FaesAP2B基因在甜蕎長雌蕊長雄蕊突變體lpls的表達分析
2020-04-08 08:21:22張良波劉志雄
植物研究 2020年2期
張 嬌 王 旋 張良波 劉志雄
(長江大學園藝園林學院,荊州 434025)
甜蕎(FagopyrumesculentumMoench.)為蓼科(Polygonaceae)蕎麥屬(FagopyrumMill)食藥同源的雜糧作物,是蕎麥屬中2個栽培種之一[1]。甜蕎籽粒除含大量蛋白質、纖維素、脂肪、淀粉等營養成分外,還含硒、銅等礦物質元素及蘆丁等生物活性成分[2~3],營養價值高,且具保健功效,現越來越受人們的青睞。
甜蕎屬特殊異花授粉植物,自然群體中pin型和thrum型花植株1∶1分離,同型花自交不親和,異型花間才能正常授粉結實,人工雜交困難,產量低[4~5]。尋找同型花和異型花間均能正常授粉結實的甜蕎新種質,對于開展甜蕎雜交育種、提高甜蕎產量、新品種培育等均具有重要的理論意義和實際應用價值。課題組在前期育種實踐中,從甜蕎品種“北早生”群體中發現了長雌蕊長雄蕊“lpls”(Long pistil and long stamen,lpls)自然變異的單株,通過隔離授粉和單粒傳法獲得了遺傳性狀穩定、親和性好、同型花、異型花間均能正常授粉結實的自然變異株系,成為開展甜蕎育種工作的重要種質資源(圖1)。深入系統研究甜蕎lpls突變體花、籽粒發育的過程與分子調控機制,對于該種質的可持續性利用有重要的意義。

圖1 甜蕎3種花型圖 A.thrum型花(短花柱長雄蕊);B.pin型花(長花柱短雄蕊);C.lpls突變體花(長花柱長雄蕊)Fig.1 Three flower types of F.esculentum A.thrum flower with short pistil and long stamen; B.pin flower with long pistil and short stamen; C.lpls flower with long pistil and long stamen
在模式植物擬南芥中,AP2基因主要參與花器官和果實的發育調控,促進花分生組織形成[6~8]。本實驗以甜蕎長雌蕊長雄蕊突變體lpls為材料,通過同源克隆結合RACE技術從甜蕎突變體lpls花芽中克隆出了1個甜蕎AP2同源基因的cDNA序列,在分析其結構的基礎上,通過實時熒光定量PCR技術(quantitative Real-time PCR,qPCR)檢測其在甜蕎突變體lpls不同營養組織和生殖結構中的表達,以期為后續驗證該基因在調控甜蕎花發育的生物學功能上提供一定實驗基礎,并為進一步分析甜蕎籽粒及花發育分子機制,及遺傳改良積累資料。
1 材料與方法
1.1 實驗材料
2018年3月21日,挑選顆粒飽滿的甜蕎‘lpls’的籽粒播種于花盆中(21 cm×14 cm×20 cm)。待5月盛花期時,分別從不同植株(≥3)上采集根、莖、幼葉、同時剝離花被片、雄蕊和雌蕊,以及發育4 d的甜蕎果實于液氮中速凍后置于-80℃冰箱保存備用。
1.2 實驗方法
1.2.1 甜蕎lpls型花植株總RNA的提取及cDNA合成
取100 mg左右植物材料采用EASY spin植物總RNA提取試劑盒(艾德萊北京)分別提取甜蕎的根、莖、幼葉、花被片、雄蕊、雌蕊和發育4 d的果實的RNA。采用HiScriptⅡ 1st Strand cDNA Synthesis Kit(諾唯贊南京)試劑盒反轉錄合成第一鏈cDNA,方法參照說明書。
1.2.2 甜蕎FaesAP2B基因的克隆
根據NCBI(https://www.nlm.nih.gov/)數據庫中公布的本課題組前期克隆的甜蕎FaesAP2基因序列(KM386628.1),在其5′非翻譯區(5′UTR)設計克隆FaesAP2B基因的特異性擴增引物GSPAP2B,以甜蕎花芽cDNA為模板,用3′-Full RACE Core Set Ver.2.0試劑盒(TaKaRa)擴增甜蕎FaesAP2B基因的cDNA序列全長,擴增PCR程序為:95℃預變性3 min,95℃變性30 s,58℃退火30 s,72℃延伸60 s,30個循環,72℃延伸10 min。擴增產物經1%瓊脂糖凝膠電泳分離、回收目的片段,經連接、轉化、陽性克隆鑒定、測序獲得甜蕎FaesAP2B基因的全長序列,實驗所用引物見表1。
