土壤微生物碳素利用效率研究進展
2020-03-27 09:24:52于貴瑞
生態學報 2020年3期
陳 智, 于貴瑞
1 中國科學院地理科學與資源研究所生態系統網絡觀測與模擬重點實驗室,北京 100101 2 中國科學院大學資源與環境學院,北京 101408
通過有效增加碳(C)固持和合理減少C排放來共同控制和減緩全球變暖是全人類應對氣候變化達成的共識[1]。為制定有效的增匯措施,人們迫切需要準確模擬和預測全球變暖與地球生態系統間的相互影響,尤其是陸地生態系統對全球變暖的反饋作用與機制[2- 3]。已有大量研究指出,全球變暖會促進土壤C釋放,對全球變暖產生正反饋[2,4]。但是,越來越多的研究開始發現,微生物在調控陸地生態系統對全球變化的反饋中起著重要作用,并且可能改變預期的反饋效應[5- 7]。例如,長期的增溫反而抑制微生物生物量和酶活性,從而減少對土壤有機碳的分解。微生物的生理代謝過程及其對外界環境變化的響應與適應成為影響陸地生態系統的反饋作用的關鍵[5]。
土壤微生物連接著土壤圈、生物圈、大氣圈、水圈、巖石圈的物質循環,參與了土壤中幾乎所有的物質轉化過程[8]。土壤微生物的生長直接取決于微生物的碳素利用效率(Microbial carbon use efficiency, CUE),即微生物將吸收C轉化為生物量C的效率。在大量的土壤C循環模型中,微生物CUE被設定為一個常數[9- 11]。然而,野外觀測和室內培養實驗并不支持這一假設。土壤微生物CUE可能隨著外界環境和養分條件等的變化而顯著變化。有研究指出,土壤微生物CUE隨著土壤養分有效性的增加而增加[12- 13],而隨著溫度的升高而降低[5,14]。但是,對于這些潛在因素的影響并沒有一致性的認識。例如,在北美大草原的研究表明,水分脅迫抑制微生物的生長和CUE[15]。然而,Herron等[16]發現土壤含水量的降低對土壤微生物CUE并無顯著影響。這些不同的研究結論體現了人們對于土壤微生物CUE變異及其影響機制認識的不足,這制約了人們對陸地生態系統的反饋的準確模擬與預測[17]。
本研究對現有文獻進行梳理和分析,概述土壤微生物CUE的定義和測定方法,重點綜述和分析CUE的變異特征及其影響因素,并指出未來的研究重點,以期為當前地球系統模型的改進及對未來的氣候變化趨勢預測提供理論依據。
1 土壤微生物碳素利用率的定義
植被通過光合作用將大氣中CO2轉化為有機物質,形成生態系統的凈初級生產力。絕大部分的植被生產力需要在分解者-土壤微生物的分解礦化作用下還原為無機養分,再被植被吸收利用,實現生態系統中物質和能量的生物地球化學循環。微生物的生理代謝過程實則是同化和異化代謝相耦合的過程。微生物將植被光合產物一部分同化為微生物生物量,另一部分以呼吸代謝形式釋放到大氣中。此過程中,微生物將植被生產力轉化為微生物生物量的效率稱為微生物的碳素利用效率[13,18],也稱為微生物生長效率或底物利用效率[19- 21]。土壤微生物CUE是土壤C循環過程中的關鍵生態學參數,它直接影響著生態系統的C素滯留時間、周轉速率以及土壤的C存儲能力[7,22- 23]。
