廣西潿洲島珊瑚礁海域魚類物種組成的調查分析
2020-03-20 03:44:50鄒琦吳志強黃亮亮丁洋黃欣
南方農業學報 2020年1期
鄒琦 吳志強 黃亮亮 丁洋 黃欣
鄒琦(1983-),江西贛州人,理學博士,畢業于桂林理工大學環境科學與工程專業,愛爾蘭科克大學(University College Cork)訪問學者,主要從事魚類生態學、海洋生物學及生物多樣性分析等研究工作。參與完成國家自然科學基金項目“廣西北部灣沿岸紅樹林區域仔稚魚的群落結構及其多樣性研究”“廣西潿洲島珊瑚礁魚類物種多樣性研究”,廣西自然科學基金項目“會仙濕地魚類群聚時空變化格局及其多樣性維持機制”等多項科研項目。現主持參與國家自然科學基金項目、廣西自然科學基金項目、江西省自然科學基金項目等國家級、省部級項目5項。在《PLoS One》《Journal of Applied Ichthyology》《Chemosphere》《南方農業學報》等國內外期刊上發表科技論文20余篇,其中,被SCI、EI收錄10余篇。作為主要成員參與出版專著和教材4部。
摘要:【目的】明確廣西潿洲島珊瑚礁海域魚類物種組成及其優勢物種,為廣西潿洲島漁業資源的合理開發及科學管理提供科學依據。【方法】于2015年10月—2016年9月,采用三重刺網結合潛水和垂釣的方法逐月對廣西潿洲島珊瑚礁海域5個采樣點進行魚類采樣調查,通過相對重要性指數(IRI)確定優勢物種,并采用相似度指數(Sc)、非加權平均聚類法(UPGMA)及無度量多維排序(NMDS)比較分析不同珊瑚礁海域的魚類物種組成結構。【結果】從廣西潿洲島珊瑚礁海域共采集魚類標本2051尾,經鑒定隸屬于2綱12目49科84屬114種,其中,軟骨魚綱有2目2科2屬2種,硬骨魚綱有10目47科112種。硬骨魚綱中以鱸形目種類最多(85種),占采集物種總數的74.56%。廣西潿洲島珊瑚礁海域四季的魚類物種數存在一定差異,春季有88種、夏季有63種、秋季有76種、冬季有50種;從魚類棲息水層來看,底層魚類有53種,中下層魚類有45種,中上層魚類有16種,分別占采集物種總數的46.49%、39.47%和14.04%;該海域魚類以暖水性魚類為主(102種),暖溫性魚類僅12種,未見冷溫性和溫水性魚類。廣西潿洲島珊瑚礁海域魚類物種組成與同緯度徐聞珊瑚礁海域的相似性最高,與海南島珊瑚礁海域的相似性相對較低,而與西沙趙述島珊瑚礁海域的相似性最低。【結論】廣西潿洲島珊瑚礁海域魚類以暖水性為主,四季魚類物種組成存在一定差異,以底層和中下層魚類為主,但魚類物種相對豐度較低且數量相對偏少,與受人類活動干擾有一定關系。因此,為更好地保護廣西潿洲島珊瑚礁海域魚類物種多樣性,建議有關部門采取相應措施減緩過度捕撈或旅游區開發等人類活動的影響。
關鍵詞: 珊瑚礁;物種組成;魚類多樣性;廣西潿洲島
中圖分類號: S932.4? ? ? ? ? ? ? ? ? ? ? ? ?文獻標志碼: A 文章編號:2095-1191(2020)01-0001-10
Abstract:【Objective】It was to clarify the composition of the fish species as well as dominant species in coral reef area of Weizhou Island in Guangxi, and provide scientific basis for the reasonable development and scientific management of fishery resources in Weizhou Island of Guangxi. 【Method】The fish samples were collected monthly from five sampling sites by multi-approaches(such as trammel nets, diving and fishing) from October 2015 to September 2016 in Weizhou Island. Through the index of relative importance(IRI), the dominant species were determined. Moreover, the species compositions from different coral reefs were compared and analyzed by the similarity index(Sc), unweighted pair group me-thod with arithmetic mean(UPGMA) and nonmetric multidimensional scaling(NMDS). 【Result】The results showed that a total of 2051 fish individuals were collected, which were identified to 114 species, belonging to 2 class, 12 orders, 49 families, 84 genera. There were two species of Chondrichthyes, belonging to two orders, two families and two genera. There were 112 species of Osteichthyes belonging to 10 orders, 47 families. Perciformes was the richest fish families (85 fish species) among the Osteichthyes, accounting for 74.56% of the total fish species. The species numbers in coral reef area of Weizhou Island in the four seasons were different, there were 88 species in spring, 63 in summer, 76 in autumn and 50 in winter. In terms of the perched water layers, there were 53 demersal fish species, 45 benthopelagic fish species, and 16 pelagic fish species, accounting for 46.49%, 39.47% and 14.04% of the total species, respectively. The warm-water fish was the dominant fish, comprising 102 species, and 12 warm-temperate fish species were collected in this area. There was no cold-temperature species or temperate water species. The fish species compositions in coral reef area of Weizhou Island had the highest similarity with coral reef area of Xuwen, where the altitude was the same. The similarity with Hainna Island was low, and the similarity with Zhaoshu Island of Xisha Islands was the lowest. 【Conclusion】The dominant fish species in coral reef areas in Weizhou Island of Guangxi are warm-water fishes, the compositions vary in the four seasons.? demersal and benthopelagic fish species are the majority, but the species relative abundance is low and species number is small, which may be related to disturbance of human activities.
