利川腌菜細菌菌群多樣性及其與風味相關性研究
2020-03-28 08:15:32向凡舒鄧長陽王彥杰侯強川
中國釀造 2020年1期
關鍵詞:分析
向凡舒,梅 洋,鄧長陽,王彥杰,郭 壯,2,侯強川
(1.湖北文理學院 食品科學技術學院,湖北 襄陽 441053;2.恩施市公共檢驗檢測中心,湖北 恩施 445000)
利川腌菜原材料主要是俗稱“瓢兒菜”或“疙瘩菜”的大青菜,經晾曬清洗后加入鹽、辣椒和大蒜等調味料后,裹扎放入壇中壓緊封壇,大約一個月后方可食用,并且隨著儲存時間延長,味道越濃郁美味。目前我國蔬菜腌制品的發酵大多借助天然附著在蔬菜表面微生物的作用進行[1],腌菜生產過程中,微生物活動不僅影響腌菜發酵時間,對腌菜風味物質及營養成分形成也有重要作用[2]。準確揭示不同腌菜的菌群組成及其與腌菜風味之間的關系,對于腌菜的規模化生產具有重要意義。分子生物學方法的興起擴大了人們對發酵蔬菜中微生物的認識,PLENGVIDHYA V等[3]在工業蔬菜發酵液中利用脫氧核糖核酸(deoxyribonucleic acid,DNA)指紋圖譜鑒定到686株乳酸菌。尚雪嬌等[4]以恩施地區腌菜為研究對象,使用變性梯度凝膠電泳結合高通量測序的方法探究了樣品中乳酸菌和細菌的多樣性。顏娜等[5]采用高通量測序技術解析了恩施地區酸蘿卜中的細菌組成。二代高通量測序技術以無需培養和檢測速度快的優點廣泛應用于發酵食品微生物多樣性的研究中,其中16S rRNA是鑒定樣品中細菌微生物組成的主要方法,大量細菌微生物多樣性的研究以樣品宏基因組DNA 16S rRNA單可變區或多可變區進行擴增和測序[6-7],以全面解析樣品中菌群的組成。此外,腌菜的風味是影響腌菜質量的重要因素之一[8],微生物通過代謝活動,可以產生糖、酸、醇、酯、氨基酸等產物[9],賦予腌菜各具特色的風味。而通過電子鼻技術可有效揭示樣品中不同揮發性物質的濃度,目前該技術已廣泛用于不同發酵食品中揮發性物質的檢測中[10-12]。
本研究采用Illumina MiSeq高通量測序技術和電子鼻技術相結合的方法,全面解析了利川腌菜中細菌菌群結構和風味組成,同時分析了兩者之間的相關關系,以期為全面掌握發酵蔬菜中細菌和風味組成及改進發酵蔬菜的制作工藝提供有益參考。
1 材料與方法
1.1 材料與試劑
1.1.1 樣品采集
從湖北省恩施土家苗族自治州利川市采集十份腌菜樣品,其中樣品LCYC1~LCYC3采集自南門菜市場、LCYC4~LCYC6采集自解放東路菜市場、LCYC7~LCYC10采集自大北門農貿市場。所有樣品外觀無明顯差異,制作原料為利川市本地產大青菜,制作地點在利川市本地,所有樣品無明顯異味和霉斑且制作時間均在1個月左右。