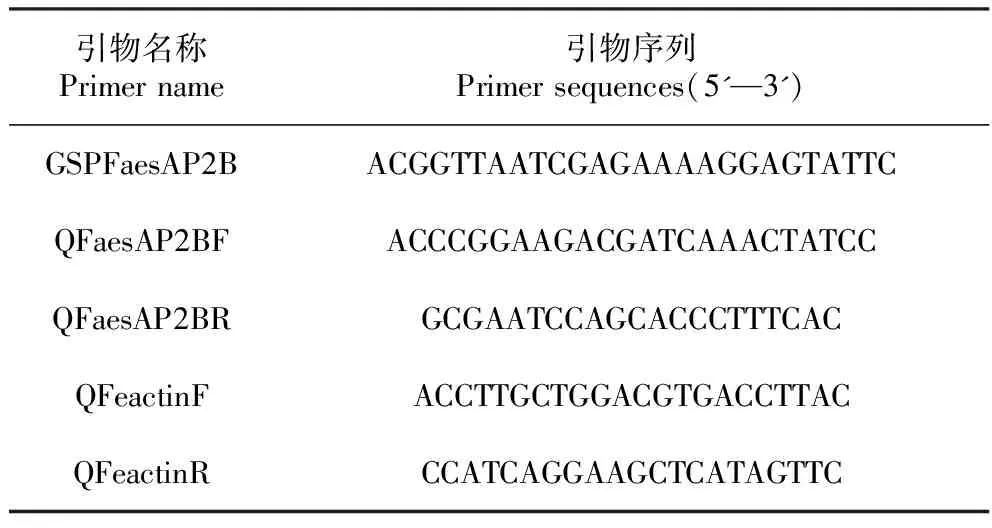
表1 引物名稱及序列
1.2.3 蛋白序列比對與分子系統發育分析
將甜蕎FaesAP2B基因開放閱讀框(Open Reading Frame,ORF)編碼的氨基酸序列在NCBI(https://www.ncbi.nlm.nih.gov/)數據庫中進行BLAST同源搜索(https://blast.ncbi.nlm.nih.gov/Blast.cgi),下載甜蕎FaesAP2B同源蛋白序列。選取其中已公布的22種來自不同植物的AP2同源蛋白(表2),用MEGA 5.0軟件的ClustalW程序,對選取的序列進行同源比對,選Neighbor-Joining(鄰位相連法,NJ法),采用1000次的自展(Bootstrap)重復,構建分子系統進化樹(圖2),用Bioedit7.0軟件將甜蕎FaesAP2B轉錄因子與大麥HvAP2、辣椒CaAP2、芍藥PIAP2、蘋果MadoAP2這4個AP2同源蛋白序列進行序列比對與結構分析。
表2 構建分析系統發育樹的AP2同源蛋白
Table 2 AP2 homologous proteins for phylogentic tree construction

蛋白名稱Protein種名SpeciesGenbank登錄號Genbank accession numberLcAP2綠獨行菜Lepidium campestre L.AGI62074.1HvAP2大麥Hordeum vulgare Linn.AGX92954.1OitaAP2意大利紅門蘭Orchisi talica Poir.AGV39618.1CaAP2辣椒Capsicum annuum Linn.AJC11183.1PIAP2芍藥Paeonia lactiflora Pall.AGI61068.1MadoAP2蘋果Malus pumila Mill.ADE41133.1ViviAP2葡萄Vitis vinifera Linn.NP_001267881.1DcOAP2木石斛Dendrobium crumenatum Sw.AAZ95247.1CiarAP2鷹嘴豆Cicer arietinum Linn.XP_004511904.1ZmAP2玉米Zea mays L.ABR19870.1MtAP2蒺藜苜蓿Medicago truncatula Gaertn.KEH31465.1 FaesAP2B甜蕎Fagopyrum esculentum Moench.QCT84045.1OsAP2水稻Oryza sativa Japonica L.CAJ86049.1KjAP2棣棠Kerria japonica(Linn.) DC.AXR86367.1CacaAP2木豆Cajanus cajan(Linn.) Millsp.XP_020237263.1AtAP2擬南芥Arabidopsis thaliana(L.) Heynh.NP_001190938.1PtAP2L1黑松Pinus thunbergii ParlatoreBAD16603.1LmAP2L1落葉松Larix gmelinii(Ruprecht) KuzenevaABM26974.1NiatAP2煙草Nicotiana tabacum Linn.OIT36841.1AdAP2蔓花生Arachis duranensisXP_015963905.1CrAP2蘇鐵Cycas revoluta Thunb.BAE48512.1PsAP2牡丹Paeonia suffruticosa Andr.ALI57167.1
1.2.4 甜蕎FaesAP2B基因的表達分析