在生態學研究中,微生物CUE通常表示為微生物的生長(μ)與吸收(U)的比值[13,18],即CUE=μ/U。微生物從外界吸收C主要用于微生物的生長(μ)、呼吸代謝(R)、胞外酶和代謝產物的分泌(EX)以及微生物死亡(BD)4個方面(圖1)。根據質量守恒原理,微生物U表示為:
U=μ+R+EX+BD
其中,土壤微生物呼吸(R)包括微生物用于生長(RG)、維持(RM)、胞外酶生產(RE)以及溢流過程(RO)中產生的呼吸[13],即:
R=RG+RM+RE+RO
根據CUE定義和質量守恒方程,微生物CUE表示為:
在自然生態系統中,EX和BD通常難以測定,并且相對于生長和呼吸量,EX和BD量很微小而常被認為可以忽略[13]。因此,CUE通常被認為是μ與R兩個過程的平衡關系,即,
這一定義被廣泛用于當前的微生物代謝和土壤碳循環模型中[13,18]。
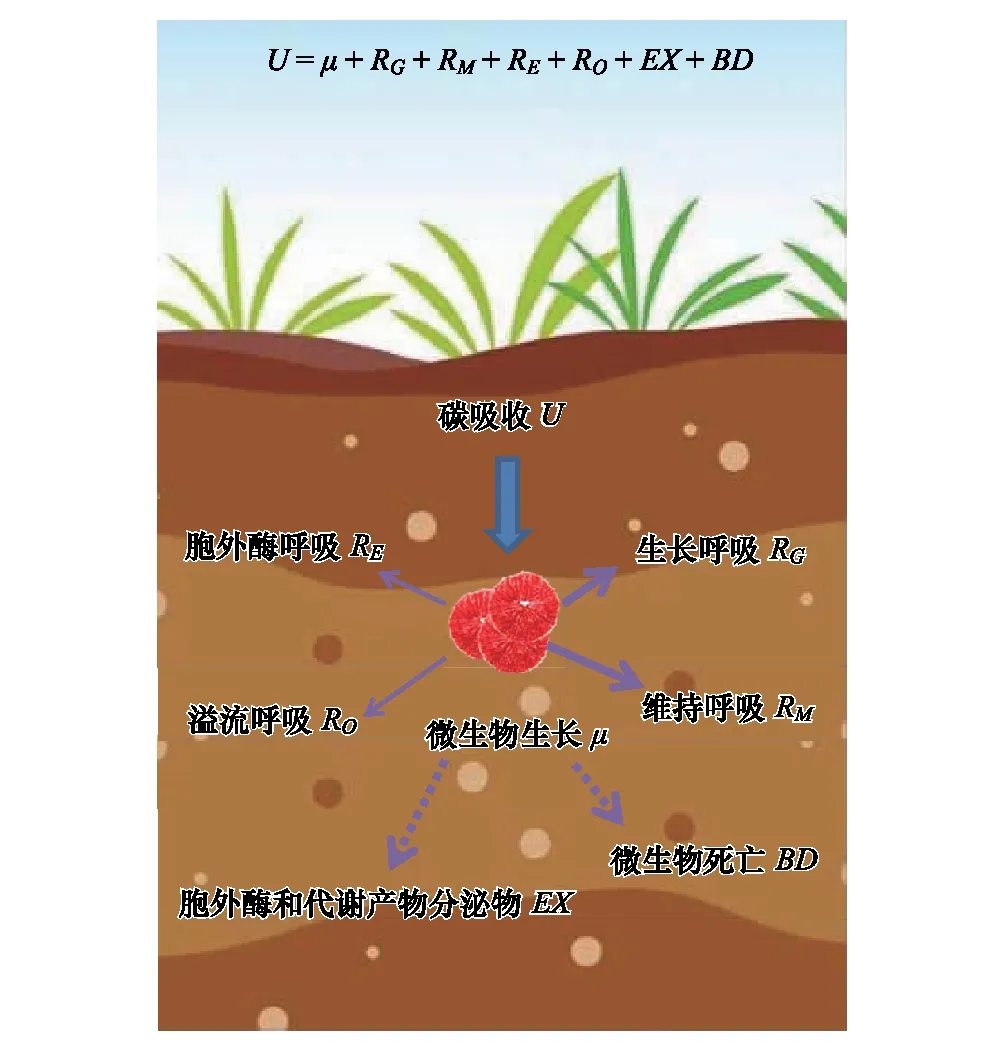
圖1 微生物生長代謝組分和質量平衡方程 Fig.1 Microbial metabolic components and equilibrium equation根據土壤微生物CUE的定義和Manzoni等[13]提出的土壤微生物代謝質量平衡方程繪制; U, 微生物碳吸收microbial carbon uptake; μ, 微生物生長microbial growth; RG, 微生物生長呼吸respiration for microbial growth; RM, 微生物維持呼吸respiration for microbial maintenance; RE, 胞外酶呼吸respiration for extracellular enzyme production; RO, 溢流呼吸overflow respiration; EX, 胞外酶和代謝產物分泌物excretion of extracellular enzymes and metabolites; BD, 微生物死亡microbial mortality
2 土壤微生物碳素利用率的測定方法
土壤微生物CUE多采用室內培養結合質量守恒法和標記物示蹤法進行測定。質量守恒法是直接測定物質質量或濃度的變化,根據物質守恒原理計算CUE。標記物示蹤法則是通過標記底物,有效追蹤底物的利用路徑,計算底物被用于生長和呼吸的比例進行測定,是目前較為廣泛采用的方法。根據研究途徑、研究對象(微生物或底物)的不同,現有分析方法可大致分為基于微生物生長速率測定法、基于微生物生物量測定法、基于底物吸收速率測定法和基于底物濃度變化測定法。這些方法各具優缺點和適用范圍(表1)。
2.1 生長速率法
生長速率法是最早的測定方法之一,它通過直接測定微生物的生物合成速率和呼吸速率計算微生物CUE。以3H-thymidine、3H-leucine等作為標記底物,通過短時間的培養,在三氯乙酸的消解作用下分離出合成的核酸或蛋白質,結合放射性同位素分析,測定微生物將3H-thymidine合成為DNA或將3H-leucine合成為蛋白質的速率[24-26]。該方法直接準確,主要用于水生生態系統。在陸地土壤中,采用以18O標記的重水H218O進行培養,測定微生物從水中吸收18O合成DNA的量,結合微生物DNA和生物量C的轉化系數計算土壤微生物CUE。該方法可以直接測定微生物的生物合成速率,但主要用于短時期培養[27-28]。
