刺盤孢菌真菌病毒研究進展
2020-03-20 03:44:50李春霞李敏高兆銀弓德強洪小雨姜瑛常圣鑫胡美姣
南方農業學報 2020年1期
李春霞 李敏 高兆銀 弓德強 洪小雨 姜瑛 常圣鑫 胡美姣
摘要:真菌病毒是指寄生于各種真菌中的病毒,其中部分弱毒相關病毒可使寄主真菌致病力發生衰退,甚至導致病原群體致病力下降,是一種具有生防潛能的微生物資源。刺盤孢菌(Colletotrichum spp.)可引起植物嚴重的炭疽病,造成巨大的經濟損失。文章歸納總結了刺盤孢菌中真菌病毒的種類和特征:僅有9種刺盤孢菌中存在真菌病毒,且病毒基因組類型均為dsRNA;絕大部分病毒為直徑25~50 nm的球狀病毒顆粒,另有目前唯一一例線狀真菌病毒;序列已知的6例刺盤孢菌病毒中有2例為雙分病毒、2例為產黃青霉病毒、2例還未確定歸類。通過分析病毒對寄主真菌的影響發現不同病毒對刺盤孢菌表型、生長、產孢和致病力等方面的影響存在明顯差異,但僅有2例為能使寄主致病力衰退的弱毒相關病毒。弱毒相關病毒的發現為植物炭疽病的生防研究提供了新契機,但由于存在真菌病毒數量不足、真菌病毒分離鑒定進展緩慢、真菌病毒傳播效率低及病毒與刺盤孢菌間的互作研究不夠深入等問題,刺盤孢菌應用于田間炭疽病的防治還有一定距離。針對上述問題,未來研究方向可從以下方面著手:采用宏轉錄組等技術鑒定和發掘更多的真菌病毒;通過遺傳轉化手段或挖掘可利用的化學物質或傳播介體提高病毒的傳播效率;建立刺盤孢菌遺傳轉化體系及病毒反向遺傳轉化系統、結合多組學手段深入研究病毒與寄主真菌互作,為刺盤孢菌的致病機理研究及真菌病毒的生防利用提供參考。
關鍵詞: 刺盤孢菌;真菌病毒;炭疽病;生物防治
中圖分類號: S476? ? ? ? ? ? ? ? ? ? ? ?文獻標志碼: A 文章編號:2095-1191(2020)01-0123-10
Abstract: Mycoviruses are parasitic viruses in various fungi. Some hypovirulence-associated mycoviruses are microbial resources with the potential of biocontrol, which reduce the pathogenicity of plant pathogenic fungi and even cause the decline of pathogenicity of pathogenic groups. Colletotrichum spp. can cause severe anthracnose in plants, leading to enormous economic loss. This paper summarized the types and characteristics of mycoviruses in Colletotrichum spp. from literature. Currently, it has been found that mycoviruses infected only nine species of Colletotrichum spp., and their genome typesare dsRNA. Most of the mycoviruses are a spherical particle with the diameter of 25-50 nm, but a filamentous mycovirus is found for the first time in the world. Among the six known sequence cases, two cases are partitivirus, two cases are chrysovirus, and two cases are unclassified. By summing up the effects of viruses on host fungi, it is shown that there are differences in phenotype, growth, sporulation, and pathogenicity among different mycoviruses but only two mycoviruses could make host pathogenicity decline. The discovery of hypovirulence-associated mycovirus provides a new opportunity for the biocontrol of plant anthracnose, but there is still a long way to go for the control of anthracnose in the field with the following reasons:the quantity of mycoviruses is insufficient; the progress of isolation and identification of mycoviruses is slow; the transmission of fungal viruses is insufficient; the interaction study between viruses and Colletotrichum spp. is not thorough enough; a long distance between the study and its application to field control. Given the above problems, it was suggested that researchers could carry out their researches from the following aspects:identifying and discovering more mycoviruses in Colletotrichum spp. by metatranscriptome technology; improving virus transmission efficiency by using genetic transformations or digging available chemicals or transmission mediators; establishment of Colletotrichum spp. genetic transformation system and mycovirus reverse genetic transformation system combine with multi-o-mics methods to study the interaction between mycoviruses and Colletotrichum spp., providing references for the study of pathogenic mechanism of Colletotrichum spp. and biocontrol of mycoviruses.