1.1.2 試劑
qiagen69514食品基因組DNA提取試劑盒:德國QIAGEN公司;脫氧核糖核苷三磷酸(deoxy-ribonucleoside triphosphate,dNTP)Mix、聚合酶鏈式反應(polymerase chain reaction,PCR)緩沖液(10×)、DNA聚合酶(5 U/μL):北京全式金生物技術有限公司;16S rRNA V1~V3區引物由武漢天一輝遠生物科技有限公司合成。
1.2 儀器與設備
CT15RE臺式冷凍離心機:日本HITACHI公司;2100芯片生物分析儀:美國Agilent公司;ND-2000C微量紫外分光光度計:美國Nano Drop公司;vetiri梯度基因擴增儀:美國AB公司;FluorChem FC3化學發光凝膠成像系統:美國Fluor Chem公司;MiseqPE300高通量測序平臺:美國Illumina公司;R920機架式服務器:美國DELL公司。
1.3 方法
1.3.1 樣品DNA提取和16S rRNA V1~V3區序列擴增
按照qiagen69514食品基因組DNA提取試劑盒說明書步驟進行10份利川腌菜樣品的宏基因組DNA提取,使用V1~V3區域引物27F(5'-AGAGTTTGATCCTGGCTCAG-3)和338R(5'-TGCTGCCTCCCGTAGGAGT-3')對樣品V1~V3區進行擴增。PCR擴增體系(20μL):10×PCR緩沖液(含Mg2+)4 μL,dNTPs Mix(2.5 mmol/L)2 μL,正反向引物(5 μmol/L)0.8 μL,DNA聚合酶0.4 μL,DNA模板10 ng,雙蒸水(ddH2O)補充至20 μL。PCR擴增條件:95 ℃預變性3 min;95 ℃變性30 s,55 ℃退火30 s,72 ℃延伸45 s,35 個循環;72 ℃末端延伸10 min。
1.3.2 基于電子鼻對樣品揮發性風味物質測定
稱取10 g腌菜于樣品瓶中,60 ℃水浴保溫20 min后室溫平衡10 min,采用頂空吸氣法,進樣吸氣流量200 mL/min,傳感器清潔95 s(如果發現曲線不平或離基線遠可再清洗一遍或直接增加清洗時間),樣品準備時間5 s,測定時間60 s,選取49 s、50 s和51 s時的響應值求取平均值進行分析,樣品間間隔1.5 min。該電子鼻中包含10個金屬氧化物傳感器陣列,用于分析物質中不同揮發性成分,傳感器陣列及性能描述見表1。

表1 傳感器陣列及性能描述Table 1 Sensor array and performance description
1.3.3 細菌多樣性分析