2.2 生物量法
在陸地生態系統,例如土壤中,通常基于微生物生物量的變化計算CUE。添加標記的活性有機碳作為底物進行室內培養,結合氯仿熏蒸法測定微生物生物量的變化。該方法簡單,可操作性強,是一種常用的標準方法。但是,由于在較短時期內難以捕捉微生物群落的生長及維持呼吸,該方法在短期內所測定的活性標記物轉化為生物量的速率并不完全等同于微生物的生長速率,而更多的是群落的吸收速率,因此存在高估CUE的現象[29- 31]。
2.3 底物吸收速率法
底物吸收速率法是從觀測底物變化的角度測定CUE。將3H-thymidine、3H-leucine等作為標記底物,進行短期室內培養測定其被吸收利用的速率。該方法相對于微生物生物量測定法,考慮了微生物產物的損失。但是,底物的吸收速率通常難以準確測定,只適用于短期的室內培養分析。
2.4 底物濃度變化法
與底物吸收速率法相似,底物濃度變化法也是從底物變化的角度測定CUE。該方法通過添加高濃度的底物(葡萄糖, 醋酸等),經過一段時間的室內培養,階段性的測定底物濃度的變化,結合同期的呼吸量計算CUE。該方法通常不需要對底物進行同位素標記,但需要提供高濃度的底物介質。由于該方法需要一定長度的培養時間,因此考慮了微生物產物的代謝和分泌過程。

表1 微生物碳素利用率測定和計算方法比較
CUE, 碳素利用率carbon use efficiency;μ, 微生物生長速率microbial growth rate;R, 微生物總呼吸速率microbial total respiration rate;U, 底物吸收速率substrate uptake rate;ΔCB, 微生物生物量變化量microbial biomass change;ΔCS, 底物濃度變化量substrate concentration change;RCUM, 累積呼吸速率cumulative respiration rate;AE, 元素(E)的吸收效率assimilation efficiency of element E;BC∶E, 微生物生物量的C∶E比the C∶E ratio of microbial biomass; TERC:E, 微生物最優生長所需C∶E比閾值threshold of C:E ratio required for microbial optimum growth
2.5 化學計量比法
基于化學計量比法是一種不需要直接測定CUE而根據元素化學計量限制理論計算CUE的方法,被廣泛用于土壤和凋落物分解模型中[18,32]。根據化學計量平衡理論,當外源養分中的必需元素(如N,P)與C素的比例低于微生物最優生長所需N/P:C元素比閾值時,微生物的生長將會受到該元素有效性的制約。CUE與養分的元素計量比存在一階函數關系,表示為CUE=AE×BC:E/TERC:E。其中,AE表示必需元素(E)的吸收效率,BC∶E表示微生物生物量的C∶E 比,TERC:E表示微生物最優生長所需C∶E比閾值[33- 35]。該方法具有理論基礎,不需要實驗測定,可參數化和模擬,但是是一種經驗關系,主要用于土壤碳循環過程模型的模擬中。