Key words: Colletotrichum spp.; mycoviruses;anthracnose; biocontrol
Foundation item: Hainan Natural Science Foundation(319MS095); Chinese Academy of Tropical Agricultural Scien-ces Basic Research Special Project(1630042019041)
0 引言
刺盤孢菌(Colletotrichum spp.)隸屬于半知菌亞門腔孢綱黑盤孢目黑盤孢科刺盤孢屬真菌,其有性階段是子囊菌亞門的小叢殼屬或無柄盤菌屬,是禾本科、果樹、蔬菜等經濟作物的寄生菌,可引起嚴重的炭疽病(周德慶和徐士菊,2005)。據統計,受刺盤孢菌侵染的影響,熱帶、亞熱帶及地中海地區出口水果品質顯著下降,所造成的經濟損失甚至可達80%以上(Joshi,2018),《2016全球植物現狀評估報告》(Willis and Bachman,2016)更是將刺盤孢菌列為十大影響植物的病原真菌之首,可見其對世界經濟和科學研究的重要性。
真菌病毒(Fungal virus或Mycovirus)是指寄生在真菌中的一系列病毒,它們廣泛地存在于各種真菌中。迄今為止,在美國國家生物技術信息中心(NCBI)數據庫可查詢到超過250種真菌病毒被成功測序并注冊(Xie and Jiang,2014)。國際病毒分類委員會(ICTV)將已發現的真菌病毒劃分為17個類群,包括16個科和1個不包括在科里的屬(Kotta-Loizou and Coutts,2017),根據基因組類型可分為雙鏈(ds)RNA病毒(8個類群):Botybirnavirus、Endornaviridae、Partitiviridae、Megabirnaviridae、Reoviridae、Chysoviridae、Totiviridae和Quadriviridae;單股正鏈RNA[(+)ssRNA]病毒(6個類群):Alphaflexiviridae、Gammaflexiviridae、Hypoviridae、Pseudoviridea、Barnaviridae和Metaviridea;單股負鏈[(-)ssRNA]病毒(1個類群):Mymonaviridea;環狀ssDNA病毒(1個類群):Genomoviridea。研究發現有部分真菌病毒可降低植物病原真菌的致病力,甚至使病原群體致病力發生衰退,這種特殊的致病功能使得真菌病毒兼具理論研究的重大意義和應用開發的廣泛前景,為農業綠色可持續發展提供堅實基礎,是一種有待開發的、具有生防潛能的微生物資源。
本研究對刺盤孢菌中真菌病毒的種類和特征進行歸納總結,分析弱毒相關病毒對刺盤孢菌表型、生長、產孢及致病力等方面的影響,指出利用真菌病毒防治植物炭疽病的優勢和存在的問題,展望未來研究方向,為刺盤孢菌真菌病毒的深入研究和利用提供參考。
1 刺盤孢菌相關真菌病毒
早在1975年Moffitt和Lister就應用血清學篩選試驗在高粱刺盤孢病菌(C. graminicola)和甘蔗刺盤孢病菌(C. falcatum)中檢測到dsRNA真菌病毒。同年,Rawlinson等(1975)從菜豆刺盤孢病菌(C. lindemuthianum)菌株中提取獲得dsRNA,電鏡觀察到直徑30 nm的等距病毒粒子。截至目前,分別在尖孢刺盤孢(C. acutatum)、山茶刺盤孢(C. camelliae)、鐮形刺盤孢(C. falcatum)、果生刺盤孢(C. fructicola)、盤長孢狀刺盤孢(C. gloeosporioides)、禾生刺盤孢(C. graminicola)、希金斯刺盤孢(C. higginsianum)、豆刺盤孢(C. lindemuthianum)和平頭刺盤孢(C. truncatum)等9種刺盤孢菌中發現真菌病毒,且基因組類型均為dsRNA,絕大部分病毒為直徑25~50 nm的球狀病毒顆粒;此外,在山茶刺盤孢中發現目前唯一一例線狀真菌病毒。以下將已發表的刺盤孢菌病毒分為序列已知和序列未知兩類進行介紹。
1. 1 序列已知的刺盤孢菌病毒
1. 1. 1 C. acutatum partitivirus 1(CaPV1) Zhong等(2014)從辣椒果實上分離的尖孢刺盤孢中發現直徑約40 nm的球形病毒樣顆粒,分離檢測后獲得2條dsRNA條帶,大小分別為1762和1381 nt;分析發現這2條dsRNA條帶各包含一個開放閱讀框(ORF),分別編碼62和40 kD蛋白;其中,62 kD蛋白與部分病毒的RNA依賴RNA聚合酶(RdRp)相似,而40 kD蛋白與所有數據庫中已發表的任何衣殼蛋白(CP)均無顯著相似性;基因組比對和系統發育分析表明,該病毒與雙分病毒科的真菌病毒密切相關,因此,將其命名為C. acutatum partitivirus 1(CaPV1)。
1. 1. 2 C. higginsianum Non-segmented dsRNA virus 1(ChNRV1) Campo等(2016)發現希金斯刺盤孢菌株存在基因組未分段的dsRNA真菌病毒,命名為C. higginsianum Non-segmented dsRNA virus 1(ChNRV1);分析發現該dsRNA大小為2923 nt,包含兩個不同的ORF:ORF1編碼一個假定的CP,而ORF2編碼一種假定的RdRp;通過對病毒RdRp進行最大似然系統發育分析發現ChNRV1既不屬于Totiviridae也不屬于Partitiviridae,而被歸為Partitiviridae家族的姐妹支——Amalgaviridae,作者認為ChNRV1要么是Amalgaviridae的新成員,要么屬于一個尚未確定的家族。
1. 1. 3 C. truncatum partitivirus 1(CtParV1) Marzano等(2016)發現平頭刺盤孢菌株中存在兩個與Partitiviridae親緣關系較近的dsRNA序列:dsRNA1(1820 nt,編碼病毒RdRp蛋白)和dsRNA2(1535 nt,編碼病毒CP蛋白),這兩個dsRNA的序列長度及其編碼的多肽長度均與gammapartitivtivirus屬病毒相近,且dsRNA1與gammapartitivtivirus屬Verticillium dahliae partitivirus 1 RNA1的相似性達87%,因此將該病毒命名為C. truncatum partitivirus 1。