用HiScript?Ⅱ QRT SuperMix for qPCR(+g DNA wiper)(Vazyme)試劑盒,除去上述7種組織總RNA中殘存的基因組DNA再反轉錄合成第一鏈cDNA。具體操作步驟嚴格按照說明書進行。根據熒光定量引物設計的原則在甜蕎FaesAP2B基因的特異性位置設計上下游引物QFaesAP2BF和QFaesAP2BR(表1),以蕎麥Faesactin(GenBank登錄號:HQ398855.1)為內參基因,內參基因引物分別為QFaesactinF和QFaesactinR(表1)。逆轉錄分別合成上述7種組織的cDNA,濃度稀釋10倍后作為模板,使用ChamQ? SYBR? qPCR Master Mix(Vazyme)試劑盒,進行qPCR擴增,檢測甜蕎FaesAP2B基因在甜蕎lpls突變體根、莖、幼葉、花被片、雄蕊、雌蕊以及發育4 d的果實中表達的組織特異性和相對表達量的變化。3個生物學重復,擴增體系及PCR程序參照說明書。基因的相對表達量利用2-ΔΔCT方法計算[9]。采用SPSS17.0軟件對結果進行統計分析,采用最小顯著性差異法(Least significant difference,LSD)對甜蕎FaesAP2B基因的表達水平進行顯著性差異分析,并用Microsoft Office Excel軟件作圖。
2 結果與分析
2.1 甜蕎FaesAP2B基因全長cDNA克隆
同源克隆的方法結合RACE技術,從甜蕎lpls突變體花芽中分離出一個AP2同源基因。其cDNA序列全長1 788 bp,其中5′UTR長51 bp,3′UTR長357 bp,其包含1個長1 380 bp完整的ORF,編碼1個由459個氨基酸殘基構成的AP2/ERF轉錄因子,命名為FaesAP2B(FagopyrumesculentumAP2B),GenBank登錄號為MK290847.1。
2.2 蛋白序列比對與分子系統發育分析
系統發育分析如圖2所示,結果表明甜蕎FaesAP2B與其他被子植物AP2蛋白聚為一類,說明其屬于AP2同源蛋白。且甜蕎FaesAP2B與其他雙子葉植物的AP2蛋白距離較近,表明其于雙子葉植物AP2親緣關系較近,而與禾本科、蘭科等單子葉植物以及松科等裸子植物分隔較遠,親緣關系相對較遠。與經典分類學研究結果基本一致。
蛋白序列比對結果如圖3所示,該蛋白含有2個高度保守的AP2結構域,第1個AP2結構域位于第132~197號氨基酸之間,由66個氨基酸殘基組成;第2個AP2結構域位于第223~279號氨基酸之間,由57個氨基酸殘基組成。第1個AP2結構域前還存在1個由10個氨基酸殘基組成的高度保守的核定位信號區,位于第121~130號氨基酸殘基之間。該蛋白的氨基酸序列與其他4種植物的AP2同源蛋白的氨基酸序列除在2個AP2結構域和核定位信號區基本相同外,其編碼的氨基酸與另外4種植物的AP2同源蛋白的氨基酸序列存在較大差異,這表明AP2同源基因的功能在不同植物中存在一定差異。
2.3 甜蕎FaesAP2B基因的表達分析
利用實時熒光定量PCR檢測甜蕎FaesAP2B基因在甜蕎長雌蕊長雄蕊突變體“lpls”的根、莖、葉、被片、雄蕊、雌蕊以及發育4 d的果實中的表達量。檢測結果顯示如圖4,FaesAP2B基因在甜蕎突變體lpls的7種不同組織中均有表達,且甜蕎FaesAP2B在花被片、雌蕊、雌蕊和發育4 d的果實中的表達量均極顯著高于其在根、莖和葉等營養器官中的表達量(LSD,P<0.01)。其中,甜蕎FaesAP2B在雄蕊中的表達量最高,且極顯著高于在其他6個器官中的表達量(LSD,P<0.01),但該基因在其根、莖、葉間的表達量無顯著性差異。
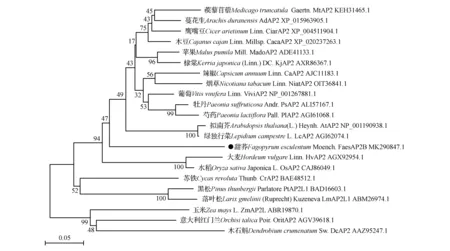
圖2 FaesAP2B蛋白與其他植物AP2同源蛋白分子系統進化樹 分支上的數字表示執行1 000次重復計算獲得的自展百分比。Fig.2 Phylogenetic analysis of FaesAP2B with other AP2 like proteins Number represents the Bootstrap percentage values calculated by 1 000 replicates.