3 土壤微生物碳素利用率的變異性
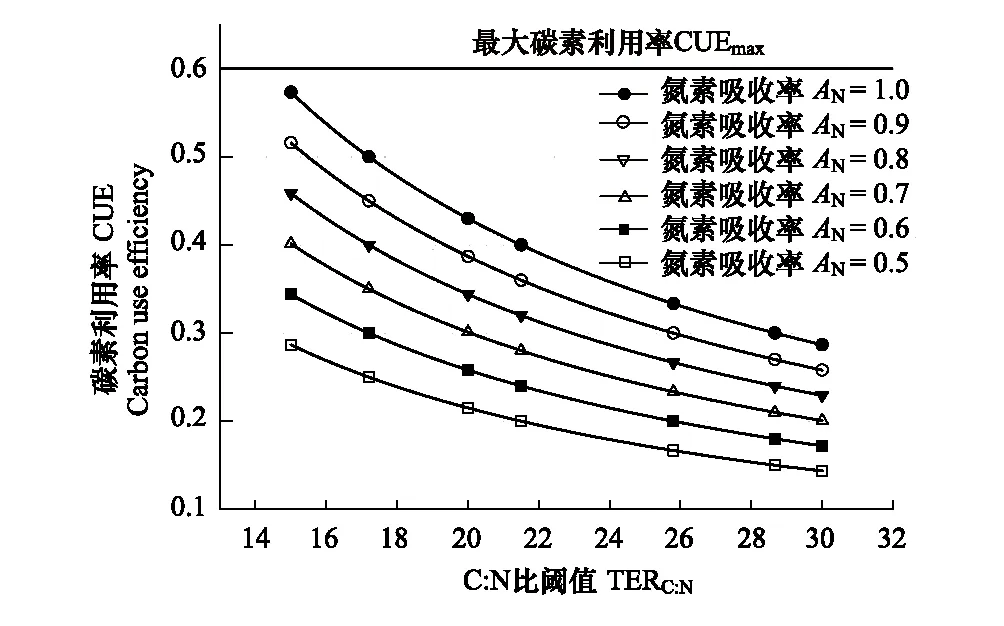
圖2 微生物碳素利用率CUE與C:N比閾值TERC:N和氮素吸收率AN的關系Fig.2 Relationships between microbial carbon use efficiency and threshold of C:N and nitrogen uptake efficiency根據生物化學元素計量平衡理論繪制,參見Sinsabaugh等[29]
在自然生態系統中,土壤微生物CUE并非常數。在理想狀態下,微生物只進行有機物同化并完全同化底物,那么碳素利用率為1。但是微生物的生長始終伴隨著呼吸代謝消耗,在熱力學限制下,微生物的最大CUE(CUEmax)小于1[38- 40]。微生物的實際生長受自身化學計量平衡的調控,表現為CUE隨著必需元素(E)的吸收效率、微生物的C∶E比和最優生長C∶E比閾值的變化而變化(圖2)。當氮素吸收效率AN=1,土壤微生物CUE在C:N比閾值TERC:N為15時可接近于最大CUE(CUEmax≈0.6);隨著TERC:N增加到30,土壤微生物CUE則降到CUEmax的1/2(CUEmax/2≈0.3)(圖2)。在實際土壤環境中,AN通常小于1,因此微生物實際CUE難以達到CUEmax。從ATP生成、電子傳遞和能量轉化3個不同途徑測定的結果表明,受到熱力學限制,微生物的實際最大CUE約為0.6[38- 39]。Sinsabaugh等[29]的研究指出,在考慮了微生物維持代謝消耗之后,熱力學最大CUE約為0.55。
基于實測數據的整合結果表明,土壤微生物CUE可在0.2—0.8的范圍內變化。Sinsabaugh等[41]通過建立的生態酶活性、生物量組成和養分濃度的化學計量模型,結合全球生態酶活性數據得出土壤微生物CUE平均約為0.29,接近于CUEmax/2。基于C:N元素化學計量平衡方程估算的結果也得出,土壤微生物CUE平均約為0.27±0.11,與基于C:P元素計量方程的估算結果相近(0.25±0.12)[18]。但是,實驗結果也顯示,不同土壤層的微生物CUE存在差異。例如,礦物層微生物CUE(0.284±0.005)高于有機層微生物CUE(0.205±0.008)[42]。不同微生物種群也表現出不同的CUE。Sinsabaugh等[43]基于實驗觀測數據的整合分析得出,細菌的CUE約為0.336±0.213,高于真菌的CUE(0.326±0.196)。土壤微生物CUE還因植被類型的不同而不同。Takriti等[44]對西伯利亞植被樣帶的調查研究表明,土壤微生物CUE從草甸草原向泰加林、苔原逐漸降低。