Key words: coral reef; species composition; fish diversity; Weizhou Island, Guangxi
Foundation item: National Natural Science Foundation of China(41566003); Guangxi Natural Science Foundation(2018GXNSFAA281022); Guangxi Bagui Scholars Special Project(2016A10)
0 引言
【研究意義】隨著人類活動不斷向海洋延伸,珊瑚礁生態系統退化的問題日益凸顯(Polmear et al.,2015),因過度捕撈、旅游開發及污染物排放等人為因素致使全球四分之三的珊瑚礁已受到影響(Burke et al.,2011),其中又以生物生存受到的威脅最嚴重(Heron et al.,2016;Zaneveld et al.,2016;Webster et al.,2018)。我國98%的珊瑚礁分布在南海,珊瑚礁生態系統功能退化也非常嚴重,如海南島沿岸珊瑚覆蓋率在過去30年中下降了80%(Hughes et al.,2013)。珊瑚礁魚類是診斷或評價修復退化珊瑚礁生態系統的關鍵組成部分(Spalding et al.,2001;MacNeil et al.,2015),因此,調查研究珊瑚礁魚類有助于了解珊瑚礁生態系統的演變過程,為開展海洋生態系統保護工作提供科學依據。【前人研究進展】目前,世界珊瑚礁魚類研究主要集中于氣候變化和人類活動影響下魚類個體、種群及群落等層面的響應特征(Chen et al.,2013),且主要針對大面積珊瑚礁集聚區,如加勒比海礁區(Hawkins and Ro-berts,1996)、東亞珊瑚礁區(Shi et al.,2003)、澳大利亞大堡礁(Abaya et al.,2018)等。Ottimofiore等(2017)研究發現,珊瑚礁及其相關生物群落在很大程度上受持續氣候變化的影響,且該結論可用于預測珊瑚礁魚類今后對氣候變化的響應情況。Pinheiro等(2017)以珊瑚礁魚類為例,構建了海平面變化與地理隔離對海洋生物多樣性影響的理論模型,并證實人類活動和氣候變化共同作用于魚類群落結構。Rodgers等(2018)研究表明,持續的海洋變暖會降低低緯度珊瑚礁魚類的生存能力。我國針對珊瑚礁魚類的研究起步較晚,其研究區域主要在南海諸島和海南島等水域。孫典榮等(2005,2006)先后對西沙和南沙群島珊瑚礁魚類資源的物種組成和區系及數量分布進行調查分析,有效填補了我國南海珊瑚礁魚類資源調查的空白。此外,孫典榮等(2005,2006)、史赟榮等(2009)、李永振等(2011)研究表明,南海珊瑚礁魚類物種多樣性大尺度分布格局的主要特征是物種多樣性較高,但分類多樣性較低,具體表現為南海西沙和中沙群島的珊瑚礁海域魚類有632種,東沙群島的珊瑚礁海域魚類有514種,南沙群島的珊瑚礁海域魚類有548種。【本研究切入點】廣西潿洲島珊瑚礁地處熱帶珊瑚分布的北緣,地域特異性使其具有極高的生態價值,但隨著廣西北部灣經濟區建設的推進,以及旅游資源和海洋資源開發等人類活動的加劇,勢必導致該海域魚類多樣性及其物種資源的下降。目前,針對廣西潿洲島珊瑚礁水域的研究主要聚焦于珊瑚物種組成及資源調查(陳剛等,2016),而有關該海域的魚類資源調查尚無研究報道。【擬解決的關鍵問題】2015年10月—2016年9月對廣西潿洲島珊瑚礁海域進行逐月采樣調查,研究該海域魚類物種組成,并明晰其魚類生態類型及主要優勢物種,為廣西潿洲島漁業資源的合理開發及科學管理提供科學依據。