1. 1. 4 C. gloeosprioides chrysovirus 1(CgCV1)
Zhong等(2016)從盤長孢狀刺盤孢中分離出一種新型dsRNA真菌病毒(C. gloeosprioides chrysovirus 1,CgCV1),由3個dsRNA基因組片段組成(dsRNA 1~dsRNA 3),長度分別為3397、2869和2630 bp,各包含一個ORF,推定dsRNA 1~dsRNA 3分別編碼RdRp、CP和蛋白酶;CgCV1 RdRp的系統發育分析表明該病毒與Fusarium oxysporum chrysovirus 1密切相關;根據病毒基因組片段大小、序列和結構特征,RdRp的氨基酸序列相似性,特定基序的特征,病毒粒子的大小和結構特征,作者認為CgCV1是Chrysovirus屬的一個新物種。
1. 1. 5 C. camelliae filamentous virus 1 (CcFV-1)
Jia等(2017)在山茶刺盤孢菌中發現一種長達4661.6 nm的線狀病毒,命名為C. camelliae filamentous virus 1(CcFV-1),是目前唯一一例線狀真菌病毒;CcFV-1有8個dsRNA,長度分別為990、1053、1085、1122、1299、2012、2253和2444 bp,共編碼10個ORF,其中ORF1編碼一個RdRp,ORF4編碼CP;系統發育分析發現CcFV-1與Aspergillus fumigatus tetramycovirus-1和Beauveria bassiana polymycovirus-1有一定的系統親緣關系,但在病毒形態和基因組組成上存在差異;CcFV1病毒的基因組大小不到3000 bp,卻包被在長約4000 nm的病毒中,這可能是一種新的病毒包裝機制。CcFV1病毒的發現對進一步了解RNA病毒的形態、進化及分子特性等具有重要意義。
1. 1. 6 C. fructicola chrysovirus 1(CfCV1) Zhai等(2018)從果生刺盤孢菌株中分離并鑒定出一種新型7段dsRNA病毒,命名為C. fructicola chrysovirus 1 (CfCV1)。CfCV1為直徑35 nm的球狀病毒粒子;dsRNA 1~dsRNA 7的全長分別為3620、2801、2687、2437、1750、1536和1211 bp;其中,dsRNA 1編碼RdRp,dsRNA 2編碼CP,dsRNA 5編碼一種C2H2型鋅指蛋白,dsRNA 3、dsRNA 4和dsRNA 6編碼其他蛋白的功能尚不清楚;基于RdRp和CP的系統發育分析表明CfCV1與Botryosphaeria dothidea chrysovirus 1和Penicillium janczewskii chrysovirus 2有系統親緣關系,同屬于產黃青霉科病毒。
1. 2 序列未知的刺盤孢菌病毒
除上述刺盤孢菌真菌病毒的基因組已進行測序外,還有一些真菌病毒基因組未完成測序,僅觀察到類似病毒的顆粒和/或提取到部分dsRNA片段。如Moffitt和Lister(1975)、Rawlinson等(1975)使用血清學方法檢測發現禾生刺盤孢、鐮形刺盤孢和豆刺盤孢含dsRNA真菌病毒,通過電鏡可觀察到豆刺盤孢的病毒粒子為直徑30 nm的等距粒子。Dale等(1988)使用纖維素柱層析法提取了14株盤長孢狀刺盤孢菌株的dsRNA,發現菌株所含dsRNA的數量和大小與炭疽病類型存在相關性。Figueirêdo等(2012)利用纖維素柱層析法從腰果刺盤孢病菌(C. gloeosporioides)菌株上分離得到9條dsRNA,透射電鏡可觀察到分離所得的RNA為直徑30~35 nm的球形病毒顆粒。Liao等(2012)利用纖維素粉抽提法提取3株不同致病力的尖孢刺盤孢病菌的dsRNA,發現不同致病力菌株中dsRNA的組成和大小存在差異,其中弱致病力菌株Coll-365中包含2條大小分別為1300和2500 bp的dsRNA,而強致病力菌株Coll-524中為1400和1700 bp的2條dsRNA條帶。Lima等(2012)對39株巴西胡椒刺盤孢病菌(C. gloeosporioides)進行dsRNA提取,只在3株中發現1條3000 bp的dsRNA條帶。
2 真菌病毒對刺盤孢菌的影響
2. 1 表型
Zhong等(2014)通過環已胺法脫除尖孢刺盤孢病菌菌株中的CaPV1病毒,獲得2株脫毒菌株,在感染病毒和脫除病毒的分離株之間,菌落表型在色素沉著和菌絲生長上有輕微差異,脫毒菌株黑色素積累顯著減少,菌絲生長松散或稀疏并出現隆起的同心輪紋圈;Jia等(2017)通過對含毒山茶刺盤孢菌株進行單孢分離獲得無毒菌株,發現脫毒后的菌株呈現隆起的同心輪紋圈,且分泌黑色素減少;而轉染CcFV-1后,菌落形態恢復至與帶毒親本相似,即菌株輪紋圈消失,黑色素增加,且隨著培養時間延長(15 d),黑色素累積顯著增多,說明病毒的存在顯著影響了寄主真菌的菌落形態。但也有研究指出病毒存在對果生刺盤孢菌株的菌落形態無明顯影響(Zhai et al.,2018),說明不同病毒粒子對寄主真菌的影響存在差異。
2. 2 生長
Jia等(2017)通過對山茶刺盤孢菌株和脫毒菌株進行生長速度測定,結果顯示3株脫毒菌株相比帶毒菌株生長速度變快(6.3~6.5 mm/d);轉染CcFV-1后,3株菌株的生長速度變慢(5.6~5.9 mm/d),與帶毒菌株LT-3-1(生長速度5.4 mm/d)近似。而Zhai等(2018)通過對帶毒果生刺盤孢菌株進行單孢分離獲得脫毒菌株,發現帶CfCV1病毒菌株與無毒菌株間菌株生長速率差異不大。
2. 3 產孢
Zhong等(2014)研究發現在尖孢刺盤孢病菌分生孢子的產生方面,帶毒菌株與脫毒菌株之間存在顯著差異,即脫毒菌株的產孢量幾乎是帶毒菌株HNZJ001的3倍,意味著病毒可能對尖孢刺盤孢病菌有一定的影響,至少在分生孢子的產生方面,這可能有利于這種病原菌的廣泛傳播。與之相反,Campo等(2016)采用環己胺處理獲得1株ChNRV1病毒固化/脫除的野生型C. higginsianum菌株IMI 349063A,研究發現無病毒菌株產孢水平是含毒菌株的8.8倍,證實C. higginsianum菌株產孢缺陷與ChNRV1病毒有關。
2. 4 致病力