在當前大量的生物地球化學循環模型中,土壤微生物CUE則通常被設定成一個參數,集中在0.25—0.6的范圍(表2)。在不同的生物地球化學模型中,因考慮不同底物組成等影響,CUE參數值存在差異。例如CENTURY模型,地下和地上有機質的分解采用不同土壤微生物CUE,分別為0.45和0.55[9]。而Daisy、NCSOIL、ICBM等模型則考慮碳庫活性的影響,對于活性碳庫,CUE為0.6,而對惰性碳庫的CUE小于0.6[10- 11,45](表2)。但是,Manzoni等[32]指出,實測的土壤微生物CUE結果往往低于模型的預設值,當前的模型在一定上低估了真實的異氧呼吸通量。
4 土壤微生物碳素利用率的影響因素
土壤微生物CUE受到來自熱力學、生理學和生態學的調控,包括溫度、水分等環境因子、土壤質地、底物和養分有效性、微生物群落組成等多種因素的影響。
4.1 熱力學限制
微生物的碳吸收過程是同化和異化的耦合過程,微生物在轉化植被光合產物為自身生物量的同時,時刻伴隨著自身呼吸的代謝消耗。在熱力學限制下,微生物的理論最大CUE約為0.8[40]。這表明,不超過80%的吸收有機C可被微生物用于自身生長,而至少20%的C需要用于維持呼吸的能量消耗。然而,實際的土壤微生物CUE難以達到理論的最大值,因為在生理和生態環境因子的制約下,微生物還需投入更多的能量到呼吸和養分吸收等過程。
4.2 生態環境因子限制
環境因子,例如溫度、濕度、降水、土壤水分,會影響微生物代謝,改變μ和R間的平衡關系,從而影響CUE[5,13]。大量研究表明,土壤微生物CUE對增溫呈現負反饋,CUE隨著溫度的升高而降低[5,13- 14,53- 54]。這是因為在溫度的調控下,微生物的生長和代謝速率均隨著溫度的升高以Arrhenius方程的形式增加[55- 57]。然而,微生物呼吸代謝的溫度敏感性高于生長響應[55],微生物呼吸的增加速度快于微生物生長,使得CUE降低[5,54,58]。Steinweg等[59]的研究指出,溫度每升高1℃,土壤微生物CUE約減少0.009。高溫脅迫下微生物CUE的負反饋效應更為顯著[53]。模擬結果表明,30年的連續增溫降低了吸收C被用于微生物生長的比例,CUE從0.31減少到0.23[5]。但是,也有研究指出,溫度的升高,土壤微生物CUE并未發生顯著變化[37,60]。土壤微生物CUE對溫度的響應方式受底物的組成和代謝階段的影響。?quist等[61]的研究指出,單分子結構底物供給下的土壤微生物CUE隨著溫度的升高而降低,而多聚體結構底物下的土壤微生物CUE則不隨溫度的升高而改變。一些研究還指出,長期的增溫將會使微生物產生適應性。長期增溫下微生物將降低其基礎呼吸速率[62]。在哈佛森林開展的連續增溫實驗表明,18年的5℃增溫使得土壤微生物CUE隨溫度升高而降低的程度要低于連續2年的增溫效應[6]。由于微生物的熱適應性與微生物群落組成改變、養分有效性降低、微生物代謝途徑的轉變等有關,同時也受底物和觀測方法的影響,微生物對溫度的響應及其機理仍存在很多不確定性。
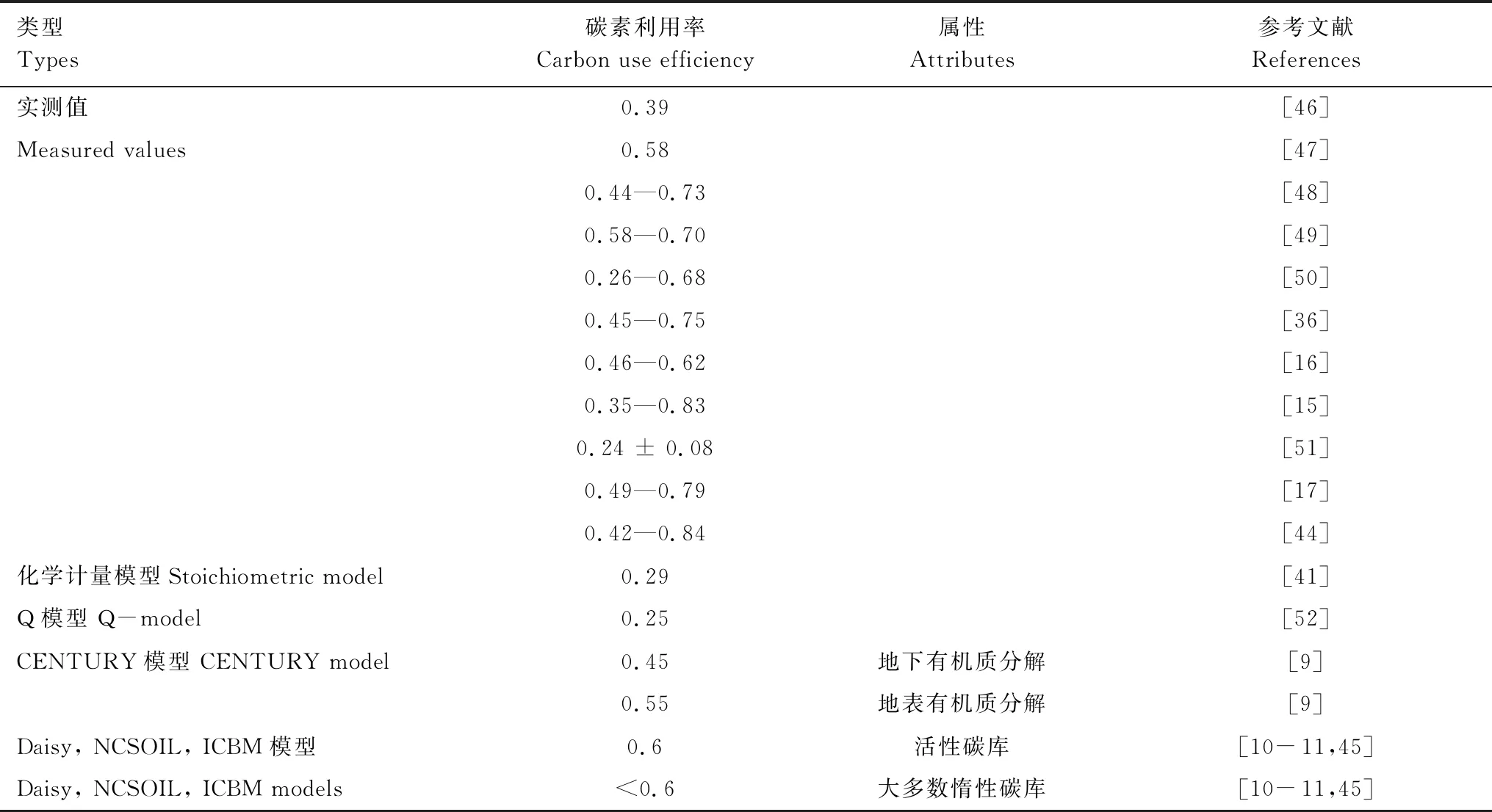
表2 微生物碳素利用率的變異性