1 材料與方法
1. 1 研究區域概況
廣西潿洲島位于我國南海北部灣東北部(東經109°00′~109°15′,北緯21°00′~21°10′),面積約26 km2,是廣西最大的海島;屬南亞熱帶季風氣候區,年均氣溫23.0 ℃,平均降水量1297 mm,年均海面溫度24.6 ℃,年均海水鹽度31.9‰(黎廣釗等,2004)。廣西潿洲島周圍沿岸的北面、東面、西南面淺海均分布有珊瑚礁,珊瑚礁總覆蓋率為17.5%~19.5%,構成廣西沿海唯一的珊瑚礁群,具有極高的資源觀賞和科研價值。
1. 2 研究方法
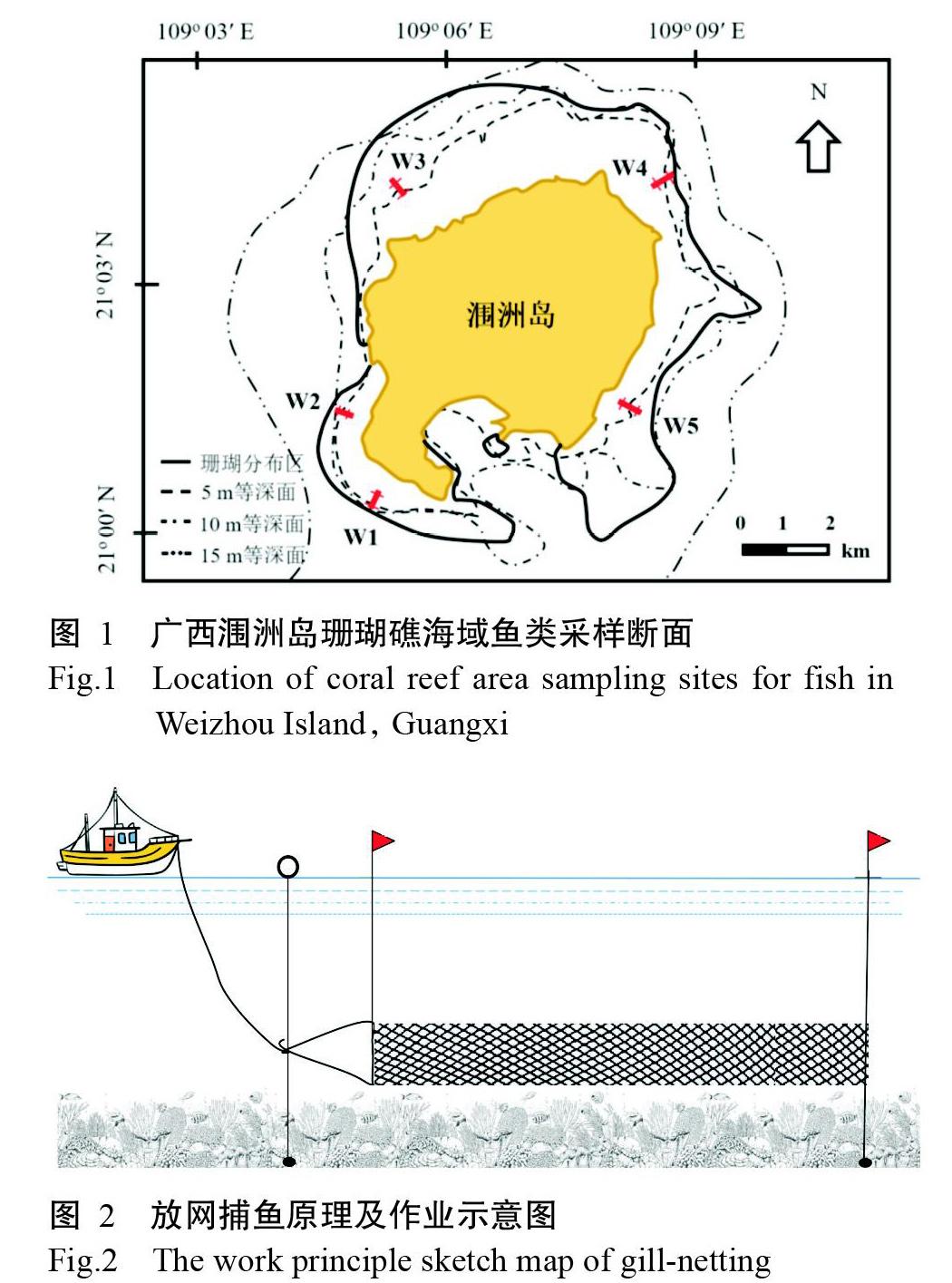
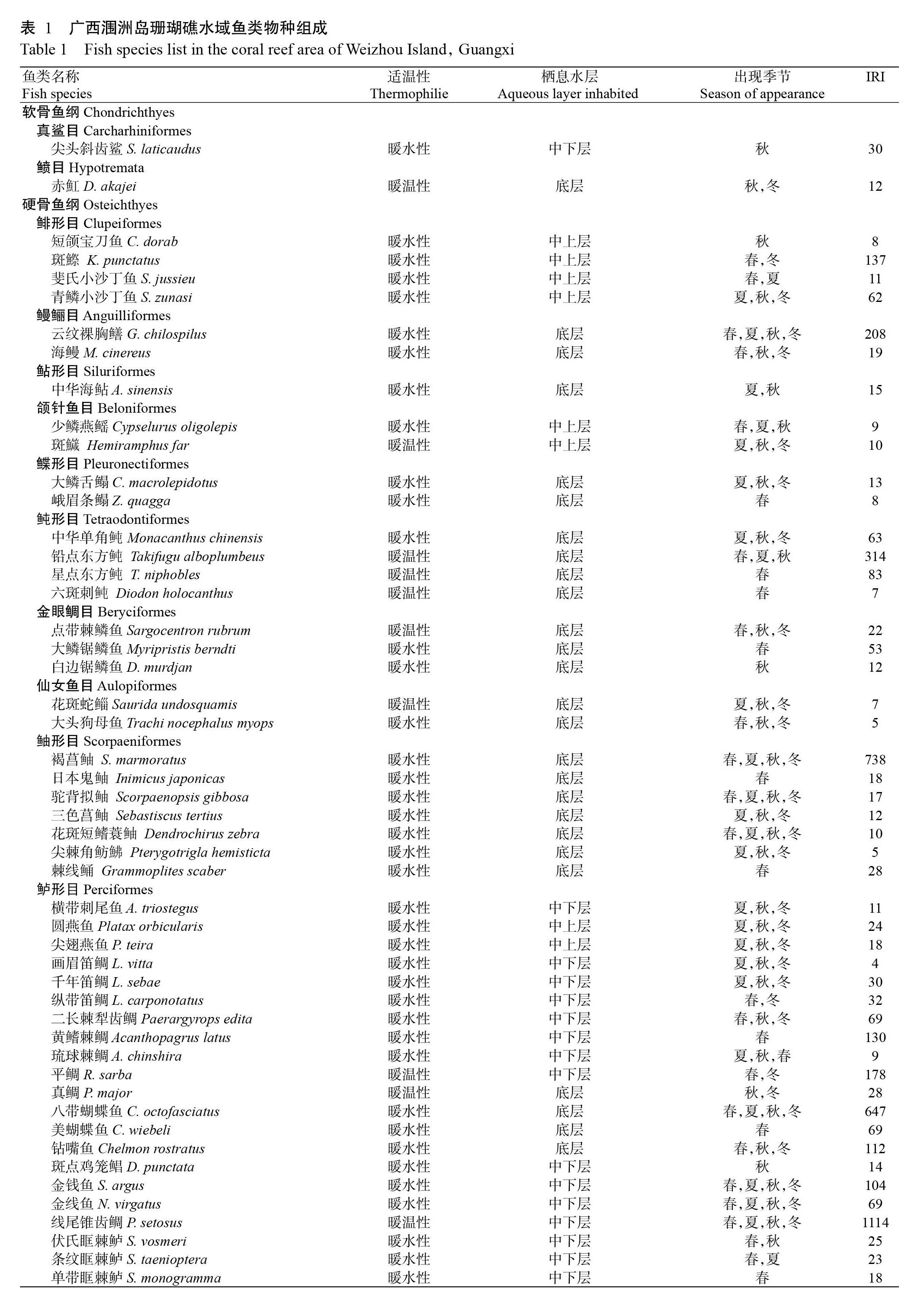
根據廣西潿洲島海域珊瑚礁分布區域及歷史調查資料,在潿洲島北部、東部和西南海岸淺水區以垂直海岸方向布設5條主剖面(圖1)。2015年10月—2016年9月逐月對該海域的5個采樣點進行采樣,包含冬季(12月—翌年2月)、春季(3—5月)、夏季(6—8月)和秋季(9—11月)4個季節。采樣網具為三重刺網,網長50 m,外網目尺寸10 cm,內網目尺寸6 cm(圖2)。放網時將多張網衣連在一起作為一組網進行采樣,放網時間一般為下午16:00—次日上午6:00;并以潛水和垂釣等方法進行輔助調查。所得漁獲物均進行分類、拍照及鑒定(李德越等,2018),其中,魚類鑒定主要參考《臺灣魚類志》(沈世杰,1993)、《中國南海西南中沙群島珊瑚礁魚類圖譜》(傅亮,2014)、《廣西北部灣海洋硬骨魚類圖鑒》(賴廷和和何斌源,2016)、《北部灣魚類圖鑒》(劉靜等,2016)及世界魚類數據庫(Fish Base)和臺灣魚類資料庫(The Fish Data Base of Taiwan)等。
1. 3 統計分析
為研究廣西潿洲島珊瑚礁海域魚類在屬、科及目等分類階元的豐度,利用相對豐度指數(R)表征不同屬、目、科魚類所占比例(Robertson,1998;Bonnici et al.,2018)。
式中,N為某種魚類個體數占魚類個體總數的百分比(%),W為某種魚類生物量占魚類總生物量的百分比(%),F為出現頻率。同時,參照陳國寶等(2006)的研究分類方法對珊瑚礁魚類優勢種進行區分:IRI≥500為優勢種,100≤IRI<500為常見種,10≤IRI<100為一般種,IRI<10為少見種。
為分析廣西潿洲島珊瑚礁海域魚類與我國其他珊瑚礁海域魚類的相似性,利用相似性指數(Sc)表征各海域間的分類相似性,參照張鐿鋰和張雪梅(1998)的方法進行計算:
式中,A和B分別為兩個海域記錄的魚類物種數,C為兩海域共有魚類物種數。Sc還可用于表征其他同級階元(科、目)的相似度(吳鐘解等,2015)。此外,采用非加權平均聚類法(UPGMA)對各珊瑚礁海域魚類組成進行聚類分析(Huang et al.,2016),并結合無度量多維排序(Non-metric multidimensional scaling, NMDS)分析珊瑚礁魚類群落的區系特征。
2 結果與分析
2. 1 魚類物種組成
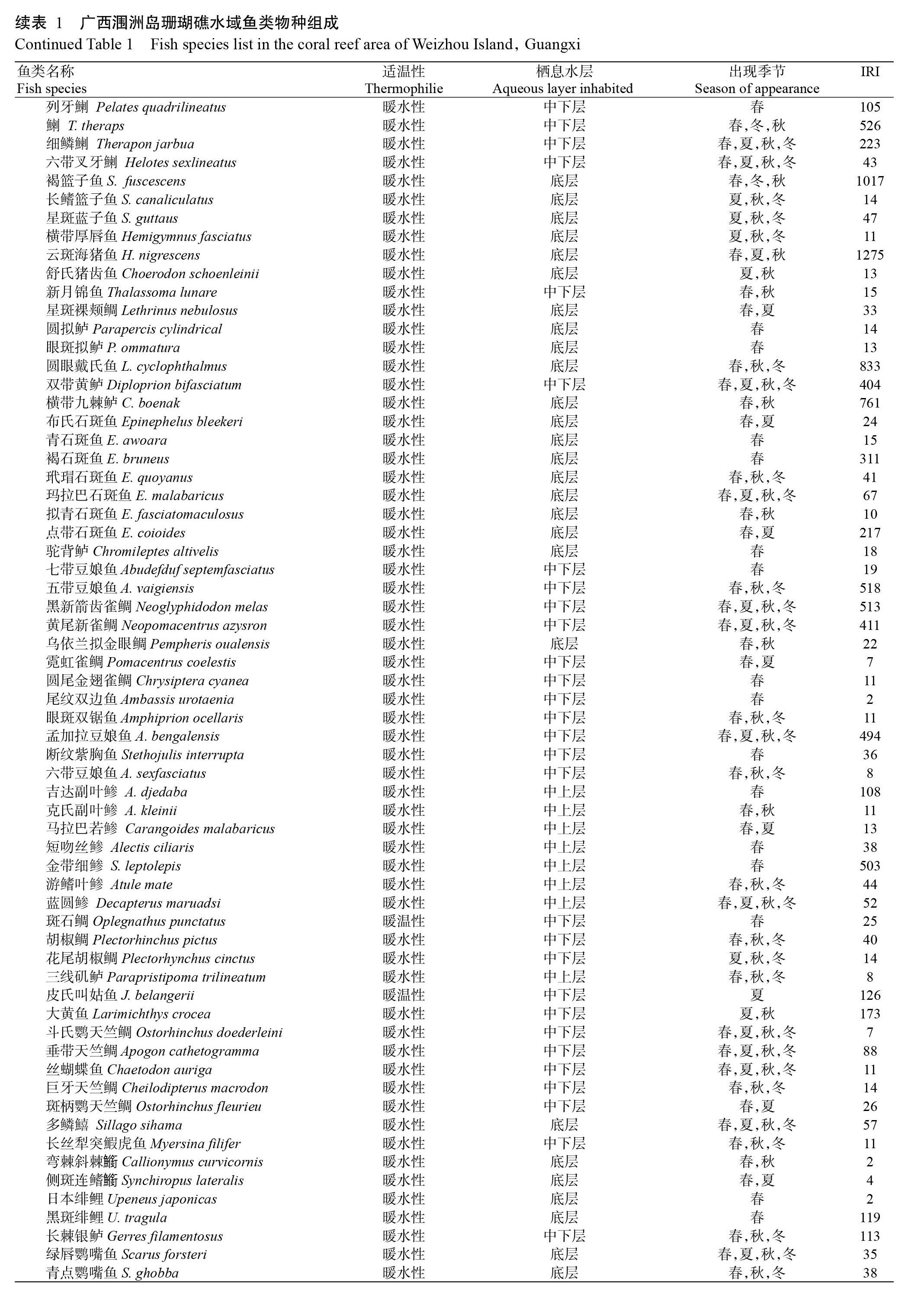
從廣西潿洲島珊瑚礁海域共采集魚類標本2051尾,經鑒定隸屬于2綱12目49科84屬114種(表1)。其中,軟骨魚綱有2目2科2屬2種,硬骨魚綱有10目47科82屬112種。鱸形目有29科59屬85種,占采集物種總數的74.56%;鲉形目有4科6屬7種,占采集物種總數的6.14%;鯡形目有2科3屬4種,占采集物種總數的3.51%;鲀形目有3科3屬4種,占采集物種總數的3.51%;金眼鯛目有1科2屬3種,占采集物種總數的2.63%;鰈形目、頜針魚目、鰻鱺目和仙女魚目各有2科2屬2種,均占采集物種總數的1.75%;鲇形目有1科1屬1種,占采集物種總數的0.88%。
