外來入侵物種紅耳龜咬合力性二型研究
2020-04-14 01:42:30李漢東王同亮胡振義饒靜秋史海濤汪繼超
四川動物 2020年2期
李漢東, 王同亮, 胡振義, 饒靜秋, 史海濤, 汪繼超
(熱帶島嶼生態學教育部重點實驗室,海南師范大學生命科學學院,海口571158)
兩性異形是指同一物種雌性和雄性個體間體型、形態、體色等方面的差異(Shine,1979;何南等,2011),反映了兩性間不同的適應性。研究發現,兩性異形普遍存在于魚類(Ijirietal.,2008;樊曉麗等,2016)、兩棲類(施林強等,2011;Wangetal.,2016)、爬行類(Berry & Shine,1980;肖繁榮等,2014)、鳥類(Selander,1966;Owens & Hartley,1998)和哺乳類(Clutton-Brocketal.,1977;Ralls,1977)等動物類群中。目前龜鱉類動物的兩性異形也開展了較多的研究,龜鱉類動物成體的體型存在3種兩性異形模式:1)雌性大于雄性,如四眼斑水龜Sacaliaquadriocellata(肖繁榮等,2014)、黃喉擬水龜Mauremysmutica(Gosnelletal.,2019);2)雄性大于雌性,如挺胸陸龜Chersinaangulata(Gosnelletal.,2019);3)雌雄個體之間無顯著差異,如日本擬水龜M.japonica(Gosnelletal.,2009)。因此龜鱉類動物體型大小的兩性異形無統一模式(Coxetal.,2007),且具有物種特異性。
咬合力體現了動物的生態適應性,直接影響動物捕獲、處理及攝取食物的效率(Ericksonetal.,2003;Christiansen & Wroe,2007;Huberetal.,2008)。目前國內外對咬合力開展了較多的研究工作,主要發現咬合力與動物的頭部形態、食性等因素有關。頭部形態能反映咬合力:通常頭部越大個體的咬合力越大(Herreletal.,2002;Herreletal.,2005a,2005b;Sustaitaetal.,2014)。此外,肉食性動物的咬合力大于植食性、雜食性動物(Herreletal.,2002),且咬合力大的動物取食效率也較高(Herreletal.,2001;Verwaijen & Herrel,2002;der Meij & Bout,2006)。然而,這方面的研究主要集中于同一動物類群的種間比較(Herreletal.,2002;霍娟等,2016;譚梁靜等,2017)、同一物種不同發育階段的比較(Ericksonetal.,2003;Herrel & O’Reilly,2006;Pfalleretal.,2010;Marshalletal.,2012)。目前同一物種雌性和雄性個體間的咬合力是否存在性二型的研究相對較少,且結論不一(霍娟等,2016;Herreletal.,2018)。因此,動物的咬合力性二型及其與形態的關系仍需進一步研究。