土壤濕度和水分有效性是影響微生物生長和呼吸過程,進而影響CUE的另一重要環境因子[15]。與溫度的影響相似,土壤水分有效性對CUE的影響復雜而多變,受到土壤水分脅迫強度、時長以及土壤類型的影響。研究表明,在短期的水分脅迫下,土壤水勢的降低會激發微生物對水分脅迫的響應,促進微生物通過增加滲透壓調控或采用短期休眠來減少干旱的影響,使得土壤微生物CUE增加[16,63]。然而,長時間的水分脅迫會降低土壤底物的溶解性和吸收性,從而抑制微生物的生長[13,64]。同時,長期的水分脅迫還會增加微生物的代謝消耗,進而降低CUE[15]。研究表明,厭氧環境下的土壤微生物CUE低于有氧環境[36]。在厭氧環境下,土壤微生物的代謝產物將由CO2轉變為不能被完全氧化的CH4形式釋放,CUE降低[65]。
4.3 底物質量和有效性
微生物吸收利用的底物來自于土壤有機碳和植被凋落物,不同的底物質量將會顯著影響土壤微生物CUE[17]。這種影響源自底物的不同物質組成、分解過程、還原性程度以及有效性等。分解結構復雜的底物(如木質素和酚類等),需要經歷更多的酶促分解反應過程,增加了微生物的呼吸代謝,CUE會降低[61,66]。對難分解性底物的降解還將增加胞外酶代謝,也使得CUE降低[67]。Takriti等[44]的研究指出,土壤微生物CUE在地理分布上的分異主要受到各樣點土壤底物質量差異的影響。
底物的C還原性程度(γS)是影響土壤微生物CUE的另一個重要因素。C還原性程度是指每摩爾C的化學能,通常用每摩爾C的電子當量表示。被微生物利用的主要底物的γS通常介于3—5的范圍內(如有機酸、葡萄糖、碳水化合物和脂類),與土壤微生物的C還原性程度相當(γB≈4.2)[39]。當底物的γs低于微生物γB時,由于單位底物所具有的能量不能滿足一個單位生物量生產的能量需求,土壤微生物CUE較低[39- 40]。對16種不同還原性程度的底物CUE的分析結果表明,糖類CUE(0.667)> 氨基糖CUE(0.601)> 氨基酸CUE(0.551)> 有機酸CUE(0.498)[17]。
此外,土壤微生物CUE還會受到底物養分有效性的影響。當養分有效性和組成發生改變時,微生物會通過改變其化學酶調控的碳同化途徑來響應養分的變化[68- 70]。大量研究表明,土壤微生物CUE會隨著養分有效性的增加而增加[12- 13]。微生物碳吸收速率與養分濃度間存在飽和函數關系。在資源限制下,微生物將維持能滿足吸收系統資源消耗下的最優碳吸收速率[41,70]。當養分有效性增加,養分濃度超過微生物的平衡濃度時,將會促進碳吸收,從而提高CUE。相反,養分的限制則會降低CUE。養分的限制會改變微生物自身的分解和合成耦合代謝過程,增加胞外酶和多糖等代謝產物,使CUE降低[71- 72]。大量的N添加實驗也證明這一點,N素等養分的添加,會刺激微生物的活性或降低微生物呼吸代謝,微生物表現出高的CUE,尤其在N素限制的環境下[20,51,73- 74]。N添加主要促進了微生物對活性C的利用效率[74]。有研究也指出,長期的N添加后活性的C庫被逐漸分解,微生物群落會最終轉變為受C素等能量限制,從而土壤微生物CUE降低。由于大部分養分添加實驗仍處在有限的時間范圍,揭示微生物如何響應與適應養分有效性的變化還需要更深入的長期觀測研究。