在科水平上(表2),鮨科有5屬10種,占采集物種總數的8.77%;雀鯛科有9種、鲹科有7種,分別占采集物種總數的7.89%和6.14%;鯛科、金線魚科和隆頭魚科均為5種,分別占采集物種總數的4.39%;鲉科、天竺鯛科、鯻科和蝴蝶魚科均為4種,分別占采集物種總數的3.51%;笛鯛科、鯡科、金鱗魚科、籃子魚科和石鱸科均3種,分別占采集物種總數的2.63%;鸚嘴魚科、白鯧科、狗母魚科、擬鱸科、鲀科、羊魚科、石首魚科和?科各有2種,分別占采集物種總數的1.75%;鲅科、寶刀魚科、刺鲀科、毒鲉科、單棘鲀科、魴鮄科、飛魚科、海鰻科、海鲇科、魟科、雞籠鯧科、金錢魚科、裸頰鯛科、擬金眼鯛科、擬雀鯛科、牛尾魚科、海鱔科、舌鰨科、石鯛科、雙邊魚科、鰨科、鱚科、鰕虎魚科、銀鱸科、真鯊科和鱵科各1種,合計占采集物種總數的22.81%。
廣西潿洲島珊瑚礁海域四季的魚類物種數存在一定差異,春季有88種、夏季有63種、秋季有76種、冬季有50種。其中,有21種魚類在四季均出現,而25種魚類僅在春季出現,4種魚類[尖頭斜齒鯊(Sco-liodon laticaudus)、白邊鋸鱗魚(Dispinus murdjan)、短頜寶刀魚(Chirocentrus dorab)和斑點雞籠鯧(Drepane punctata)]僅在秋季出現,而皮氏叫姑魚(Johnius belangerii)僅在夏季出現(表1)。由表1還可看出,云斑海豬魚(Halichoeres nigrescens)、線尾錐齒鯛(Pentapodus setosus)、褐籃子魚(Siganus fuscescens)、圓眼戴氏魚(Labracinus cyclophthalmus)、橫帶九棘鱸(Cephalopholis boenak)、褐菖鲉(Sebastiscus marmoratus)、八帶蝴蝶魚(Chaetodon octofasciatus)、鯻(Terapon theraps)、五帶豆娘魚(Abudefduf vaigiensis)、黑新箭齒雀鯛(Neoglyphidodon melas)和金帶細鲹(Selaroides leptolepis)11個魚類物種的IRI>500,為優勢物種。
2. 2 魚類生態特征
廣西潿洲島珊瑚礁海域魚類主要由底層魚類、中下層魚類和中上層魚類組成(表1)。其中,底層魚類主要有赤魟(Dasytis akajei)、云紋裸胸鱔(Gymnothorax chilospilus)、海鰻(Muraenesox cinereus)、中華海鲇(Arius sinensis)、大鱗舌鰨(Cynoglossus macrolepidotus)和峨眉條鰨(Zebrias quagga)等53種,占采集物種總數的46.49%;中下層魚類主要有尖頭斜齒鯊(Scoliodon laticaudus)、橫帶刺尾魚(Acanthurus triostegus)、畫眉笛鯛(Lutjanus vitta)、金錢魚(Scatophagus argus)、金線魚(Nemipterus virgatus)和伏氏眶棘鱸(Scolopsis vosmeri)等45種,占采集物種總數的39.47%;中上層魚類主要有短頜寶刀魚(Chirocentrus dorab)、斑鰶(Konosirus punctatus)、斐氏小沙丁魚(Sardinella jussieu)、斑鱵(Hemiramphus far)、圓燕魚(Platax orbicularis)和吉達副葉鲹(Alepes djedaba)等16種,占采集物種總數的14.04%。