對測序后的序列進行質控(目的片段長度>300 bp,引物和標簽序列完全匹配,整條序列質量>20 的堿基所占比例須>93%),使用QIIME數據分析(V1.7)平臺參照CAPORASO J G等[13]的方法對高質量序列進行后續分析,以97%相似度劃分操作分類單元(operational taxonomic units,OTUs),選取每一OTU的代表性序列使用Greengenes(V13.8)和核糖體數據庫(ribosomal database project,RDP)(V11.5)數據庫進行同源性比對,統計樣品中細菌在各分類學水平上的菌群組成。
由表2可知,在1h時12T-103菌株對Cry1Ac蛋白降解降較為明顯,由15. 95 μg·L-1降低至0. 68 μg·L-1,降低程度達極顯著水平(p<0. 01),其降解率達92. 26%,此后12T-103菌株對Cry1Ac蛋白降解較為緩慢且不顯著。由此,12T-103菌株對Cry1Ac蛋白的降解能力較強。
1.3.4 數據處理
使用Origin(V8.6)軟件和R語言軟件(v3.3.2)對數據進行進一步的分析和可視化。使用非參數的Kruskal-Wallis和Mann-Whitney秩和檢驗分別對多組和兩組樣品進行差異顯著性檢驗。
2 結果與分析
2.1 利川腌菜樣品中細菌豐富度及多樣性分析
對10份腌菜樣品細菌16S rRNA基因序列V1~V3區進行Miseq高通量測序,通過α多樣性曲線評估各樣品測序量是否足夠及細菌微生物多樣性,結果如圖1所示。
由圖1可知,10份腌菜的稀疏曲線均未達到平臺期,表明隨著測序量增大仍然會發現新物種,但香農曲線已經到達平臺期,說明隨著測序量增加細菌多樣性基本不發生變化,當前測序量已足以揭示樣品中主要菌群的組成。
利川腌菜的測序序列信息和α多樣性指數如表2所示。由表2可知,共得到341 560條高質量序列,平均每份樣品34 156條序列(15 927~52 610條),對這些代表性序列在97%相似度水平上劃分OTU,共獲得14 228個典型OTU,平均每份樣品3 843個OTU(1 001~6 671)。
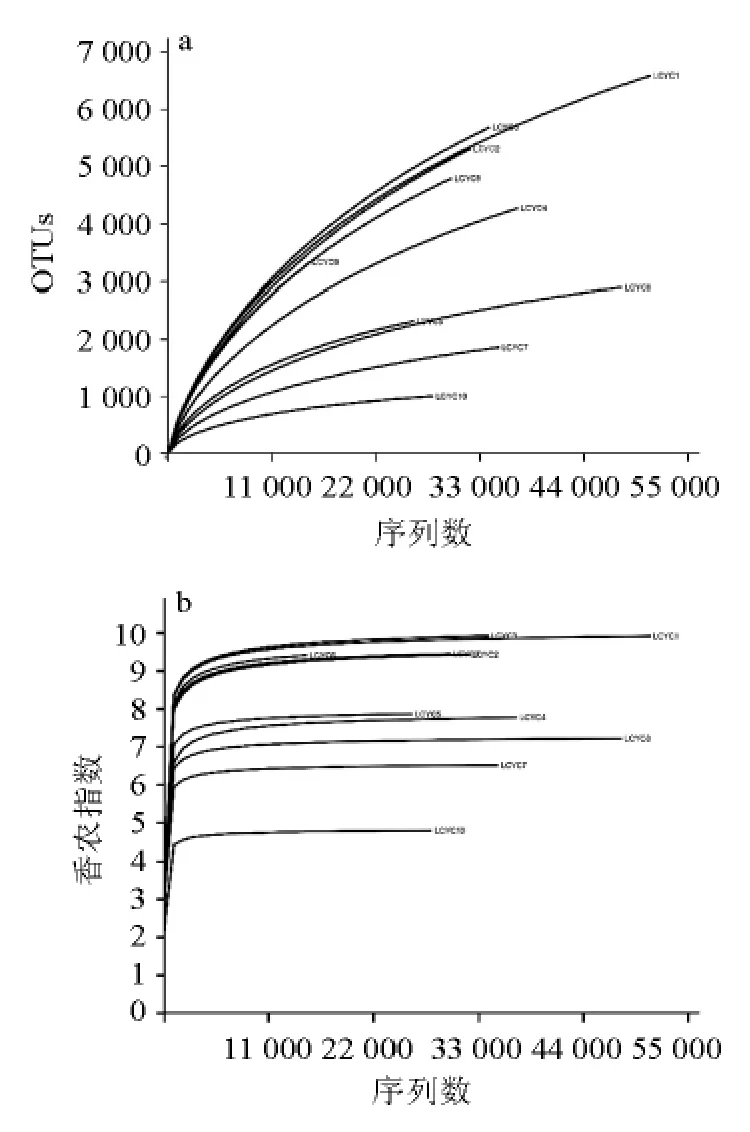
圖1 各樣品細菌組成的稀疏曲線(a)和香農曲線(b)Fig.1 Rarefaction curve (a) and Shannon curve (b) of bacteria compositions in each sample

表2 利川腌菜樣品中細菌群落多樣性和豐富度分析Table 2 Analysis of bacterial community diversity and abundance of Lichuan pickles samples