前人研究發現有一類病毒能使寄主真菌出現產孢能力下降、生長速率減慢、致病力降低等癥狀,這一類真菌病毒稱為弱毒相關病毒(Hypovirulence-associated mycovirus)。目前,在刺盤孢菌中僅發現2例能使寄主致病力衰退的弱毒相關病毒:Jia等(2017)通過對帶毒山茶刺盤孢菌株和脫毒菌株進行致病力測定,發現3株脫毒菌株相比帶毒菌株致病力增強(6.1~14.9 mm);而轉染CcFV-1后,3株菌株的致病力減弱(1.0~4.0 mm),與帶毒菌株LT-3-1(病斑長度1.0 mm)近似,因此作者認為CcFV-1通過損害山茶刺盤孢細胞的動態平衡引發弱毒現象,從而影響寄主形態,如色素生產和分配、生長率和菌絲輪紋。值得一提的是,CcFV-1裸露的dsRNA也具有感染性,可侵染真菌原生質體誘導其產生并積累病毒顆粒,同時,被CcFV-1病毒感染后的真菌仍能維持生長活動,CcFV-1這種強傳播又不影響宿主生存的能力使其具有顯著的生防潛能。Zhai等(2018)通過對帶毒果生刺盤孢進行單孢分離獲得脫毒菌株,發現帶CfCV1病毒菌株與脫毒菌株間菌落形態和菌株生長速率差異不大,但對離體果實的弱毒力作用較無CfCV1菌株強。雖然Liao等(2012)研究發現帶毒尖孢刺盤孢Coll-365對辣椒的致病力顯著低于Coll-153(無毒)和Coll-524(帶毒),侵染辣椒果實96 hpi后仍無法產生分生孢子盤,但作者并未對菌株進行脫毒或將病毒轉入無毒菌株,因此不能確定菌株Coll-365所帶病毒為弱毒相關病毒。
2. 5 其他
病原真菌分泌的角質酶、蛋白酶及漆酶等可降解寄主植物的細胞壁和角質層等,以利于病原真菌的侵入、定殖與擴展。另外,真菌附著胞還可通過在細胞內積累大量的丙三醇從而形成巨大的膨脹壓力幫助其穿透植物寄主角質層啟動侵染植物的進程(Money,1999)。Liao等(2012)檢測發現攜帶不同類型dsRNA的尖孢刺盤孢具有不同的酶活力和細胞膨壓,其中弱致病力帶毒菌株Coll-365具有較高的蛋白酶和角質酶活性,而強致病力帶毒菌株Coll-524的漆酶活性最高,且Coll-365的細胞膨壓(<4.9 Mpa)低于Coll-524和無毒菌株Coll-153(>6.0 Mpa)。同樣由于Liao等(2012)未對帶毒菌株進行脫毒或對無毒菌株進行轉染,因此無法準確證實病毒與菌株酶活力和細胞膨壓的相關性。Zhong等(2014)研究也指出攜帶CaPV1病毒的尖孢刺盤孢菌株與脫毒菌株的漆酶活性無明顯差異。Campo等(2016)發現ChNRV1病毒粒子的積累量受真菌DCL1和AGO1蛋白的負向調控,但病毒粒子并未被真菌完全清除,作者猜測ChNRV1可能在尚未研究的環境中為尖孢刺盤孢提供了一些優勢,也可能作為載體促進真菌基因組的可塑性,通過進化促進新的毒力性狀出現。Jia等(2017)對比脫毒菌株與帶毒菌株的菌絲超微細胞結構,在帶毒菌株菌絲體細胞中發現大量的為病毒提供復制和組裝場所的囊泡結構,缺少液泡和伏魯寧體,并且囊泡內部還觀察到疑似病毒樣顆粒,說明CcFV-1病毒的存在顯著影響了菌絲的細胞結構。
如上所述,確實有部分病毒對寄主真菌的產孢能力、生長速率、致病力等存在顯著影響,但也有研究表明某些真菌病毒對寄主沒有明顯的影響(Buck,2017)。在菜豆刺盤孢病菌中,Rawlinson等(1975)對比同一株系的帶毒菌株與無毒菌株,發現病毒的存在并不會對菜豆刺盤孢病菌菌株/菌落的形態、產孢特性及菌株致病力產生影響,甚至在不同碳源培養基(無碳源或1%葡萄糖、聚果膠或纖維素的Mathur培養基)中,帶毒菌株與無毒菌株的菌絲生長速率均無明顯差異。
3 利用真菌病毒防治植物炭疽病存在的問題與前景展望
3. 1 存在問題
刺盤孢屬真菌的種類多達600余種,且多數種類可引起植物嚴重的炭疽病,造成巨大損失(Jaya-wardena et al.,2016;Carbú et al.,2019)。目前植物炭疽病的防治主要通過化學藥劑,若藥劑施用不當,不僅防效顯著下降,還將污染環境,影響人類健康。利用弱毒相關病毒防治植物真菌病害不僅綠色安全高效,而且與其他生物防治方法相比具有明顯優勢:首先,弱毒相關病毒一旦進入寄主病原菌即可抑制病斑擴展,防效精準快速;其次,含弱毒相關病毒的弱致病力菌株進入寄主植物后不僅不會傷害寄主植物,還會誘導寄主植物產生防御反應,增強寄主對強致病力菌株的抗性;再者,弱毒相關病毒一旦進入農田生態系統,既可長久定殖于病原菌體內,又可在不同菌株間進行傳播,防效長、作用范圍廣(Xie and Jiang,2014)。最經典的是1965年Grente等通過寄生板栗疫病菌(Cryphonectria parasitica)的弱毒相關病毒C. parasitica hypovirus 1防治歐洲板栗疫病的蔓延,直至1994年,Heiniger和Rigling對當時歐洲板栗疫病情況的調查綜述中還指出,含有弱毒相關病毒的低毒菌株不僅成功控制了歐洲板栗疫病病情,還顯著降低了板栗疫病的嚴重性,由此可見真菌病毒確實具有巨大的生防潛能。但是,利用真菌病毒防治植物炭疽病目前仍存在許多問題,主要表現為:
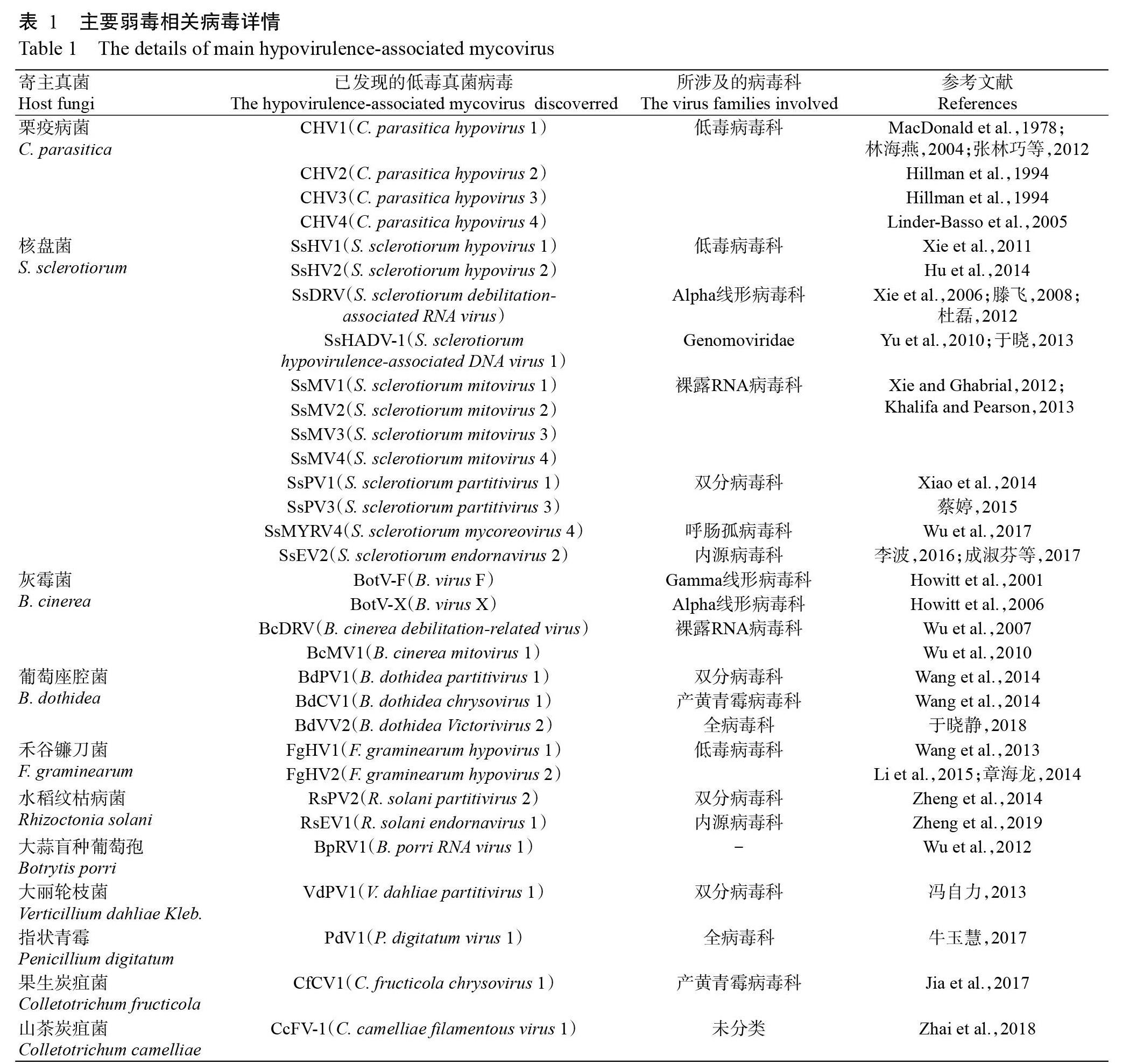
(1)刺盤孢菌真菌病毒數量不足。目前,有超過250種真菌病毒被成功測序并在NCBI注冊(Xie and Jiang,2014),其中可引起植物病原真菌致病力衰退的弱毒相關真菌病毒至少有30余例(表1),其寄主包括栗疫病菌(C. parasitica)、核盤菌(Sclerotinia sclerotiorum)、灰霉菌(Botrytis cinerea)、葡萄座腔菌(Botryosphaeria dothidea)、禾谷鐮刀菌(Fusarium graminearum)和水稻紋枯病菌(Rhizoctonia solani)等10余種真菌。由表1可知,核盤菌中發現的弱毒相關病毒數量較多,種類豐富,為防治菌核病提供了堅實后盾。相比于核盤菌,600多種刺盤孢菌中僅有9種刺盤孢菌報道發現真菌病毒顯得遠遠不夠,且僅有2例弱毒相關病毒能降低寄主致病力,因此,還需進一步挖掘刺盤孢菌中的真菌病毒,為防治植物炭疽病提供支撐。
(2)真菌病毒分離鑒定進展緩慢。早期刺盤孢菌真菌病毒的分離鑒定大多是通過顯微鏡觀察和/或血清學檢測法(Moffitt and Lister,1975;Rawlinson et al.,1975;Figueirêdo et al.,2012;Zhong et al.,2014),針對基因組為dsRNA類型的真菌病毒也可使用纖維素粉抽提結合酶解法獲得菌株內的dsRNA病毒(Liao et al.,2012;Rosseto et al.,2016;Zhong et al.,2016)。顯微鏡鑒定結果準確,但對儀器設備要求較高,血清學可批量檢測樣品,但對微量病毒檢出率低,如Lima等(2012)、Rosseto等(2016)研究中刺盤孢菌真菌病毒檢出率較低,分別為7.7%(3/39)和3.85%(1/26),原因可能是刺盤孢菌中仍有部分真菌病毒基因組類型為單鏈RNA或DNA。此外,在顯微鏡下未觀察到病毒顆粒并不意味著沒有真菌病毒。正如Nuss(2005)所描述,這可能是因為許多真菌病毒并不編碼衣殼蛋白,而是以裸露的dsRNA分子存在,因此,醋酸雙氧鈾無法染色病毒蛋白,從而無法觀察到病毒顆粒,而纖維素粉抽提結合酶解法僅局限于dsRNA病毒,因此,還需探索更高效準確的真菌病毒檢測鑒定方法。