魚類按其生存水體溫度變化幅度可分為冷溫性、溫水性、暖溫性和暖水性四大類,對應的適宜溫度范圍分別為5~10 ℃、10~20 ℃、15~25 ℃和20~30 ℃。廣西潿洲島地處熱帶北緣區域,其珊瑚礁海域出現的魚類以暖水性魚類為主(表1),除赤魟、斑鱵、鉛點東方鲀(Takifugu alboplumbeus)、星點東方鲀(T. Niphobles)、六斑刺鲀(Diodon holocanthus)、點帶棘鱗魚(Sargocentron rubrum)、花斑蛇鯔(Saurida undosquamis)、平鯛(Rhabdosargus sarba)、真鯛(Pagrus major)、線尾錐齒鯛、斑石鯛(Oplegnathus punctatus)和皮氏叫姑魚等12種為暖溫性魚類外,其余的102種均為暖水性魚類,占采集物種總數的89.47%;未見冷溫性和溫水性魚類。
2. 3 魚類物種相似性
廣西潿洲島珊瑚礁是典型的熱帶北緣珊瑚礁生態系統,因此選擇不同緯度的珊瑚礁海域(圖3)魚類物種組成與其進行相似性指數(Sc)對比分析,包括徐聞珊瑚礁海域(楊國歡等,2008)、海南島珊瑚礁海域(吳鐘解等,2015)和西沙趙述島珊瑚礁海域(楊位迪等,2018)。結果顯示,廣西潿洲島與徐聞均屬于北部灣的海域,二者的珊瑚礁魚類物種相似度較高,共有魚類有35種;其中,單棘鲀科、海鲇科、雞籠鯧科、金錢魚科和海鱔科的種類Sc均為100%,蝴蝶魚科、海鰻科和鯻科的種類Sc分別為50%、50%和75%(表2)。廣西潿洲島與相對較遠的海南島,其魚類物種相似度較低,共有魚類有27種;在49個科中只有銀鱸科、刺鲀科和擬鱸科的種類Sc達50%,其余各科的種類Sc均較低。廣西潿洲島與西沙趙述島的珊瑚礁魚類物種相似度最低,共有魚類僅7種;雖然二者的蝴蝶魚科、鸚嘴魚科、隆頭魚科、擬鱸科、雀鯛科和石鱸科具有一定相似度,但其種類Sc均未超過50%。
UPGMA聚類分析結果和NMDS排序結果均表明,廣西潿洲島珊瑚礁海域的魚類物種組成與同緯度徐聞珊瑚礁海域的相似性最高,與海南島珊瑚礁海域的相似性相對較低,而與西沙趙述島珊瑚礁海域的相似性最低(圖4)。
3 討論
珊瑚礁海域分布的珊瑚物種數目(x)與魚類物種數目(y)存在密切關系(y=-13.63+3.92x)(?hman et al.,1997)。廣西潿洲島珊瑚礁海域記錄的石珊瑚和柳珊瑚共有55種(王文歡,2017),故可推算廣西潿洲島珊瑚礁海域的魚類實際數目應在200種以上。本研究在廣西潿洲島珊瑚礁海域共發現魚類物種有114種,相對于國內其他低緯度珊瑚礁海域,其魚類物種數量偏少,究其原因主要是:一方面,海洋生物物種數以印尼—菲律賓海域為中心,隨緯度的升高而梯度減少(Bellwood and Hughes,2001)。廣西潿洲島珊瑚礁地處熱帶北緣,故珊瑚礁魚類數目相對較少。另一方面,我國西沙、南沙等珊瑚礁海域尚屬于人類活動較少的區域,而廣西潿洲島海域受人類活動干擾較嚴重,包括過度捕撈、船舶航運、生活污水排放、水產養殖及旅游區建設等(史海燕,2012)。
