一株嗜鹽桿菌對(duì)中肋骨條藻的溶藻作用機(jī)理研究
2020-03-19 05:19:16韓貝貝張書(shū)飛吳風(fēng)霞史榮君黃洪輝
生態(tài)科學(xué) 2020年1期
關(guān)鍵詞:研究
韓貝貝,張書(shū)飛,吳風(fēng)霞史榮君黃洪輝
1.中國(guó)水產(chǎn)科學(xué)研究院南海水產(chǎn)研究所,廣東省漁業(yè)生態(tài)環(huán)境重點(diǎn)實(shí)驗(yàn)室,農(nóng)業(yè)農(nóng)村部南海漁業(yè)資源開(kāi)發(fā)利用重點(diǎn)實(shí)驗(yàn)室,廣州510300
2.上海海洋大學(xué)水產(chǎn)與生命學(xué)院,上海201306
0 前言
近年來(lái),藻華發(fā)生頻率增加、規(guī)模擴(kuò)大、危害加劇,已成為一個(gè)全球性的環(huán)境健康問(wèn)題[1]。而中肋骨條藻(Skeletonema costatum)作為一種廣溫廣鹽性近岸硅藻,廣泛分布在我國(guó)沿海地區(qū),每年在我國(guó)沿海都會(huì)引發(fā)藻華,對(duì)海洋生態(tài)系統(tǒng)造成嚴(yán)重影響。因此,尋求一種有效的藻華治理方法顯得尤為迫切。
海洋中細(xì)菌是微生物種群的重要組成部分,與藻類共同調(diào)控海洋生態(tài)系統(tǒng)的結(jié)構(gòu)和功能[2]。在藻際環(huán)境中存在著錯(cuò)綜復(fù)雜的菌藻互作關(guān)系,一方面,藻類對(duì)細(xì)菌生長(zhǎng)的促進(jìn)或抑制:藻類為細(xì)菌提供營(yíng)養(yǎng)物質(zhì),促進(jìn)細(xì)菌生長(zhǎng);藻類也可以通過(guò)釋放抗菌物質(zhì)殺死部分細(xì)菌[3-4],或通過(guò)抑制細(xì)菌的密度感應(yīng)系統(tǒng),抑制細(xì)菌的代謝[5]。另一方面,細(xì)菌對(duì)藻類生長(zhǎng)的促進(jìn)或抑制:細(xì)菌可以重新礦化藻源物質(zhì)為藻類的生長(zhǎng)提供無(wú)機(jī)營(yíng)養(yǎng)和必要的生長(zhǎng)因子[6],包括氮[7]、磷[8]、維生素[9]、鐵載體[10]和生長(zhǎng)素[11]等;有些溶藻菌可以直接或間接地抑制藻類的生長(zhǎng),甚至裂解藻細(xì)胞。近年來(lái),研究發(fā)現(xiàn),復(fù)雜的“藻—菌”關(guān)系在藻華的消亡過(guò)程中發(fā)揮著重要的作用。在藻華末期,溶藻菌的大量繁殖加速了藻華的衰退。溶藻菌的發(fā)現(xiàn)為藻華的控制提供了可能途徑,因此,海洋溶藻菌的溶藻作用成為藻華治理領(lǐng)域的研究熱點(diǎn)。
目前,已有大量關(guān)于溶藻菌的研究和報(bào)道,但關(guān)于嗜鹽桿菌的溶藻研究還鮮有報(bào)道,以往大多數(shù)文獻(xiàn)是關(guān)于嗜鹽桿菌的分離與基因組序列分析研究的報(bào)道[12-13],也有文獻(xiàn)報(bào)道該屬細(xì)菌在微生物工業(yè)發(fā)酵中的應(yīng)用[14]等,本實(shí)驗(yàn)室前期從深圳大鵬灣藻華爆發(fā)海域分離篩選到了一株嗜鹽桿菌(Halobacillus kuroshimensis,HSQAY1,Genbank 登陸號(hào) JX308828)[15],該菌株為革蘭氏陽(yáng)性、桿狀中等嗜鹽細(xì)菌,菌落呈圓形、黃橙色[12]。