(3)真菌病毒傳播效率低。真菌病毒粒子普遍不具有侵染性,不能在體外對寄主真菌進行傳播,而真菌的細胞壁也在一定程度上攔截了真菌細胞對病毒的攝入。因此,大部分真菌病毒只能通過寄主細胞分裂和孢子產生的垂直傳播方式或菌絲細胞融合的水平傳播方式在細胞內傳播,其中垂直傳播主要通過產生無性孢子進行(Varga et al.,2003),而水平傳播僅限于在遺傳上可親和的菌株間進行(Hutchison et al.,2009)。Hutchison等(2009)研究表明,營養體不親和的菌株在菌絲接觸時會出現程序性細胞死亡現象,使真菌病毒不能通過菌絲融合進行水平傳播。另外,攜帶弱毒相關病毒的真菌菌株一般生命力弱、不易存活,在自然環境中競爭力差,進一步影響了病毒的傳播力。所以傳播效率不足是真菌病毒生防應用的主要限制因素。在核盤菌中已發現病毒SsHADV-1不僅能在寄主細胞內傳播,降低寄主致病力,還可通過純病毒顆粒和基因組DNA進行體外侵染,抑制PDA上核盤菌的生長,或接噴在擬南芥和甘藍油菜葉子上,可感染無病毒油菜菌核病菌絲,減緩發病程度,提高野外培養油菜籽的產量(Yu et al.,2010,2013;于曉,2013);而病毒SsMYRV4可作為其他病毒粒子傳播的橋梁,介導弱毒相關病毒在不同菌株個體間傳播,降低病原菌群體的致病力(Wu et al.,2017)。在刺盤孢菌中,僅發現一例病毒粒子裸露的dsRNA具有感染性,可通過原生質體融合侵染真菌原生質體,誘導其產生并積累CcFV-1病毒顆粒(Jia等,2017),但未有研究指出其具有田間防治效果,值得進一步探究。因此,還需進一步發掘具有高傳播效率的真菌病毒資源及探索更高效的輔助病毒傳播的方法,提高真菌病毒應用潛力和價值。
(4)真菌病毒與刺盤孢菌間的互作研究不夠深入。大量研究表明,弱毒相關病毒的侵染將造成寄主的基因表達異常、蛋白質合成受阻或代謝等功能發生改變。攜帶弱毒相關病毒的弱毒株可視為遺傳背景相同且無病毒的強毒株在毒力調控途徑中發生突變的突變體,故可通過比較強毒株與遺傳背景相同的弱毒株之間的表達差異來研究影響真菌毒力的因子(康寧,2004)。因此,弱毒相關病毒除具有應用于生物防治的潛力外,還可作為模式病毒用于病毒與寄主的互作以及植物病原真菌的致病機理研究(李文靜等,2015)。目前,研究弱毒相關病毒與寄主真菌互作相對成熟的系統主要包括:CHV1-栗疫病菌互作系統、SsDRV-核盤菌互作系統、FgV1-鐮刀菌互作系統和BVX/BVF-灰霉菌互作系統(劉忱等,2018)。在刺盤孢真菌病毒的報道中僅有Zhong等(2014)、Campo等(2016)、Jia等(2017)、Zhai等(2018)的研究成功獲得脫毒菌株,通過對比帶毒菌株與脫毒菌株,驗證病毒粒子對寄主真菌的影響;只有Campo等(2016)研究了寄主真菌對病毒粒子的影響,指出C. higginsianum中涉及ChDCL1和ChAGO1的RNA沉默可能具有抗病毒作用;僅有Jia等(2017)報道將CcFV-1病毒dsRNA成功轉染了脫毒C. camelliae菌株。由此可見,病毒與刺盤孢菌間的互作研究仍需進一步深入。
3. 2 前景展望
3. 2. 1 病毒挖掘 近期研究表明,利用宏轉錄組技術可不依賴于病毒培養及病毒序列,直接以環境(樣本)中病毒核酸為研究對象,快速準確地鑒定出樣本中所有的病毒序列組成,為病毒資源的挖掘與研究提供了極大便利(Shi et al.,2016)。宏轉錄組是一門研究環境中總RNA的技術,由于真菌病毒基因組多為dsRNA和ssRNA類型,因此,宏轉錄組技術的開發和應用對刺盤孢菌真菌病毒研究具有重要意義。Marzano等(2016)研究指出,相比傳統先富集樣本中病毒再提取病毒RNA進行測序的方法,宏轉錄組技術直接提取真菌總RNA進行測序,避免了富集過程造成大量病毒序列信息的損失。Mu等(2017)利用該技術對澳大利亞84株核盤菌進行檢測,發現57例真菌病毒,包括32例新病毒,為植物菌核病的防治提供了豐富的生防資源。筆者通過宏轉錄組測序(數據未發表)發現弱致病力芒果刺盤孢病菌(Colletotrichum spp.)中的病毒基因組序列占比遠高于強致病菌株,且弱致病力菌株中平均70%的病毒基因組序列為Partitiviridae,而強致病力菌株中未發現Partitiviridae病毒序列,由此推定弱致病力菌株中可能存在能引起致病力衰退的弱毒相關病毒,且該病毒與Partitiviridae親緣關系較近。因此,利用宏轉錄組技術可快速鑒定刺盤孢菌中的真菌病毒,挖掘更多具有生防潛力的真菌病毒,促進植物炭疽病的生防研究。
3. 2. 2 提高病毒傳播效率 除篩選具有強侵染力的弱毒相關真菌病毒資源外,還可通過人為技術手段打破菌株營養體不親和限制,幫助病毒粒子轉移和繁殖。(1)利用化學物質。Hutchison等(2009)研究發現,磷脂酰肌醇、活性氧(ROS)及Ca2+信號途徑均與粗糙脈孢霉的營養不親和性密切相關。Kenichi等(2013)研究發現,鋅的化合物可減弱白紋羽病菌菌株間的營養體不親和性,提高菌株間病毒粒子的傳播效率。因此,Xie和Jiang(2014)推測,ROS清除劑或可用于克服真菌間的營養不親和性。(2)遺傳轉化。通過原生質體融合(Lecoq et al.,1979)、電擊法(Chen et al.,1994)、聚乙二醇(PEG)介導(Varga et al.,1994)、基因槍轟炸(Sasaki et al.,2002)和農桿菌介導(于曉,2013)等方法將病毒粒子/病毒RNA/病毒cDNA轉入真菌原生質體獲得可復制的病毒,以克服營養體非親和性限制,實現病毒粒體在不同菌株間的擴散,甚至在遠緣不同種之間轉移,拓寬病毒的寄主范圍。另一方面,Zhang和Nuss(2016)、Stauder等(2018)通過破壞板栗疫病菌的多個營養不親和位點打破菌株間的營養不親和,使改造后的病原菌成為低毒病毒的超級供體,通過菌絲融合將病毒傳播至不同菌株,用于野外板栗疫病菌的防治。(3)挖掘傳播介體。Liu等(2016)研究發現厲眼蕈蚊幼蟲食用了感染SsHADV-1病毒的核盤菌后,SsHADV-1會在厲眼蕈蚊體內及其細胞中復制積累,然后再經厲眼蕈蚊轉移至其他不攜帶病毒的核盤菌菌株。因此,食用真菌的昆蟲或可發掘成為真菌病毒傳播介體。
3. 2. 3 建立真菌和病毒的遺傳操作系統 栗疫病菌、核盤菌、灰霉菌和禾谷鐮刀菌的基因組均為單倍體,遺傳性狀穩定,不僅建立了原生質體再生、基因敲除或互補等分子遺傳轉化體系,還建立了病毒侵染性克隆和高效病毒反向遺傳操作系統,可簡便、快速地克隆和鑒定出寄主靶基因。由此可知,建立真菌遺傳操作系統和病毒反向遺傳學系統,是研究寄主真菌與真菌病毒互作機制的首要條件(Nuss,2011)。迄今已有27種刺盤孢菌基因組(https://www.ncbi.nlm.nih.gov/genome)被測定,同時,盤長孢狀刺盤孢菌、瓜類刺盤孢菌和禾生刺盤孢菌等多種刺盤孢菌的遺傳轉化體系已成功構建(Yan et al.,2018),這為進一步研究刺盤孢菌與病毒的分子互作提供了有力支持。但刺盤孢菌中病毒反向遺傳學系統仍存在許多困難需要克服,如病毒侵染性克隆、轉化及病毒基因組/病毒粒子轉染等,因此,還需認真借鑒CHV1-栗疫病菌和SsDRV-核盤菌等互作系統中的分子技術在刺盤孢菌中建立相應的病毒反向遺傳學系統。