廣西潿洲島珊瑚礁海域的魚類以鱸形目為主,鱸形目魚類占采集物種總數的74.56%,與南海其他珊瑚礁海域的魚類目級組成(李永振,2010)一致。在魚類科級組成方面,雀鯛科有9種、鲹科有7種,分別占采集物種總數的7.89%和6.14%;其次是鯛科、金線魚科和隆頭魚科均為5種,分別占采集物種總數的4.39%;鲉科、天竺鯛科、鯻科和蝴蝶魚科均為4種,分別占采集物種總數的3.51%。上述科目魚類具有較高的經濟和觀賞價值,其中蝴蝶魚科魚類還是珊瑚礁生態系統健康水平重要的指示種(Kulbicki et al.,2005)。本研究的調查結果顯示,廣西潿洲島珊瑚礁海域的八帶蝴蝶魚為優勢種,其IRI為647,但蝴蝶魚科魚類僅有4種,從魚類物種組成上進一步驗證周浩郎和黎廣釗(2014)研究得出的觀點,即廣西潿洲島珊瑚礁已處于衰退過程中的亞健康狀態。而從魚類棲息水層來看,廣西潿洲島珊瑚礁水域的底層魚類種數最多,其次是近底層(中下層)魚類,中上層魚類相對較少,與中沙、西沙和南沙群島等珊瑚礁海域的研究結果(孫典榮等,2006;李永振,2010)具有一致性,可能是珊瑚礁海域底層生境復雜且營養豐富的特點更適合底棲魚類生存(楊國歡等,2008)。此外,廣西潿洲島珊瑚礁海域的魚類以暖水性為主,四季魚類物種數存在一定差異,與廣西潿洲島地處亞熱帶北緣,氣候屬南亞熱帶季風氣候區有關,其年均海面溫度為24.6 ℃。Richards等(2012)研究認為,海水溫度變化對珊瑚礁魚類的活動與生長發育均有一定影響。因此,在耐受溫度范圍內,廣西潿洲島珊瑚礁海域魚類數目總體上隨溫度的升高而增加,隨溫度的降低而減少。
廣西潿洲島和徐聞位于我國大陸南部沿岸淺海(圖3),緯度位置較接近,均屬于熱帶北緣氣候,且兩地受同一洋流影響,故魚類物種組成相似性最高;海南島珊瑚礁海域和西沙趙述島珊瑚礁海域所處的緯度相對較低,其氣候變化及南海洋流等海洋環境因素與廣西潿洲島珊瑚礁海域不同(Chen et al.,2004)。受氣候帶和洋流等因素的影響,廣西潿洲島珊瑚礁海域與海南島珊瑚礁海域的魚類相似性相對較低,與西沙趙述島珊瑚礁海域魚類相似性最低;同時與調查方法也有一定關系,徐聞珊瑚礁海域的調查方法為定置網和刺網按季度調查(楊國歡等,2008),此法對體型較大魚類有較好的選擇性,而海南島珊瑚礁海域(吳鐘解等,2015)和西沙趙述島珊瑚礁海域(楊位迪等,2018)的魚類統計數據來自水下錄像,此法對小型魚類的發現概率較高。
4 結論
廣西潿洲島珊瑚礁海域魚類以暖水性為主,四季魚類物種組成有一定差異,且以底層和中下層魚類為主。廣西潿洲島珊瑚礁海域魚類物種組成與同緯度徐聞珊瑚礁海域的相似性最高,其魚類物種相對豐度較低且數量相對偏少,與受人類活動干擾有一定關系。因此,為更好地保護廣西潿洲島珊瑚礁海域魚類物種多樣性,建議有關部門采取相應措施減緩過度捕撈或旅游區開發等人類活動的影響。
參考文獻:
陳剛,趙美霞,劉斌,張春華,梁群. 2016. 基于Reef Check調查的潿洲島珊瑚礁生態狀況評價[J]. 熱帶地理,36(1):66-71. [Chen G,Zhao M X,Liu B,Zhang C H,Liang Q. 2016. Ecological situation of coral reefs in the Weizhou Island based on Reef Check[J]. Tropical Geography,36(1):66-71.]
陳國寶,李永振,陳新軍,舒黎明. 2006. 南海重要珊瑚礁過渡性水域軟骨魚類的組成與分布[J]. 上海水產大學學報,15(4):461-467. [Chen G B,Li Y Z,Chen X J,Shu L M. 2006. Composition and distribution of cartilaginous fishes in the adjacent waters area of main islands and reefs of South China Sea[J]. Journal of Shanghai Fisheries University,15(4):461-467.]