紅耳龜Trachemysscriptaelegans又名巴西龜,原產于美國密西西比河至墨西哥灣周圍地區(Gibbons,1990);20世紀80年代,該物種經香港入侵我國內陸地區,野外分布區涉及22個省市(史海濤等,2009)。雌性紅耳龜成體的體型顯著大于雄性(Gibbons & Lovich,1990),體色(汪繼超等,2013)、聽力敏感性(Wangetal.,2019)也存在兩性異形。紅耳龜為雜食性動物,食域范圍廣泛,雌雄之間對動、植物性食物的取食頻次和比重均無顯著差異(Wangetal.,2013)。研究發現,紅耳龜咬合力發育存在可塑性,成體咬合力顯著大于幼體(Herrel & O’Reilly,2006)。目前紅耳龜成體咬合力是否存在兩性異形及其與形態的關系尚不清楚。本研究采用力值傳感器量化了紅耳龜的咬合力,分析了咬合力可能的性別差異及其與體型大小兩性異形的關系,以期為理解該物種的生態適應性提供參考。
1 材料與方法
1.1 實驗材料
2015年8—10月在海南省文昌岸興龜類養殖專業合作社購買紅耳龜成體,飼養于海南師范大學生態園龜鱉生態養殖池內(200 cm×150 cm×120 cm),水深高過紅耳龜的背甲約5 cm,每隔3 d飼喂龜糧(海豚水族有限公司)1次,當日取出剩余食物,清洗養殖箱并換水。馴化約1周后,隨機選取30只健康個體進行實驗。
1.2 咬合力測定
咬合力采用咬合力測量儀器(南京神源生智能科技有限公司)測定。該設備主要包括咬合力傳感器(S2-500NHL-001,0~500 N/0.001 N)、模擬數據采集卡(NBIT-DUS-2404A)和信號采集軟件(DSU2404A)三部分,其工作原理是模擬數據采集卡將作用在傳感器上的力值信號轉換成電信號,然后通過信號采集軟件將力值的變化以波形圖的形式顯示出來。測量時,一手托住紅耳龜腹部,另一只手輕放在紅耳龜背甲上,將其頭部靠近咬合力傳感器的探頭,等待紅耳龜主動咬合。每只紅耳龜重復測量7次,去掉7次數據的最大值和最小值,剩余的5次數據的平均值作為每只紅耳龜的咬合力。
1.3 形態特征測量
采用電子秤(莫林貿易有限公司;30 kg/1 g)稱量體質量。采用數顯游標卡尺(香港五羊工具有限公司;0~300 mm/0.01 mm)測量背甲長(頸盾前緣至臀盾后緣最大直線長度)、頭長(吻端至上、下頜關節后緣的直線距離)、頭寬(頭左、右兩側間最大距離)和頭高(頭上、下兩側間最大距離)。并計算頭部大小=(3.141 5×頭長×頭寬×頭高)/6(Macías-Ordóez & Draud,2005)。
1.4 數據分析
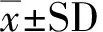
2 結果
2.1 紅耳龜的咬合力