3. 2. 4 結合多組學手段研究病毒與寄主真菌互作
弱毒相關病毒與寄主真菌這一特殊系統再結合快速發展的組學研究技術將為研究病毒在寄主中復制并引起癥狀的機理、病原真菌致病力/毒力的構成及調控機理等病毒學和病原真菌學的核心問題提供極大便利。Allen等(2003)應用轉錄組學技術在一定程度上揭示了低毒病毒CHV1-EP713對板栗疫病菌基因轉錄調節的規模和程度:超過2200個差異表達基因,涉及的生物功能包括應激反應、碳代謝和轉錄調節等。Kwon等(2009)利用比較蛋白組學研究弱毒相關病毒Fusarium graminearum virus-DK21(FgV-DK21)的侵染對禾谷鐮刀菌的影響,發現病毒特異性地影響寄主碳代謝、氨基酸代謝及能量代謝等通路中關鍵酶和成分的積累。Lee Marzano等(2018)研究發現,弱毒相關病毒侵染核盤菌后寄主真菌會產生大量特殊的mRNA和sRNA重新編碼產物。因此,利用組學手段可確定真菌病毒侵染對刺盤孢菌轉錄、翻譯和加工的調控模式,確定病毒與刺盤孢菌互作的關鍵基因及刺盤孢菌的致病關鍵基因,為利用真菌病毒防治刺盤孢菌引起的植物炭疽病打下堅實基礎。
參考文獻:
蔡婷. 2015. 核盤菌弱毒菌株8-2-3中病毒分子特性及其對寄主生物學特性的影響[D]. 武漢: 華中農業大學. [Cai T. 2015. Molecular characterization of mycoviruses from Sclerotinia sclerotiorum hypovirulence strain 8-2-3 and their influence on the host[D]. Wuhan: Huazhong Agricultural University.]
成淑芬,李波,謝甲濤,陳桃,程家森,付艷蘋,姜道宏. 2017. 核盤菌低毒相關內源病毒SsEV2的分子特性研究[C]//中國植物病理學會2017年學術年會論文集. 北京:中國農業科學技術出版社. [Cheng S F,Li B,Xie J T,Chen T,Cheng J S,Fu Y P,Jiang D H. 2017. Molecular Cha-racterization of debilitation-associated Sclerotinia sclerotiorum endogenous virus SsEV2[C]//Proceedings of the Aca-demic Annual Conference of Chinese Society of Plant Pathology. Beijing: China Agricultural Science and Techno-logy Press.]
杜磊. 2012. 核盤菌致病力衰退相關病毒SsDRV防治油菜菌核病的研究[D]. 武漢: 華中農業大學. [Du L. 2012. Research on biological control of sclerotinia stem rot of oilseed rape with Sclerotinia sclerotiorum debilitation-associated RNA virus[D]. Wuhan: Huazhong Agricultural University.]
章海龍. 2014. 禾谷鐮刀菌低毒病毒FgHV 2-JS 16的全序列分析與生物學功能研究[D]. 蘭州:甘肅農業大學. [Zhang H L. 2014. Sequence analysis and biological characteristics of hypovirus FgHV 2-JS 16 from Fusarium graminearum[D]. Lanzhou:Gansu Agricultural University.]
周德慶,徐士菊. 2005. 微生物學詞典[M]. 天津:天津科學技術出版社. [Zhou D Q,Xu S J. 2005. Glossary of microbiology[M]. Tianjin: Tianjin Science and Technology Press.]
Allen T D,Dawe A L,Nuss D L. 2003. Use of cDNA microarrays to monitor transcriptional responses of the chestnut blight fungus Cryphonectria parasiticato infection by virulence-attenuating hypoviruses[J]. Eukaryotic Cell,2(6): 1253-1265.
Buck K W. 2017. Fungal virology—An overview[M]//Buck K W. Fungal virology. Boca Raton: CRC Press: 1-84.
Campo S,Gilbert K B,Carrington J C. 2016. Small RNA-based antiviral defense in the phytopathogenic fungus Colleto-trichum higginsianum[J]. PLoS Pathogens,12(6):1-36.
Carbú M,Moraga J,Cantoral J M,Collado I G,Garrido C. 2019. Recent approaches on the genomic analysis of the phytopathogenic fungus Colletotrichum spp.[J]. Phytochemistry Reviews,doi:10.1007/s11101-019-09608-0.
Chen B,Choi G H,Nuss D L. 1994. Attenuation of fungal virulence by synthetic infectious hypovirus transcripts[J]. Science,264(5166): 1762-1764.
Dale J L,Manners J M,Irwin J A G. 1988. Colletotrichum gloeosporioides isolates causing different anthracnose di-seases on Stylosanthes in Australia carry distinct double-stranded RNAs[J]. Transactions of the British Mycological Society,91(4): 671-676.