研究發(fā)現(xiàn)該菌株的上清液對(duì)中肋骨條藻具有強(qiáng)烈的溶藻作用,但對(duì)其溶藻作用機(jī)理尚不清楚。因此本研究以中肋骨條藻為研究對(duì)象,通過(guò)研究溶藻作用下中肋骨條藻細(xì)胞形態(tài)結(jié)構(gòu)、相關(guān)生理參數(shù)以及與氮代謝、抗氧化系統(tǒng)相關(guān)酶活性等的變化,對(duì)其中的溶藻作用機(jī)理進(jìn)行了探討,研究結(jié)果將拓展我們對(duì)藻華過(guò)程中“藻—菌”關(guān)系的認(rèn)識(shí),對(duì)藻華的生物治理具有重要的科學(xué)意義。
1 材料與方法
1.1 材料
1.1.1 溶藻細(xì)菌及無(wú)菌上清液的制備
嗜鹽桿菌菌株HSQAY1于深圳大鵬灣藻華爆發(fā)海域分離并保藏[15]。實(shí)驗(yàn)時(shí),用接種環(huán)挑取一環(huán)菌落接種至100 mL 2216E液體培養(yǎng)基中,28 ℃靜置活化24 h,再以1:100比例接種至2216E液體培養(yǎng)基中,28℃、160 rpm 振蕩培養(yǎng) 48 h,將培養(yǎng)后的菌液以6000 g離心20 min、0.22 μm濾膜過(guò)濾并用平板法檢驗(yàn)無(wú)菌后,獲得溶藻菌的無(wú)菌上清液(簡(jiǎn)稱上清液)。
1.1.2 實(shí)驗(yàn)藻種及培養(yǎng)條件
中肋骨條藻由近海海洋環(huán)境科學(xué)國(guó)家重點(diǎn)實(shí)驗(yàn)室(廈門大學(xué))海洋藻類保種中心提供。藻種經(jīng)活化后采用 f/2培養(yǎng)基[16]培養(yǎng),溫度20℃,光照強(qiáng)度100 μE·m-2·s-1,光暗周期 14 h:10 h。
1.2 實(shí)驗(yàn)方法
1.2.1 上清液最適濃度的確定及樣品收集
隨著國(guó)家“一帶一路”戰(zhàn)略的實(shí)施,中國(guó)走向世界的步伐加快,文化的交流也顯得尤為重要,國(guó)際學(xué)術(shù)文化、知識(shí)資源的相互交流與促進(jìn)將成為科技發(fā)展的重要推動(dòng)劑。黨的十九大報(bào)告中指出推動(dòng)文化事業(yè)和文化產(chǎn)業(yè)發(fā)展,推進(jìn)國(guó)際傳播能力建設(shè),講好中國(guó)故事,展現(xiàn)真實(shí)、立體、全面的中國(guó),提高國(guó)家文化軟實(shí)力。而國(guó)家學(xué)術(shù)文化、科技文化的發(fā)展能夠展現(xiàn)國(guó)家科技的實(shí)力與水平。
上清液分別按2.5%、5%、7.5%、10%(v/v)添加至指數(shù)期的中肋骨條藻藻液中,對(duì)照組為未添加上清液的正常培養(yǎng),另設(shè)置一組添加10%(v/v)的2216E培養(yǎng)基。各組每天相同時(shí)間取1 mL藻液,盧戈試劑染色后于光學(xué)顯微鏡下計(jì)數(shù),最終根據(jù)細(xì)胞計(jì)數(shù)結(jié)果繪制生長(zhǎng)曲線,確定最適的上清液處理濃度。
根據(jù)上清液最適濃度結(jié)果,取指數(shù)期的中肋骨條藻藻液,設(shè)置3組平行并分別加入5%(v/v)的上清液進(jìn)行培養(yǎng),分別收集處理0 h、2 h、12 h、24 h的藻液,其中15 mL藻液用于細(xì)胞形態(tài)結(jié)構(gòu)觀察、15 mL藻液用于葉綠素?zé)晒鉁y(cè)定、4 L藻液經(jīng)3 μm濾膜過(guò)濾,5000 g離心15 min后用于酶活性測(cè)定、20 mL藻液采用相同方法收集用于葉綠素a含量測(cè)定、200 mL藻液采用相同方法收集用于總氮含量測(cè)定。