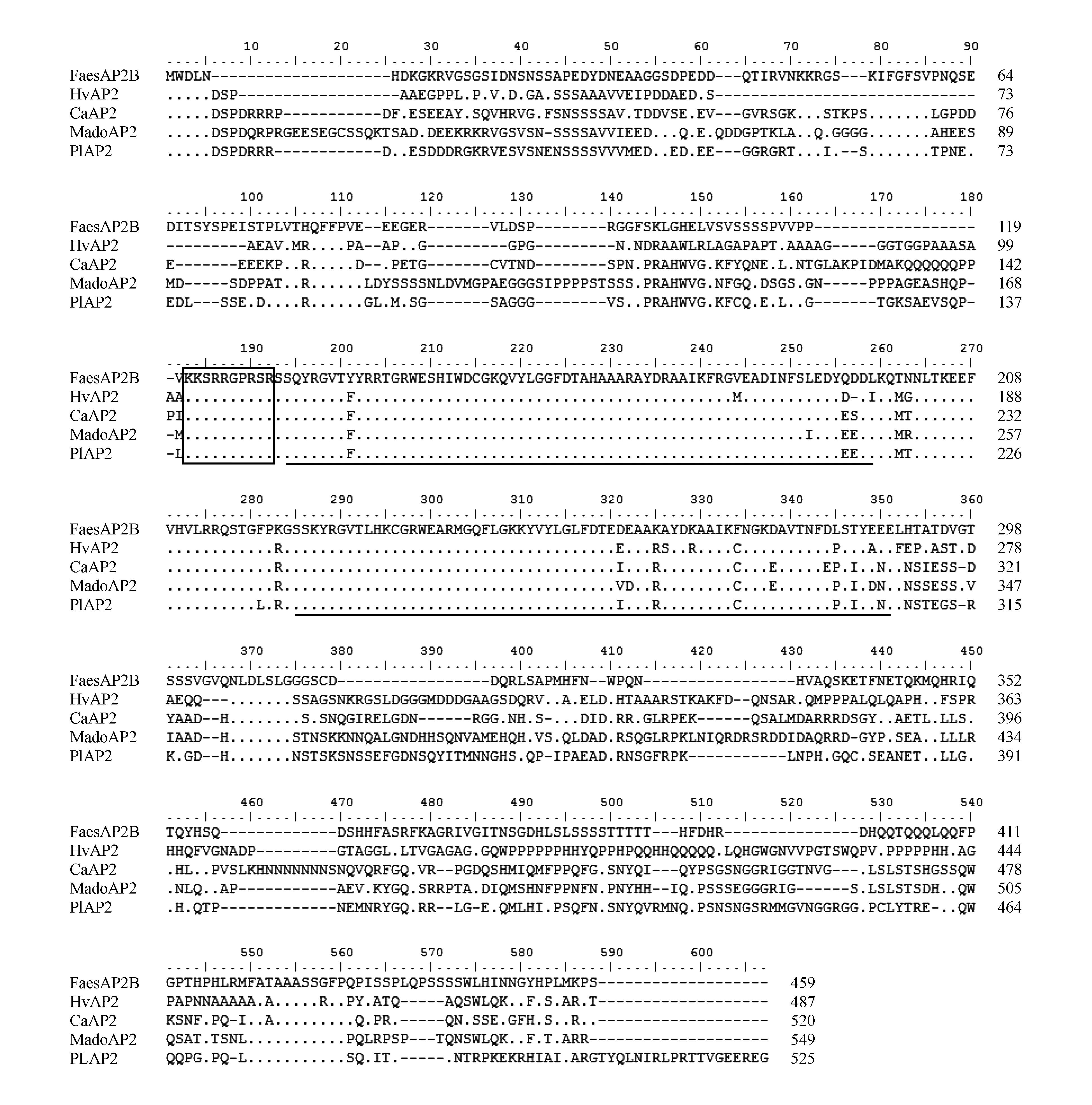
圖3 FaesAP2B蛋白序列比對與結構分析 方框為核定位信號區,下劃線表示AP2結構域。其中FaesAP2B、HvAP2、CaAP2、MadeAP2、PIAP2分別為甜蕎、大麥、辣椒、蘋果、芍藥的AP2同源蛋白;點表示相同的氨基酸。Fig.3 Alignment of the putative amino acid sequence of FaesAP2B with other homologous proteins sequences Putative nuclear localization signals of FaesAP2 in front of the first AP2 domain are boxed; Underlined regions represent AP2domains. FaesAP2B,HvAP2,CaAP2,PIAP2,MadoAP2 of AP2 homologous proteins from Fagopyrum esculentum Moench.,Hordeum vulgare Linn.,Capsicum annuum Linn.,Malus pumila Mill.,Paeonia lactiflora Pall.,respectively; Dots represent the same amino acid.