雌性紅耳龜(n=16)的咬合力(71.92 N±15.59 N)顯著大于雄性(n=14)的(55.24 N±10.09 N)(P=0.047)(圖1)。
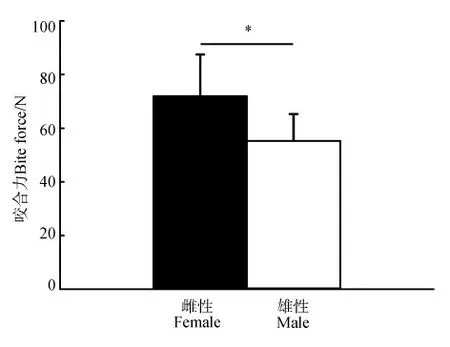
圖1 紅耳龜雌性和雄性個體間咬合力比較Fig. 1 Comparison of bite force between female and maleTrachemys scripta elegans
*P<0.05
2.2 紅耳龜的形態指標
雌性紅耳龜的體質量(F=10.360,P=0.003)、頭部大小(F=6.850,P=0.014)、背甲長(t=2.951,P=0.008)、頭寬(F=10.614,P=0.003)均顯著大于雄性,而頭長、頭高在雌性和雄性個體間的差異無統計學意義(P>0.05)(表1)。
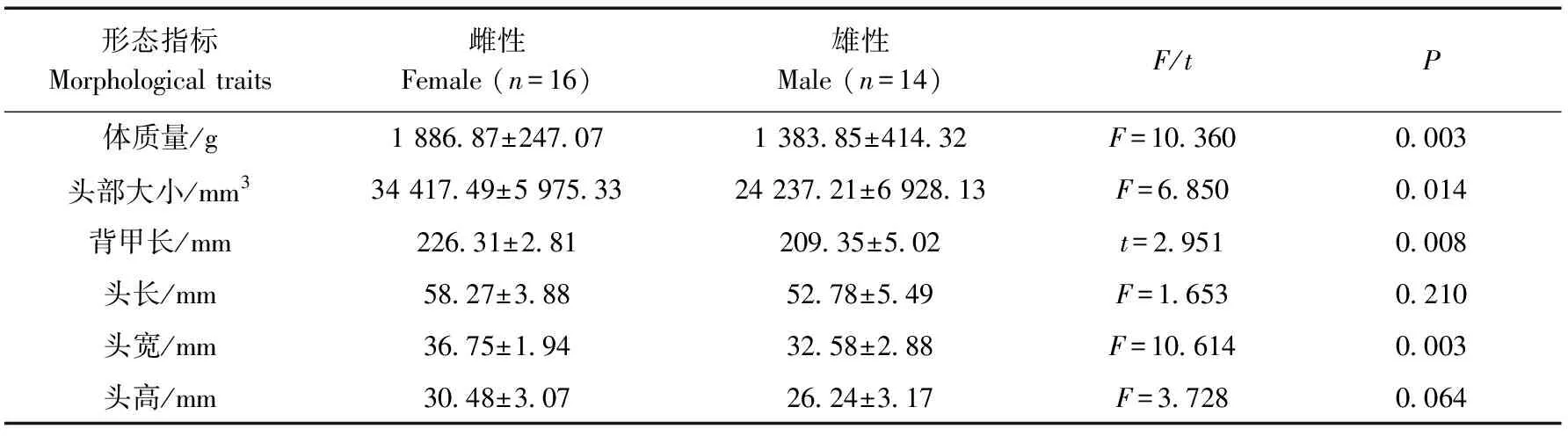
表1 紅耳龜雌性和雄性個體的形態特征Table 1 The morphological traits in female and male Trachemys scripta elegans
2.3 紅耳龜咬合力與形態指標間的線性關系
紅耳龜的咬合力與體質量(R2=0.437,F1,28=21.693,P=0.002)、頭部大小(R2=0.444,F1,28=22.319,P<0.001)、背甲長(R2=0.396,F1,28=18.356,P=0.001)、頭長(R2=0.312,F1,28=12.691,P=0.002)、頭寬(R2=0.515,F1,28=29.724,P=0.001)、頭高(R2=0.380,F1,28=17.164,P=0.001)均呈顯著的線性關系(表2)。
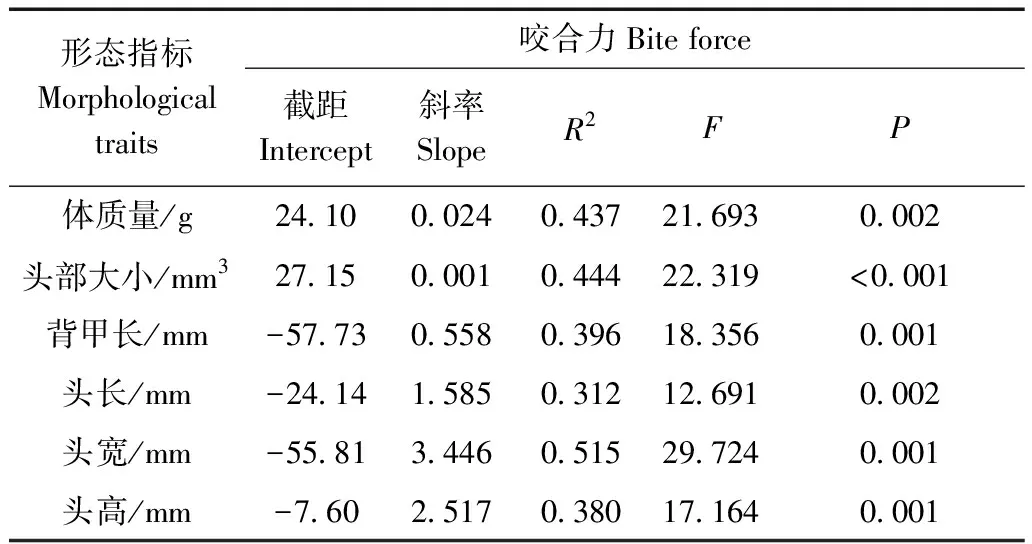
表2 紅耳龜咬合力與形態指標的線性回歸分析Table 2 Regression analysis between the bite force andmorphological traits of Trachemys scripta elegans
3 討論
本研究測定了30只紅耳龜成體的咬合力,其大小為63.96 N±15.27 N。Herrel等(2002)報道了紅耳龜的咬合力為14.59 N±18.76 N,這小于本研究的測定結果。分析發現,Herrel等(2002)測定的紅耳龜個體相對較小,背甲長僅為88.77 mm±53.26 mm,而本研究中紅耳龜的背甲長為218.40 mm±17.24 mm。此外Herrel和O’Reilly(2006)測定了包括幼體和成體在內的 32只紅耳龜的咬合力,范圍為1.02~104.14 N,且成體咬合力大于幼體。本研究測定的咬合力結果與Herrel和O’Reilly(2006)測定的紅耳龜成體的咬合力結果相似。因此,紅耳龜咬合力受體型大小的影響。