為滿足對多種養分元素的需求,土壤微生物還會進行細胞內的資源調節和再分配,進一步影響微生物的生長和CUE[75- 78]。當受某種養分限制時,微生物會投入更多的能量來獲取缺乏的營養元素,從而抑制了微生物生長和CUE[12- 13]。例如,受P素限制時,微生物會增加用于P元素資源獲取的投入,使得CUE降低[29]。實驗表明,養分的不足將抑制微生物的生長,使得CUE降低[61]。這種養分有效性的限制作用通常是多種養分有效性共同限制的結果[29]。
4.4 生態化學計量學限制
底物的元素化學計量比的變化顯著影響微生物的生長和CUE[13,79]。分解者-微生物具有穩定的元素化學計量比,且不同于外界環境和分解底物的化學計量組成。研究表明,土壤微生物的C/N約為7—8.6[79- 81],高于浮游生物(6.6)和水生生物(8.3),而低于土壤有機碳(12.3)和植被凋落物(18.2—75)[79- 83]。土壤微生物的C/P約為23—60[79- 80],顯著低于浮游生物(106)、水生生態系統(166)、土壤有機碳(72)和植被凋落物(100—837)[79- 83]。
根據化學計量比限制理論,土壤微生物會優先吸收利用元素組成最適宜其生長所需的物質,以有利于維持自身的化學計量比平衡。當底物的C含量與必需元素E含量的比值(C∶E)低于微生物最優生長所需元素比例的閾值(TERC∶E)時,有利于元素的礦化;而當C∶E高于TERC∶E時,有利于元素的固持。因此,分解N、P元素含量高的底物,土壤微生物表現為C限制,CUE較高。當分解底物N、P元素含量低時,土壤微生物可能有兩種反應:一是抑制對底物C和養分的吸收;二是將多余的C以溢流呼吸或代謝分泌物的形式釋放[35]。第一種反應將不影響微生物的CUE,然而第二種反應由于增加了呼吸代謝和分泌物釋放,將會降低土壤微生物CUE。
大量研究指出,土壤微生物CUE與土壤有機質C∶N、C∶P比等呈負相關關系[33,42,44]。Lee 等[73]的研究發現,農田和落葉林中的土壤微生物CUE均隨著土壤可溶性C∶N的升高而降低。Takriti等[44]對不同海拔上土壤微生物CUE的變異研究也指出,在有機層和礦物層表層土壤中,隨著緯度的升高,土壤C∶N升高,土壤微生物CUE呈下降趨勢。
4.5 微生物群落組成
土壤微生物CUE還受到微生物群落結構和組成的影響,因為不同的微生物種群具有不同的有機質分解和吸收速率[84- 85]。有研究表明,采用‘機會’生長策略的快速生長型微生物通常更適應高養分含量的環境,相對于緩慢生長的微生物具有較低的CUE[86- 88]。Adu等[47]的研究也指出,以真菌為主的微生物群落的CUE往往高于細菌為主的微生物群落。真菌的C∶N∶P變異范圍通常寬于細菌,并且真菌的C∶N高于細菌具有更高的C素需求,表現出高的CUE[87]。但是,也有研究指出,土壤群落的CUE并無顯著差異[19- 20]。Thiet等[20]在農田生態系統的研究中發現,高真菌/細菌比的群落與低真菌/細菌比的群落的CUE并無顯著差異,分別為0.59±0.02和0.61±0.01。有研究也指出,底物C∶E的增加可以增加真菌群落的CUE,但卻降低細菌群落的CUE[87]。微生物群落的種間競爭會降低微生物的CUE[89]。由于微生物的群落組成極易受到外界環境變化以及人為活動的影響,也隨著底物質量和組成的變化而變化,量化不同群落組成的CUE的差異仍需進一步深入研究[19,87]。