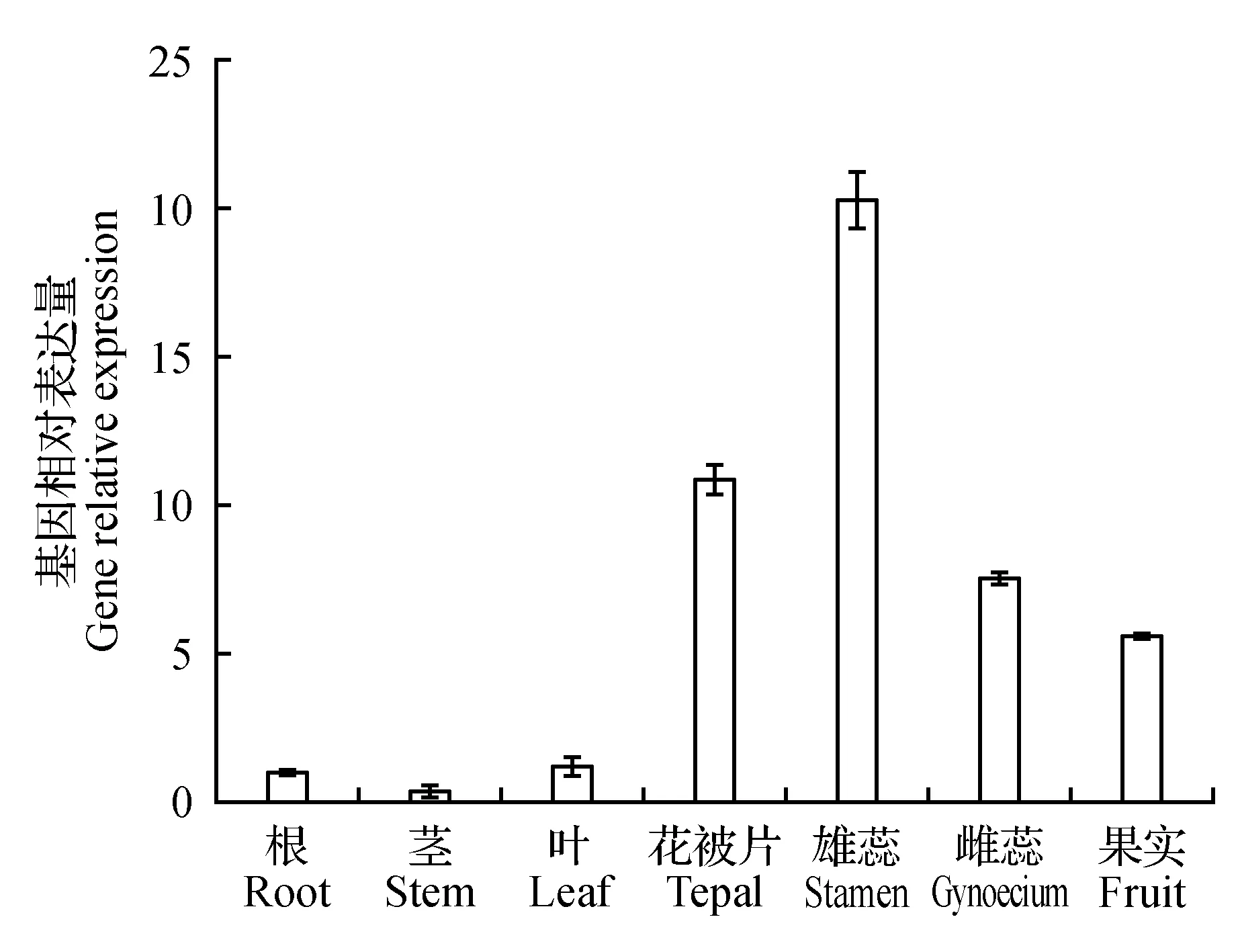
圖4 FaesAP2B基因在甜蕎不同組織器官中的表達量Fig.4 Expression of FaesAP2B in different organs of F.esculentum
3 討論
在模式物種擬南芥中,AP2基因是典型的A功能基因,是唯一一類不屬于MADs-box基因家族的花發育基因[10]。其控制第1、2輪(花萼、花瓣)花器官的發育,并抑制C類基因在外2輪花器官中表達[11~12]。擬南芥AP2基因具有編碼AP2/ERF轉錄因子家族的特有的結構域,AP2/ERF超基因家族存在于所有植物中,其中,ERF亞類氨基酸序列中只包含1個高度保守的AP2結構域,而AP2亞類含有2個串聯的AP2結構域,每個功能結構域有60~70個氨基酸組成,且高度保守[13~14]。AP2/ERF超級基因家族在植物花器官發育、花序分生組織發育的調控以及胚、胚乳、果實種子的發育,種子大小控制等方面發揮重要作用,與作物種子器官的產量與質量有密切關系[15~16]。同時,還參與植物生長激素的響應調節以及在逆境脅迫的信號轉導中發揮作用[17~19]。
作為A類基因,AP2在植物花分生組織形成、花器官發育的調控網絡中發揮重要作用。在強ap2突變體擬南芥中,其花瓣缺失,第1輪和第2輪花器官數量減少,并且第1輪花萼同源轉變成心皮狀結構,而在弱ap2突變體擬南芥中,其花萼葉化,花瓣雄蕊化[20~21]。這些現象說明AP2基因在控制擬南芥的花被(花萼和花瓣)發育起著重要的作用。此外也有研究發現AP2不僅僅在花器官中表達,在莖、葉中也有表達[22~23]。在辣椒中,AP2同源基因CaAP2在其葉、根莖、果皮中均檢測到了該基因的表達,且主要在營養分生組織階段發揮作用,而在成花過渡階段表達量下降[24]。