2.2 利川腌菜中細菌種群結構分析
分別在門和屬的水平對利川腌菜中的細菌組成進行了注釋和分析。在門水平,所有樣品共注釋到13個細菌門,平均每個樣品可檢測到9.0±2.5個細菌門,結果如圖2a所示。由圖2a可知,其中平均相對含量>1%的菌門有4個,分別為厚壁菌門(Firmicutes)(44.4%)、變形菌門(Proteobacteria)(40.4%)、放線菌門(Actinobacteria)(13.5%)和擬桿菌門(Bacteroidetes)(1.5%),雖然各樣品門水平組成不盡相同,但各樣品優勢菌門一致。
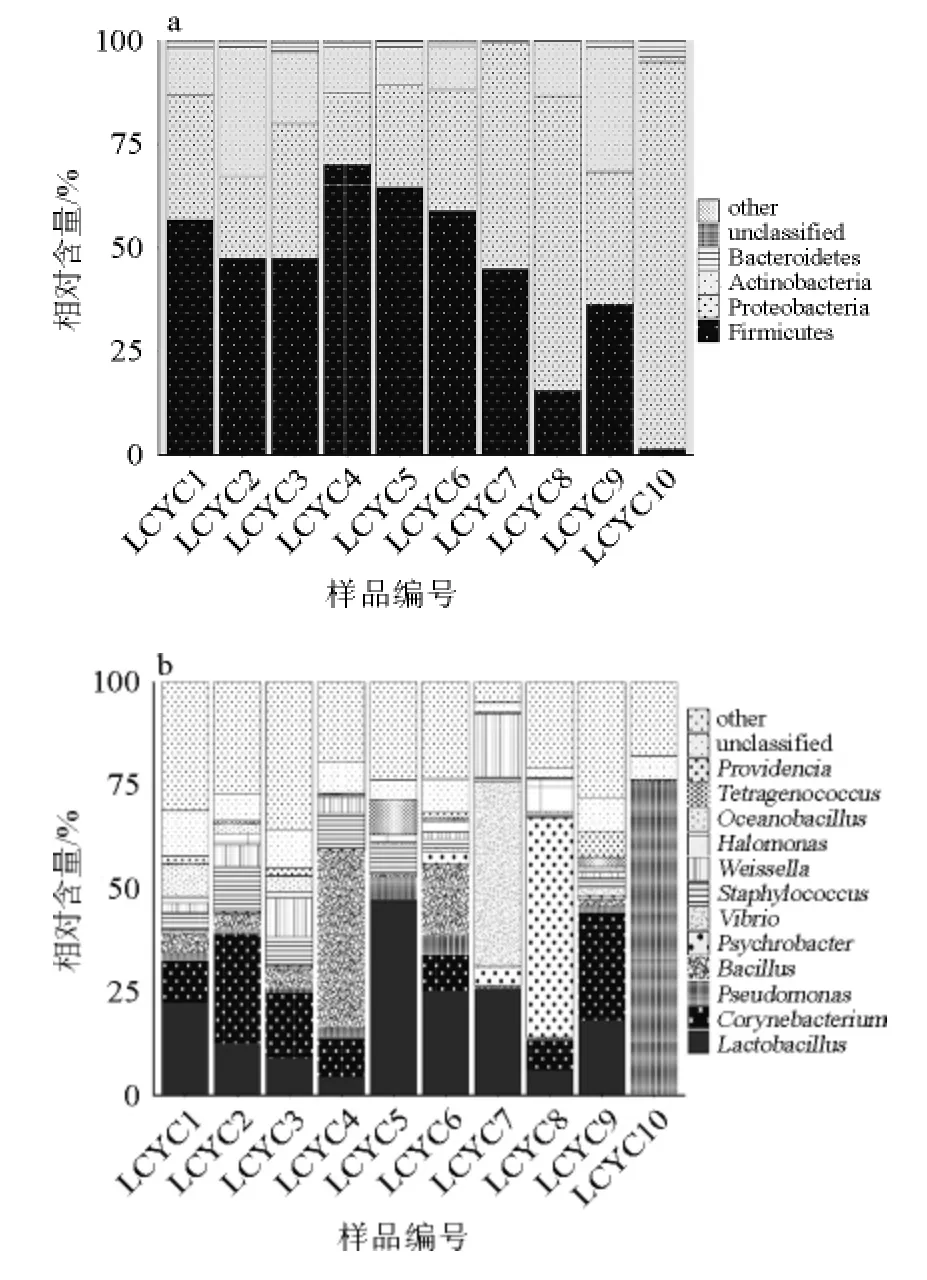
圖2 基于門水平(a)和屬水平(b)利川腌菜樣品中菌群組成Fig.2 Compositions of microbiota at phylum (a) and genus (b) level in different Lichuan pickles samples
由圖2b可知,在屬水平,所有樣品共注釋到386個屬,平均每個樣品可檢測到(210±59)個細菌屬。其中12個菌屬的平均相對含量>1%,主要包括乳桿菌屬(Lactobacillus,17.1%),棒狀桿菌屬(Corynebacterium,10.5%),假單胞菌屬(Pseudomonas,9.5%),芽孢桿菌屬(Bacillus,7.6%),嗜冷桿菌屬(Psychrobacter,6.4%),弧菌屬(Vibrio,4.9%),葡萄球菌屬(Staphylococcus,4.3%),魏斯氏菌屬(Weissella,4.1%),鹽單胞菌(Halomonas,1.9%),大洋芽孢桿菌屬(Oceanobacillus,1.6%),四聯球菌屬(Tetragenococcus,1.2%)和普羅維登斯菌屬(Providencia,1.2%)。不同樣品的優勢屬亦存在一定的差異,如樣品LCYC5、LCYC6和LCYC7中乳桿菌屬的相對含量較高,樣品LCYC4和樣品LCYC6中芽孢桿菌屬的相對含量較高,而樣品LCYC7、LCYC8和LCYC10分別擁有較高數量的弧菌屬、假單胞菌屬和嗜冷桿菌屬。利川腌菜樣品菌群在屬水平存在著較大差異,但導致菌群差異的具體原因尚不是十分清楚,推測可能與利川腌菜多由自家生產,非工業化統一生產有關。
2.3 利川腌菜樣品中細菌菌群差異性分析
對測序獲得的有效數據進行Beta多樣性分析,分別基于加權和非加權Unifrac距離對10份腌菜樣品進行了主坐標分析,以直觀的方式展現不同樣品間菌群的差異和相似性,結果見圖3。