1.2.2 細(xì)胞形態(tài)結(jié)構(gòu)觀察
向收集的藻液中加入 0.5%(v/v)戊二醛固定,光學(xué)顯微鏡(AX10;Carl Zeiss Meditec AG)下觀察細(xì)胞形態(tài)結(jié)構(gòu)并隨機(jī)選取50個(gè)細(xì)胞測(cè)量其單細(xì)胞長(zhǎng)度、單細(xì)胞直徑以及胞間支持突長(zhǎng)度。
1.2.3 總蛋白質(zhì)、葉綠素a及總氮含量的測(cè)定
總蛋白質(zhì)含量采用總蛋白含量測(cè)定試劑盒(南京建成生物工程公司)測(cè)定。將藻細(xì)胞重懸于生理鹽水,4℃超聲波破碎。細(xì)胞勻漿以4000 g、4 ℃離心10 min后取上清液,根據(jù)試劑盒操作步驟測(cè)定總蛋白質(zhì)含量。
葉綠素a含量采用甲醇提取法進(jìn)行測(cè)定[17]。向收集的藻細(xì)胞中加入5 mL甲醇混勻,4 ℃避光萃取24 h后,7000 g離心5 min取上清,用酶標(biāo)儀(xMark;Bio-rad)測(cè)定上清液在750 nm、665 nm和652 nm波長(zhǎng)處的吸光度。葉綠素a含量(μg·mL-1)的計(jì)算公式:
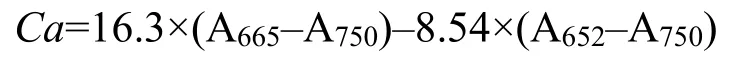
其中,A652、A665和A750分別代表上清液在652 nm、665 nm和750 nm處的吸光度,葉綠素a含量的最終結(jié)果以每107個(gè)細(xì)胞中葉綠素a含量表示(μg·107cell-1)。
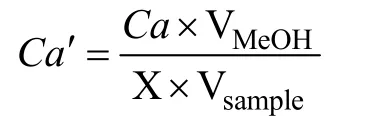
其中,VMeOH代表甲醇的體積(5 mL),Vsample代表樣本的體積(20 mL),X 代表藻細(xì)胞密度(cells·mL-1)。
細(xì)胞總氮含量參照Abreu等[18]的方法測(cè)定。將收集的藻細(xì)胞于60 ℃烘干24 h后稱其干重,烘干的樣品于研缽中磨成粉末狀,用35 mm×35 mm的錫紙將樣品包埋并精確稱重之后利用C/H/N/S/O元素分析儀(Vario ELIII;Elemetar Analysensysteme Gmbh)進(jìn)行測(cè)定(%),最終結(jié)果以每 109個(gè)細(xì)胞中氮含量表示(mg·109cell-1)。
1.2.4 酶活性的測(cè)定
將收集的藻細(xì)胞重懸于相應(yīng)勻漿介質(zhì)中,4 ℃超聲波破碎后細(xì)胞勻漿以4000 g、4 ℃離心10 min,上清液即為粗酶液。細(xì)胞抗氧化系統(tǒng)相關(guān)的丙二醛(MDA)含量、超氧化物歧化酶(SOD)、過(guò)氧化物酶(POD)活性以及細(xì)胞氮代謝相關(guān)的硝酸還原酶(NR)、亞硝酸還原酶(NiR)及谷氨酰胺合成酶(GS)活性均采用相應(yīng)試劑盒(南京建成生物工程公司、北京百奧萊博科技有限公司)進(jìn)行測(cè)定。
1.2.5 葉綠素?zé)晒鉁y(cè)定
藻細(xì)胞光合系統(tǒng) II(PS II)中的光合熒光效率使用超便攜式調(diào)制葉綠素?zé)晒鈨x(MINI-PAM- II;Walz)測(cè)定。將收集的藻液暗處理30 min后測(cè)定最大量子產(chǎn)量Fv/Fm。在光化光條件下(光照強(qiáng)度設(shè)定為90 μmol·m-2·s-1)測(cè)定實(shí)際量子產(chǎn)量Y(II)。