傅亮. 2014. 中國南海西南中沙群島珊瑚礁魚類圖譜[M]. 北京:中信出版社. [Fu L. 2014. Coral reef fishes of the South China Sea. The Xisha, Nansha and Zhongsha Islands[M]. Beijing:China Citic Press.]
賴廷和,何斌源. 2016. 廣西北部灣海洋硬骨魚類圖鑒[M]. 北京:科學出版社. [Lai T H,He B Y. 2016. Marine ostei-chthyse fishes in Guangxi Beibu Gulf of China[M]. Beijing:Science Press.]
黎廣釗,梁文,農華瓊,劉敬合. 2004. 潿洲島珊瑚礁生態環境條件初步研究[J]. 廣西科學,11(4):379-384. [Li G Z,Liang W,Nong H Q,Liu J H. 2004. Preliminary study on conditions of coral reef ecological environment along the coast of Weizhou Island[J]. Guangxi Sciences,11(4):379-384.]
李德越,李榮輝,吳志強,趙立朝,藍剛,丁洋. 2018. 廣西南寧大王灘水庫魚類物種組成及多樣性分析[J]. 南方水產科學,14(2):110-117. [Li D Y,Li R H,Wu Z Q,Zhao L C,Lan G,Ding Y. 2018. Species composition and diversity of fish in Dawangtan Reservoir in Nanning,Guangxi[J]. South China Fisheries Science,14(2):110-117.]
李永振. 2010. 西沙、中沙和南沙群島海域珊瑚礁魚類物種多樣性與生物學研究[D]. 青島:中國海洋大學. [Li Y Z. 2010. Species diversity and biology of fish in coral reef waters of Xisha,Zhongsha and Nansha Islands,South China Sea[D]. Qingdao:Ocean University of China.]
李永振,史赟榮,艾紅,董麗娜,李娜娜,李夏,高天翔. 2011. 南海珊瑚礁海域魚類分類多樣性大尺度分布格局[J]. 中國水產科學,18(3):619-628. [Li Y Z,Shi Y R,Ai H,Dong L N,Li N N,Li X,Gao T X. 2011. Large scale distribution patterns of taxonomic diversity of fish in coral reef waters,South China Sea[J]. Journal of Fishery Scien-ces of China,18(3):619-628.]
劉靜,吳仁協,康斌,馬琳. 2016. 北部灣魚類圖鑒[M]. 北京:科學出版社. [Liu J,Wu R X,Kang B,Ma L. 2016. Fishes of Beibu Gulf[M]. Beijing:Science Press.]
沈世杰. 1993. 臺灣魚類志[M]. 臺北:臺灣大學動物學系. [Shen S J. 1993. Fishes of Taiwan,China[M]. Taipei:Printing of Department of Zoology,Taiwan University.]
史海燕. 2012. 廣西北海潿洲島珊瑚礁海域生態環境監測與評價[D]. 青島:中國海洋大學. [Shi H Y. 2012. Monitoring and evaluation of ecological environment of coral reef areas in Weizhou Island,Beihai,Guangxi[D]. Qing-dao:Ocean University of China.]
史赟榮,李永振,盧偉華,孫冬芳. 2009. 東沙群島珊瑚礁海域魚類物種分類多樣性研究[J]. 南方水產,5(2):10-16. [Shi Y R,Li Y Z,Lu W H,Sun D F. 2009. Taxonomic diversity of fish species in coral reef area from Dongsha Islands[J]. South China Fisheries Sciences,5(2):10-16.]
孫典榮,林昭進,邱永松. 2005. 西沙群島重要島礁魚類資源調查[J]. 中國海洋大學學報,35(2):225-231. [Sun D R,Lin Z J, Qiu Y S. 2005. Survey of coral reef fish resources of the Xisha Islands[J]. Periodical of Ocean University of China,35(2):225-231.]
孫典榮,邱永松,林昭進,王雪輝. 2006. 中沙群島春季珊瑚礁魚類資源組成的初步研究[J]. 海洋湖沼通報,(3):85-92. [Sun D R,Qiu Y S,Lin Z J,Wang X H. 2006. Preliminary studies on the composition of coral reef fish resources in the waters of Zhongsha Island in spring[J]. Transactions of Oceanology and Limnology,(3):85-92.]
王文歡. 2017. 近30年來北部灣潿洲島造礁石珊瑚群落演變及影響因素[D]. 南寧:廣西大學. [Wang W H. 2017. Evolvement and influential factors of coral community over past three decades in Weizhou Island reef,Beibu Gulf[D]. Nanning:Guangxi University.]
吳鐘解,蔡澤富,陳石泉,張光星,李向民,王道儒,姚海君. 2015. 海南島東岸、南岸淺水礁區近10年珊瑚礁魚類種類分布及多樣性探討[J]. 水產學報,39(8):1203-1217. [Wu Z J,Cai Z F,Chen S Q,Zhang G X,Li X M,Wang D R,Yao H J. 2015. Species distribution and diversity of the coral reef fishes in the shallow reefs along the east and south coasts of Hainan Island[J]. Journal of Fishe-ries of China,39(8):1203-1217.]
楊國歡,侯秀瓊,陳春亮,孫省利. 2008. 徐聞珊瑚礁保護區魚類種類組成初步研究[J]. 水產科學,27(10):533-538. [Yang G H,Hou X Q,Chen C L,Sun X L. 2008. Fish species composition in the coral reef of coastal Xuwen County[J]. Fisheries Science,27(10):533-538.]
楊位迪,胡俊彤,林柏岸,黃海,劉敏. 2018. 西沙趙述島海域珊瑚礁魚類物種多樣性[J]. 廈門大學學報(自然科學版),57(6):819-826. [Yang W D,Hu J T,Lin B A,Huang H,Liu M. 2018. Species diversity of coral reef fishes in Zhaoshu Island waters,Xisha Islands[J]. Journal of Xiamen University(Natural Science),57(6):819-826.]