Figueirêdo L C D,Figueirêdo G S D,Giancoli ? C,Tanaka F A,Silva L A,Kitajima E W,Filho S A,Azevedo J L. 2012. Detection of isometric,dsRNA-containing viral particles in Colletotrichum gloeosporioides isolated from cashew tree[J]. Tropical Plant Pathology,37(2): 142-145.
Grente J. 1965. Les formes hypovirulentes d'Endothia parasitica et les espoirs de lutte contre le chancre du ch?taignier[J]. Comptes Rendus? Acad Agric France,51: 1033-1037.
Heiniger U,Rigling D. 1994. Biological control of chestnut blight in Europe[J]. Annual Review of Phytopathology,32: 581-599.
Hillman B I,Halpern B T,Brown M P. 1994. A viral dsRNA element of the chestnut blight fungus with a distinct genetic organization[J]. Virology,201(2): 241-250.
Howitt R L,Beever R E,Pearson M N,Forster R L. 2001. Genome characterization of Botrytis virus F,a flexuous rod-shaped mycovirus resembling plant ‘potex-like viruses[J]. Journal of General Virology,82(1): 67-78.
Howitt R L,Beever R E,Pearson M N,Forster R L. 2006. Genome characterization of a flexuous rod-shaped mycovirus,Botrytis virus X,reveals high amino acid identity to genes from plant ‘potex-like viruses[J]. Archives of Virology,151(3): 563-579.
Hu Z J,Wu S S,Cheng J S,Fu Y P,Jiang D H,Xie J T. 2014. Molecular characterization of two positive-strand RNA viruses co-infecting a hypovirulent strain of Sclerotinia sclerotiorum[J]. Virology,464: 450-459.
Hutchison E,Brown S,Tian C,Glass N L. 2009. Transcriptional profiling and functional analysis of heterokaryon incompatibility in Neurospora crassa reveals that reactive oxygen species,but not metacaspases,are associated with programmed cell death[J]. Microbiology,155: 3957-3970.
Jayawardena R S,Hyde K D,Damm U,Cai L,Liu M,Li X H,Zhang W,Zhao W S,Yan J Y. 2016. Notes on currently accepted species of Colletotrichum[J]. Mycosphere,7(8): 1192-1260.
Jia H X,Dong K L,Zhou L L,Wang G P,Hong N,Jiang D H,Xu W X. 2017. A dsRNA virus with filamentous viral particles[J]. Nature Communications,8(1): 1-12.
Joshi R. 2018. A review on Colletotrichum spp. virulence mechanism against host plant defensive factors[J]. Journal of Medicinal Plants Studies,6(6): 64-67.
Kenichi I,Kanako I,Chiaki K,Takahiro U,Atsuko S,Satoko K,Pyoyun P. 2013. Potentiation of mycovirus transmission by zinc compounds via attenuation of heterogenic incompatibility in Rosellinia necatrix[J]. Applied and Environmental Microbiology,79(12): 3684-3691.
Khalifa M E,Pearson M N. 2013. Molecular characterization of three mitoviruses co-infecting a hypovirulent isolate of Sclerotinia sclerotiorum fungus[J]. Virology,441(1): 22-30.
Kotta-Loizou I,Coutts R H A. 2017. Mycoviruses in Aspergilli: A comprehensive review[J]. Frontiers in Microbiology,8: 1-15.
Kwon S J,Cho S Y,Lee K M,Yu J,Son M,Kim K H. 2009. Proteomic analysis of fungal host factors differentially expressed by Fusarium graminearum infected with Fusa-rium graminearum virus-DK21[J]. Virus Research,144(1-2): 96-106.
Lecoq H,Boissonnetmenès M,Delhotal P. 1979. Infectivity and transmission of fungal viruses[M]. Berlin: Springer: 34-47.
Lee Marzano S Y,Neupane A,Domier L. 2018. Transcriptio-nal and small RNA responses of the white mold fungus Sclerotinia sclerotiorum to infection by a virulence-atte-nuating hypovirus[J]. Viruses,10(12): 1-25.
Li P F,Zhang H L,Chen X G,Qiu D W,Guo L H. 2015. Molecular characterization of a novel hypovirus from the plant pathogenic fungus Fusarium graminearum[J]. Viro-logy,481: 151-160.
Liao C Y,Chen M Y,Chen Y K,Wang T C,Sheu Z M,Kuo K C,Chang P F L,Chung K R,Lee M H. 2012. Characterization of three Colletotrichum acutatum isolates from Capsicum spp.[J]. European Journal of Plant Pathology,133(3): 599-608.
Lima J S,Figueiredo J G,Gomes R G,Stringari D,Goulin E H,Adamoski D,Kava-Cordeiro V,Galli-Terasawa L V,Glienke C. 2012. Genetic diversity of Colletotrichum spp. an endophytic fungi in a medicinal plant,Brazilian pepper tree[J]. International Scholarly Research Notices: Microbiology,2012: 1-7.
Linder-Basso D,Dynek J N,Hillman B I. 2005. Genome ana-lysis of Cryphonectria hypovirus 4,the most common hypovirus species in North America[J]. Virology,337(1): 192-203.
Liu S,Xie J T,Cheng J S,Li B,Chen T,Fu Y P,Li G Q,Wang M Q,Jin H N,Wan H,Jiang D H. 2016. Fungal DNA virus infects a mycophagous insect and utilizes it as a transmission vector[J]. Proceedings of the National Academy of Sciences of the United States of America,113(45): 12803-12808.
MacDonald W L,Cech F C,Luchok J,Smith C. 1978. Proceedings of the American chestnut symposium[M]. West Virginia: West Virginia University:1-122.
Marzano S L,Nelson B D,Ajayi-Oyetunde O,Bradley C A,Hughes T J,Hartman G L,Eastburn D M,Domier L L. 2016. Identification of diverse mycoviruses through metatranscriptomics characterization of the viromes of five major fungal plant pathogens[J]. Journal of Virology,90(15): 6846-6863.
Moffitt E M,Lister R M. 1975. Application of a serological screening test for detecting double-stranded RNA mycoviruses[J]. Phytopathology,65: 581-589.
Money N P. 1999. Fungus punches its way in[J]. Nature,401: 332-333.
Mu F,Xie J T,Cheng S F,You M P,Barbetti M J,Jia J C,Wang Q Q,Cheng J S,Fu Y P,Chen T,Jiang D H. 2017. Virome characterization of a collection of Sclerotinia sclerotiorum from Australia[J]. Frontiers in Microbiology,8: 1-18.
Nuss D L. 2005. Hypovirulence: Mycoviruses at the fungal-plant interface[J]. Nature Reviews: Microbiology,3(8): 632-642.
Nuss D L. 2011. Mycoviruses,RNA silencing,and viral RNA recombination[J]. Advances in Virus Research,80: 25-48.
Rawlinson C J,Carpenter J M,Muthyalu G. 1975. Double-stranded RNA virus in Colletotrichum lindemuthianum[J]. Transactions of the British Mycological Society,65(2): 305-308.
