也明顯減弱。
4 結論
本研究發現,雷竹氮素克隆整合特性存在明顯的分株年齡效應,1年生立竹會將部分氮素傳輸至2年生立竹,但主要是滿足自身生長發育的需要,表現為“利已”行為,而2年生立竹將氮素大量傳輸至1年生立竹,表現為明顯的“利他”行為。1年生、2年生立竹在傳輸氮素養分時,均存在一定的時間效應和氮素濃度效應,當氮素處理濃度過大或處理后時間過長時,這種傳輸能力減弱,試驗的中濃度N是顯著變化的“拐點”。1年生、2年生立竹氮素處理均會對相連的立竹N、P資源利用效率的提高有促進作用,但會隨氮素濃度的增大呈減弱趨勢,同時維持1年生、2年生立竹葉片N/P的相對穩定性,并使雷竹葉片C、N、P含量及其化學計量比關系產生適應性調節作用。在雷竹林經營中,宜選擇2年生立竹采取株穴法施入氮肥,可以顯著地提高肥料的利用率,并實現肥料使用的精準化和減量化。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