2.4 利川腌菜中揮發性風味物質分析
電子鼻技術是基于氣體傳感陣列響應圖譜來識別氣味的電子系統,廣泛應用于風味物質的分析檢測中[15-16]。本實驗使用該技術對10份利川樣品風味物質進行分析,各樣品風味物質檢測結果見表3。
由表3可知,利川腌菜樣品間不同風味的響應值存在較大差異,整體而言,利川腌菜中W5S、W1S、W1W和W2S的響應值最高,平均響應值分別為44.83、79.90、52.04和43.25,表明腌菜風味物質中氮氧化合物、甲烷、萜類化合物和乙醇較豐富。萜類化合物在酸和酶水解作用下易形成游離態揮發性物質,從而賦予產品香氣。此外,該物質香氣閾值低,在低濃度下可對產品香氣產生較大影響[17]。醇類物質主要來源于微生物酒精發酵,使產品具有輕快醇香味[18]。相反,W1C、W3C和W5C的響應值較小,平均響應值僅為0.10、0.17和0.18,表明利川腌菜中芳香類物質含量較低。
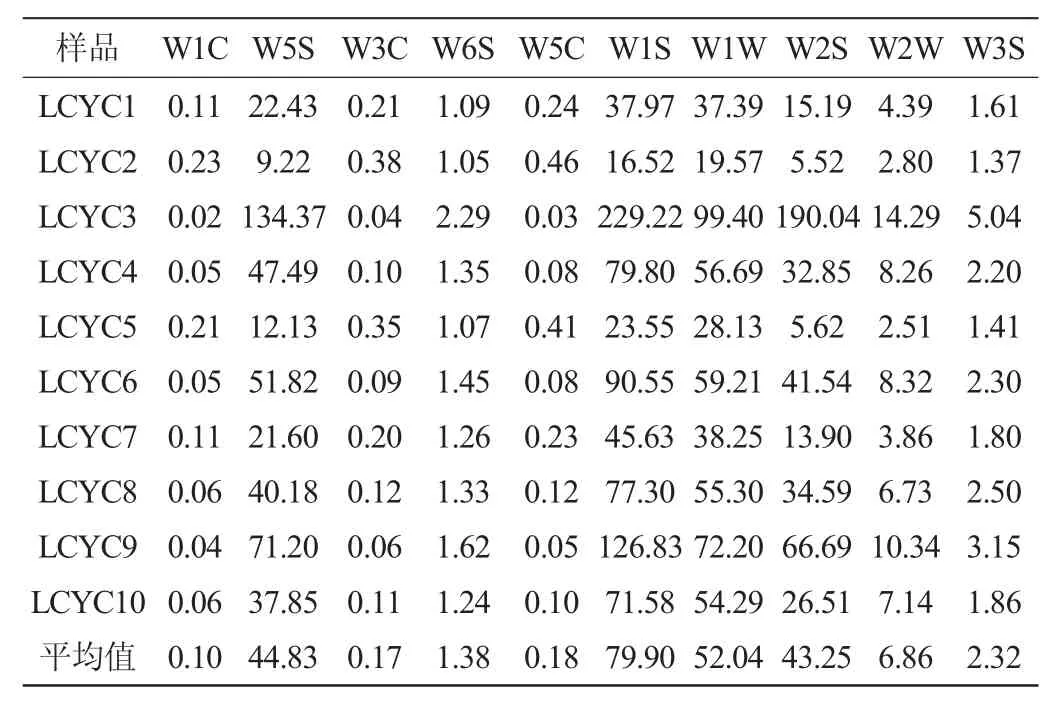
表3 利川腌菜樣品中揮發性風味物質電子鼻傳感器響應值Table 3 Flavor response value of volatile flavor compounds in Lichuan pickles samples
2.5 利川腌菜中揮發性風味物質差異分析
主成分分析展現了不同樣品間的風味差異,結果見圖4。由圖4可知,10份樣品大致分為3簇,其中樣品LCYC3單獨為一簇(組1),樣品LCYC2和LCYC5(組2)為一簇,其余樣品為一簇(組3)。對上述3組樣品的風味響應值統計分析,結果見圖5。
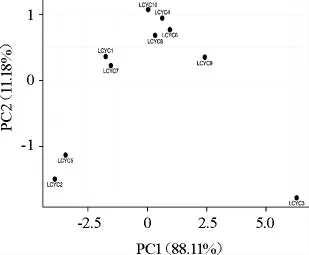
圖4 基于主成分分析利川腌菜樣品中風味響應值差異分析Fig.4 Difference analysis of flavor response value of Lichuan pickles samples based on principal component analysis

圖5 基于主坐標聚類的利川腌菜樣品風味響應值差異分析Fig.5 Difference analysis of flavor response value of three groups of Lichuan pickles samples based on principal coordinate clustering
由圖5可知,三組樣品存在較大差異的風味是W5S、W1S、W1W和W2S,分別代表氮氧化合物、甲烷、萜類化合物和乙醇,上述風味在組1中的響應值分別為134.37、229.22、99.40和190.04;組2中的響應值分別為10.67、20.04、23.85和5.57;組3中的響應值分別為41.79、75.67、53.33和33.04。說明這四種風味物質對利川腌菜整體風味的貢獻較大。