張鐿鋰,張雪梅. 1998. 植物區系地理研究中的重要參數──相似性系數[J]. 干旱區研究,15(1):59-63. [Zhang Y L,Zhang X M. 1998. Coeeficient of similarity—An important parameter in floristic geography[J]. Arid Zone Research,15(1):59-63.]
周浩郎,黎廣釗. 2014. 潿洲島珊瑚礁健康評估[J]. 廣西科學院學報,30(4):238-247. [Zhou H L,Li G Z. 2014. Assessment on the health of coral reefs at Weizhou Island[J]. Journal of Guangxi Academy of Sciences,30(4):238-247.]
Abaya L M,Wiegner T N,Colbert S L,Beets J P,Carlson K M,Kramer K L,Most R,Couch C S. 2018. A multi-indicator approach for identifying shoreline sewage pollution hotspots adjacent to coral reefs[J]. Marine Pollution Bulletin,129(1):70-80.
Bellwood D R,Hughes T P. 2001. Regional-Scale assembly rules and biodiversity of coral reefs[J]. Science,292(5521):1532-1535.
Bonnici L,Bonello J J,Schembri P J. 2018. Diet and trophic level of the longnose spurdog,Squalus blainville(Risso,1826) in the 25-nautical mile fisheries management zone around the Maltese Islands[J]. Regional Studies in Marine Science,19:33-42.
Burke L,Reytar K,Spalding M. Perry A. 2011. Reefs at risk revisited[R]. Washington D.C.:World Resources Institute.
Chen C A,Ablan M C A,McManus J W,Bell J D,Tuan V S,Cabanban A S,Shao K T. 2004. Population structure and genetic variability of six bar wrasse(Thallasoma hardwicki) in northern South China Sea revealed by mitochondrial control region sequences[J]. Marine Biotechnology(New York),6(4):312-326.
Chen T R,Li S,Yu K F,Zheng Z Y,Wang L R,Chen T G. 2013. Increasing temperature anomalies reduce coral growth in the Weizhou Island,northern South China Sea[J]. Estuarine,Coastal and Shelf Science,130(S1):121-126.
Hawkins J P,Roberts C M. 1996. The growth of coastal tou-rism in the Red Sea:Present and future effects on coral reefs[J]. Biological Conservation,76(2):216.
Heron S F,Maynard J A,van Hooidonk R,Eakin C M. 2016. Warming trends and bleaching stress of the worlds coral reefs 1985-2012[J]. Scientific Reports,6(1):38402.
Huang L L,Huang D L,Wu Z Q,Kang B,Chen Z B. 2016. Temporal variation of fish diversity and assemblages and their associations to environmental variables in the mangrove of Qinzhou Harbor,Guangxi Province,China[J]. Turkish Journal of Fisheries and Aquatic Sciences,16(2):297-310.
Hughes T P,Huang H,Young M A L. 2013. The wicked problem of Chinas disappearing coral reefs[J]. Conservation Biology,27(2):261-269.
Kulbicki M,Bozec Y M,Green A. 2005. Implications of biogeography in the use of butterflyfishes(Chaetodontidae) as indicators for Western and Central Pacific areas[J]. Aquatic Conservation Marine and Freshwater Ecosystems,15(S1):S109-S126.
MacNeil M A,Graham N A J,Cinner J E,Wilson S K,Williams I D,Maina J,Newman S,Friedlander A M,Jupiter S,Polunin N V,McClanahan T R. 2015. Recovery potential of the worlds coral reef fishes[J]. Nature,520(7547):341-344.
?hman M C,Rajasuriya A,?lafsson E. 1997. Reef fish assemblages in north-western Sri Lanka:Distribution pa-tterns and influences of fishing practises[J]. Environmental Biology of Fishes,49(1):45-61.
Ottimofiore E,Albouy C,Leprieur F,Descombes P,Kulbicki M,Mouillot D,Parravicini V,Pellissier L. 2017. Responses of coral reef fishes to past climate changes are related to life-history traits[J]. Ecology and Evolution,7(6):1996-2005.
Pinheiro H T,Bernardi G,Simon T,Joyeux J C,Macieira R M,Gasparini J L,Rocha C,Rocha L A. 2017. Island biogeography of marine organisms[J]. Nature,549(7670):82-85.
Polmear R,Stark J S,Roberts D,McMinn A. 2015. The effects of oil pollution on Antarctic benthic diatom communities over 5 years[J]. Marine Pollution Bulletin,90(1-2):33-40.
Richards B L,Williams I D,Vetter O J,Williams G J. 2012. Environmental factors affecting large-bodied coral reef fish assemblages in the Mariana Archipelago[J]. PLoS One,7(2):e31374.
Robertson D R. 1998. Do coral-reef fish faunas have a distinctive taxonomic structure?[J]. Coral Reefs,17(2):179-186.
Rodgers G G,Donelson J M,Mccormick M I,Munday P L. 2018. In hot water:Sustained ocean warming reduces survival of a low-latitude coral reef fish[J]. Marine Biology,165:73. doi:10.1007/s00227-018-3333-z.
Shi Q,Zhang Y C,Sun D H. 2003. Characteristics of growth rate of coral porites from Sanya Bay,Hainan Island and its relationship to environmental variables[J]. Marine Science Bulletin,5(2):62-71.
Spalding M D,Ravilious C,Green E P. 2001. World atlas of coral reefs[M]. California: University of California Press.
Webster J M,Carlos B J,Marc H,Potts D C,Yasufumi I,Yusuke Y,Fujita K,Bourillot R,Esat T M,Fallon S,Thompson W G,Thomas A L,Kan H,McGregor H V,Hinestrosa G,Obrochta S P,Lougheed B C. 2018. Response of the Great Barrier Reef to sea-level and environmental changes over the past 30000 years[J]. Nature Geoscience,11(6):426-432.
Zaneveld J R,Burkepile D E,Shantz A A,Pritchard C E,McMinds R,Payet J P,Welsh R,Correa A M,Lemoine N P,Rosales S,Fuchs C,Maynard J A,Thurber R V. 2016. Overfishing and nutrient pollution interact with temperature to disrupt coral reefs down to microbial scales[J]. Nature Communications,7:11833. doi:10.1038/ncomms11833.
(責任編輯 蘭宗寶)