Rosseto P,Costa A T,Polonio J C,da Silva A A,Pamphile J A,Azevedo J L. 2016. Investigation of mycoviruses in endophytic and phytopathogenic strains of Colletotrichum from different hosts[J]. Genetics and Molecular Research,doi:10.4238/gmr.15017651.
Sasaki A,Onoue M,Kanematsu S,Suzaki K,Miyanishi M,Suzuki N,Nuss D L,Yoshida K. 2002. Extending chestnut blight hypovirus host range within diaporthales by bio-listic delivery of viral cDNA[J]. Molecular Plant-Microbe Interactions,15 (8): 780.
Shi M,Lin X D,Tian J H,Chen L J,Chen X,Li C X,Qin X C,Li J,Cao J P,Eden J S,Buchmann J,Wang W,Xu J,Holmes E C,Zhang Y Z. 2016. Redefining the invertebrate RNA virosphere[J]. Nature,540(7634): 539-543.
Stauder C M,Nuss D L,Zhang D X,Double M L,MacDonald W L,Metheny A M,Kasson M T. 2018. Enhanced hypovirus transmission by engineered super donor strains of the chestnut blight fungus,Cryphonectria parasitica,into a natural population of strains exhibiting diverse vegetative compatibility genotypes[J]. Virology,528: 1-6.
Varga J,Kevei F,Vágv?lgyi C,Vriesema A,Croft J. 1994. Double-stranded RNA mycoviruses in section Nigri of the Aspergillus genus[J]. Canadian Journal of Microbio-logy,40(4): 325-329.
Varga J,Vágv?lgyi C,Tóth B. 2003. Recent advances in mycovirus research[J]. Acta Microbiologica et Immunologica Hungarica,50(1): 77-94.
Wang L P,Jiang J J,Wang Y F,Hong N,Zhang F P,Xu W X,Wang G P. 2014. Hypovirulence of the phytopathogenic fungus Botryosphaeria dothidea: Association with a coinfecting chrysovirus and a partitivirus[J]. Journal of Viro-logy,88(13): 7517-7527.
Wang S C,Kondo H,Liu L,Guo L H,Qiu D W. 2013. A no-vel virus in the family Hypoviridae from the plant pathogenic fungus Fusarium graminearum[J]. Virus Research,174(1-2): 69-77.
Willis K J,Bachman S. 2016. State of the worlds plants[R]. London:Royal Botanic Gardens.
Wu M D,Jin F Y,Zhang J,Yang L,Jiang D H,Li G Q. 2012. Characterization of a novel bipartite double-stranded RNA mycovirus conferring hypovirulence in the phytopathogenic fungus Botrytis porri[J]. Journal of Virology,86(12): 6605-6619.
Wu M D,Zhang L,Li G Q,Jiang D H,Ghabrial S A. 2010. Genome characterization of a debilitation-associated mitovirus infecting the phytopathogenic fungus Botrytis cinerea[J]. Virology,406(1): 117-126.
Wu M D,Zhang L,Li G Q,Jiang D H,Hou M S,Huang H C. 2007. Hypovirulence and double-stranded RNA in Botrytis cinerea[J]. Phytopathology,97(12): 1590-1599.
Wu S S,Cheng J S,Fu Y P,Chen T,Jiang D H,Ghabrial S A,Xie J T. 2017. Virus-mediated suppression of host non-self recognition facilitates horizontal transmission of heterologous viruses[J]. PLoS Pathogens,13(3): 1-25.
Xiao X Q,Cheng J S,Tang J H,Fu Y P,Jiang D H,Baker T S,Ghabrial S A,Xie J T. 2014. A novel partitivirus that confers hypovirulence on plant pathogenic fungi[J]. Journal of Virology,88(17): 10120-10133.
Xie J,Ghabrial S A. 2012. Molecular characterization of two mitoviruses co-infecting a hypovirulent isolate of the plant pathogenic fungus Sclerotinia sclerotiorum[J]. Virology,428(2): 77-85.
Xie J,Wei D,Jiang D,Fu Y,Li G,Ghabrial S,Peng Y. 2006. Characterization of debilitation-associated mycovirus infecting the plant-pathogenic fungus Sclerotinia sclerotiorum[J]. Journal of General Virology,87(1): 241-249.
Xie J T,Jiang D H. 2014. New insights into mycoviruses and exploration for the biological control of crop fungal di-seases[J]. Annual Review of Phytopathology,52: 45-68.
Xie J T,Xiao X Q,Fu Y P,Liu H Q,Cheng J S,Ghabrial S A,Li G Q,Jiang D H. 2011. A novel mycovirus closely related to hypoviruses that infects the plant pathogenic fungus Sclerotinia sclerotiorum[J]. Virology,418(1): 49-56.
Yan Y Q,Yuan Q F,Tang J T,Huang J B,Hsiang T,Wei Y D,Zheng L. 2018. Colletotrichum higginsianum as a model for understanding host-pathogen interactions: A review[J]. International Journal of Molecular Sciences,19(7): 1-18.
Yu X,Li B,Fu Y,Jiang D,Ghabrial S A,Li G,Peng Y,Xie J,Cheng J,Huang J,Yi X. 2010. A geminivirus-related DNA mycovirus that confers hypovirulence to a plant pathogenic fungus[J]. Proceedings of the National Aca-demy of Sciences of the United States of America,107(18): 8387-8392.
Yu X,Li B,Fu Y,Xie J,Cheng J,Ghabrial S A,Li G,Yi X,Jiang D. 2013. Extracellular transmission of a DNA mycovirus and its use as a natural fungicide[J]. Proceedings of the National Academy of Sciences of the United States of America,110(4): 1452-1457.
Zhai L F,Zhang M X,Hong N,Xiao F,Fu M,Xiang J,Wang G P. 2018. Identification and characterization of a novel hepta-segmented dsRNA virus from the phytopathogenic fungus Colletotrichum fructicola[J]. Frontiers in Microbio-logy,9 (754): 1-13.
Zhang D X,Nuss D L. 2016. Engineering super mycovirus donor strains of chestnut blight fungus by systematic disruption of multilocus vic genes[J]. Proceedings of the National Academy of Sciences of the United States of Ame-rica,113(8): 2062-2067.
Zheng L,Shu C,Zhang M L,Yang M,Zhou E. 2019. Molecular characterization of a novel endornavirus conferring hypovirulence in rice sheath blight fungus Rhizoctonia solani AG-1 IA strain GD-2[J]. Viruses,11(2): 1-14.
Zheng L,Zhang M L,Chen Q G,Zhu M H,Zhou E. 2014. A novel mycovirus closely related to viruses in the genus Alphapartitivirus confers hypovirulence in the phytopathogenic fungus Rhizoctonia solani[J]. Virology,456-457: 220-226.
Zhong J,Chen D,Lei X H,Zhu H J,Zhu J Z,Gao B D. 2014. Detection and characterization of a novel Gammapartitivirus in the phytopathogenic fungus Colletotrichum acutatum strain HNZJ001[J]. Virus Research,190: 104-109.
Zhong J,Pang X D,Zhu H J,Gao B D,Huang W K,Zhou Q. 2016. Molecular characterization of a trisegmented mycovirus from the plant pathogenic fungus Colletotrichum gloeosporioides[J]. Viruses,8(10): 1-12.
(責任編輯 麻小燕)