2.6 菌群結構與揮發性物質相關性分析
由菌群結構分析和風味物質分析發現,不同樣品菌群結構和風味物質之間都存在較大的差異。因此,本研究將相對含量>0.5%的菌屬與風味響應值之間進行了Spearman相關性分析,結果見圖6。由圖6可知,部分菌群與風味響應值之間存在顯著相關性:其中變形桿菌與W3S、W2W、W6S、W2S、W5S、W1S和W1W呈顯著正相關(P<0.05),與W3C、W5C、W1C等芳香類風味響應值呈顯著負相關(P<0.05);嗜冷桿菌屬與W3S和W6S呈顯著正相關(P<0.05);而腸球菌屬與W2W、W2S和W5S呈顯著正相關(P<0.05),眾多研究者認為腸球菌廣泛存在于發酵食品中,不僅影響食品的風味、香氣和質地,還可產生乙醇、過氧化氫等物質[19-20],這與本研究結果一致。此外,風味響應值W5S、W1S、W1W和W2S與樣品中70.4%相對含量>0.5%的菌屬如腸球菌屬、弧菌屬、芽孢桿菌屬等存在正相關關系,與之相反,風味響應值W1C、W3C和W5C則與上述菌群存在負相關關系,盡管上述相關性均不顯著。可能在這些菌屬的共同作用下,最終發酵蔬菜產品中上述響應值對應的氮氧化合物、甲烷、萜類化合物和醇類物質的含量較高,而芳香類物質含量較低。相關性分析結果表明發酵利川腌菜中風味物質受多種微生物共同作用的影響,而非單一微生物發酵產生。

圖6 主要細菌屬與風味響應值相關性分析Fig.6 Correlation analysis between major bacterial genera and flavor response value
3 結論
本研究以利川地區腌菜為研究對象,利用Illumina MiSeq高通量測序技術和電子鼻技術全面解析腌菜中細菌多樣性及其風味物質,發現其細菌門主要為厚壁菌門(Firmicutes)、變形菌門(Proteobacteria)和放線菌門(Actinobacteria),主要細菌屬為乳桿菌屬(Lactobacillus)、棒狀桿菌屬(Corynebacterium)和假單胞菌屬(Pseudomonas)。利川腌菜中氮氧化合物、甲烷、萜類化合物和乙醇對利川腌菜整體風味的貢獻較大。不同樣品細菌組成和風味物質存在差異,腌菜中菌群的組成與腌菜風味之間存在密切聯系,提示在腌菜的工業化生產過程中需要密切關注發酵罐中腌菜的菌群的組成。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