5 問題與展望
5.1 研究尺度辨析
微生物CUE通指微生物吸收利用C并將其轉化為自身產物的效率,但它卻具有特定的時間和空間屬性[90]。Geyer等[91]將微生物CUE分為3個層次:種群CUE(CUEP)、群落CUE(CUEC)和生態系統CUE(CUEE)。這3個層次的CUE表征了從種群到生態系統不同尺度上微生物的生物合成效率,包含了不同的微生物生理和化學過程[91]。前期大量的研究更多關注的是種群和群落尺度的微生物CUE,近期有越來越多的研究著眼于探討生態系統尺度CUE的變化及其對環境的響應[92- 93]。不同研究尺度、觀測階段和測定條件下獲取的CUE結果比較將給解析結果帶來更大難度和不確定性[90]。在未來的CUE研究中,亟需更清晰地辨明微生物CUE的特定研究尺度和過程以準確地比較和應用。
5.2 代謝組分分析
當前土壤CUE的計算方法不統一并且存在高估現象。微生物的呼吸是多個組分的呼吸的總和,包括了用于生長的生長呼吸、有機體運轉的維持呼吸、分泌胞外酶的呼吸以及溢流的呼吸。當前微生物CUE研究大多只考慮了微生物的生長呼吸或生長和維持呼吸,極少考慮胞外酶和溢流呼吸組分[13]。這使得CUE被高估,同時也使得CUE的計算結果存在很大差異。在今后的研究中,還需充分考慮胞外酶和溢流呼吸的損失。例如,在室內培養的測定過程中,選取適宜的培養時長觀測包含微生物產物代謝和分泌的完整過程。此外,采用同位素標記與代謝組學相結合方法測算酶的產量和轉化。
5.3 多要素交互影響研究
由于復雜的土壤-植物-微生物交互作用,環境變化對土壤微生物CUE的影響以及土壤微生物CUE對全球變化的響應與反饋仍存在很大的不確定性。例如,土壤微生物CUE是否隨著溫度升高而持續降低?是否存在熱適應性?長期的外源養分輸入下土壤微生物CUE將如何變化?等。由于土壤微生物CUE對環境變化的響應與適應因不同的底物質量、有效性、元素化學計量組成、微生物活性、群落組成而復雜多變[58],在未來的過程機理研究中,還需要綜合探討環境和生物多要素交互影響下,微生物的生理響應過程與對CUE的影響和調節機制。
5.4 土壤碳循環模型優化
土壤微生物CUE是土壤C循環模型中的重要參數。現有許多模型假定CUE是一個常數或很少考慮,使得無法準確模擬土壤微生物CUE對外界環境條件變化的動態響應過程及其對土壤C循環的影響[7,94]。有的模型則將微生物CUE表達為溫度、養分有效性或養分元素比的一階函數關系,而忽略了微生物的生理過程,如微生物代謝途徑、群落組成、種間關系等的變化。此外,大量生物地球化學模型中,微生物呼吸過程通常被認為主要與微生物C平衡相關,而與N平衡無關,更是缺乏考慮P元素的動態變化的影響[29,35]。在未來的土壤C周轉模型優化中,還需綜合考慮元素計量平衡制約、微生物對外界環境條件變化的動態響應過程,以及微生物代謝途徑、群落組成、種間關系、微生物周轉等生理過程變化。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