本研究發現,雌性紅耳龜成體的咬合力顯著大于雄性,表明該物種的咬合力存在性二型。此外紅耳龜的咬合力與頭部形態顯著相關,且雌性個體的頭部大小、頭寬均顯著大于雄性。因此,頭寬的性別分化引起了該物種咬合力的兩性異形。這與Herrel等(2018)對菱斑龜Malaclemysterrapin咬合力與頭部形態關系的研究結論相似。在鳥類研究中也發現,個體的頭越大、喙越寬,其咬合力也越大(Herreletal.,2005a,2005b;Sustaita & Rubega,2014)。頭部空間增大可以為牽引肌提供更大的附著面,從而引起咬合力的增大(der Meij & Bout,2004;Herreletal.,2005a)。
目前性選擇壓力、生育力選擇壓力和生態位分離假說可解釋動物兩性異形的形成(Shine,1989;Olssonetetal.,2002;廖灝泓等,2013)。本研究發現,雌性紅耳龜體型顯著大于雄性。生育力選擇壓力假說認為,雌性體型的增大有利于增加繁殖輸出(Olssonetal.,2002;Kupfer,2009;Luoetal.,2012),這也導致雌性個體需要攝取更多的食物以滿足能量需求。咬合力與動物處理食物的時間有關,咬合力大的個體取食效率提高(Herreletal.,2001;Verwaijen & Herrel,2002;der Meij & Bout,2006)。因此,雌性紅耳龜采用增大咬合力的策略,提高自身的取食效率,以滿足大體型及繁殖的能量需求,從而提高自身的適合度。此外,繁殖期的雌性紅耳龜需要在陸地上完成巢址選擇、筑巢、產卵等活動,更大的咬合力可能有助于應對更為復雜的陸地環境。
生態位分離假說認為,動物通過兩性異形以適應不同的生態位(Slatkin,1984;Shine,1989),從而降低了捕食壓力(Aguirreetal.,2002;Herreletal.,2004)。Herrel等(2018)報道了菱斑龜的咬合力性二型引起了雌性和雄性個體食物結構的分化。本研究發現紅耳龜的咬合力存在性二型,但已有研究發現雌雄性個體間對取食食物的頻次和比重沒有顯著差異(Wangetal.,2013)。進一步分析發現,菱斑龜的食性專一,種內的食物競爭壓力較大,食物結構的分化有利于減少食物競爭壓力,而紅耳龜為雜食性動物且食域范圍廣泛,雌雄之間食物競爭壓力相對較小。因此,動物咬合力的性二型與食性分化可能與環境食物的豐富度有關。在個體發育過程中,紅耳龜咬合力性二型與頭部形態性別分化的時間關系值得進一步研究。
致謝:海南師范大學生命科學學院郭蕊老師、肖繁榮老師在數據處理和文章修改過程中給予幫助;海南師范大學本科生黃穎和吳迪在飼養實驗動物和形態指標測定過程中給予幫助;南京神源生智能科技有限公司王耀技術員在設備調試和數據分析過程中給予幫助,在此一并表示感謝。