同樣,AP2同源基因在美味獼猴桃(Actinidiadeliciosa(A.Chev.) C.F.Liang et A.R.Ferguson)和睡蓮(NymphaeatetragonaGeorgi)的營養器官及生殖器官也都有表達,這與甜蕎FaesAP2B基因的表達模式類似。且過表達的睡蓮AP2基因NsAP2還可以影響GA的生物合成途徑從而改變節間長度。另外,NsAP2異位表達也可使擬南芥花瓣數量增加[25~26];在芍藥中,AP2的同源基因PIAP2主要調控心皮和萼片的發育,而在花瓣和雄蕊中維持低水平表達[27];矮牽牛(Petuniahybrida(J.D.Hooker) Vilmorin)的AP2同源基因PHAP2A過表達可以恢復擬南芥ap2-1突變體表型,但當該基因被敲除后發現并沒有相應的突變體表型,這說明其可能存在功能冗余[28];而在玉米中,檢測到AP2-like基因在18種組織中均有表達[29];在落葉喬木雜交落葉松中,AP2的同源基因LmAP2L1可調控體胚發生后期胚胎的萌發[30]。甘薯(Dioscoreaesculenta(Lour.) Burkill)中,AP2同源基因則作為轉錄因子在甘薯非生物脅迫抵抗能力上發揮重要作用[31];而在大麥中,HvAP2基因由mir172調控,參與穗分化以及節間長度的調控,并與大麥花序的大小以及形狀,且過量HvAP2 mRNA可抑制節間生長并增加穗密度[32]。以上研究結果都表明AP2同源基因在不同物種中,其表達模式以及功能不盡相同。但總的來說主要在花和果實的發育中發揮重要作用。
在本研究中,我們通過實時熒光定量PCR發現甜蕎FaesAP2B基因在花器官及果實中表達量較高,尤其是雄蕊中,另外在生殖器官外的組織中也檢測到了甜蕎FaesAP2B基因的表達,這些結果表明,甜蕎FaesAP2B基因可能參與調控甜蕎花及果實的發育,且主要在雄蕊的發育過程中發揮重要作用。另外,其作用不只局限于花發育,還可能參與了甜蕎整個生長發育過程。其具體功能仍需進一步研究。