1.3 數(shù)據(jù)處理與分析
數(shù)據(jù)統(tǒng)計(jì)分析采用one-way ANOVA(SPSS 22.0 for Windows)檢驗(yàn)各個(gè)實(shí)驗(yàn)處理組之間的顯著性差異(p<0.05)。
2 結(jié)果
2.1 不同濃度上清液對(duì)藻細(xì)胞生長(zhǎng)的影響
細(xì)胞計(jì)數(shù)結(jié)果表明,加入 10%(v/v)2216E培養(yǎng)基的實(shí)驗(yàn)組與對(duì)照組的中肋骨條藻生長(zhǎng)趨勢(shì)無(wú)顯著差異(圖1),表明2216E培養(yǎng)基不影響中肋骨條藻的正常生長(zhǎng)。而加入不同濃度上清液的處理組中,溶藻效果隨著上清液濃度的升高而增強(qiáng),5%(v/v)上清液處理后24 h內(nèi)中肋骨條藻細(xì)胞處于抑制狀態(tài)且大部分細(xì)胞未破裂死亡,因此本研究選用5%(v/v)的上清液濃度進(jìn)行后續(xù)的實(shí)驗(yàn)。
2.2 溶藻作用下細(xì)胞形態(tài)結(jié)構(gòu)的變化
正常培養(yǎng)條件下,中肋骨條藻細(xì)胞呈鏈狀結(jié)構(gòu),單細(xì)胞呈凸鏡或短圓柱狀(圖2a、2b)。上清液處理2 h后,藻細(xì)胞的形態(tài)結(jié)構(gòu)與0 h無(wú)明顯變化,細(xì)胞鏈狀結(jié)構(gòu)及單細(xì)胞結(jié)構(gòu)完整(圖2c、2d)。上清液處理12 h后,部分細(xì)胞鏈狀結(jié)構(gòu)發(fā)生斷裂,細(xì)胞多以短鏈或單細(xì)胞存在,同時(shí)發(fā)現(xiàn)少量細(xì)胞形成泡狀(圖2e-2h)。24 h后,細(xì)胞多以單體存在,少見(jiàn)鏈狀細(xì)胞,部分泡狀細(xì)胞發(fā)生破裂降解(圖2i-2l)。

圖1 不同濃度上清液處理下中肋骨條藻的生長(zhǎng)曲線Figure 1 Growth curves of Skeletonema costatum after treated with different amounts of bacterial supernatant

圖2 上清液處理?xiàng)l件下中肋骨條藻細(xì)胞的形態(tài)結(jié)構(gòu)變化Figure 2 Variation of Skeletonema costatum cells structure after treated with bacterial supernatant
溶藻作用下,單細(xì)胞長(zhǎng)度、單細(xì)胞直徑以及細(xì)胞間支持突長(zhǎng)度的測(cè)量統(tǒng)計(jì)結(jié)果如圖3所示。上清液處理 2 h后,中肋骨條藻單細(xì)胞長(zhǎng)度顯著增加(p<0.01),12 h 后達(dá)到最大值,約為 0 h 組的 2.8 倍。而單細(xì)胞直徑和細(xì)胞間支持突長(zhǎng)度在各處理組間差異均不顯著。

圖3 上清液處理?xiàng)l件下中肋骨條藻細(xì)胞的大小變化Figure 3 Cell dimension variation of Skeletonema costatum after treated with bacterial supernatant
2.3 溶藻作用下藻細(xì)胞總蛋白質(zhì)、葉綠素a、總氮含量的變化
上清液處理2 h后,藻細(xì)胞內(nèi)總蛋白質(zhì)、葉綠素a以及總氮含量與0 h組無(wú)顯著差異,而處理12 h和24 h后,藻細(xì)胞內(nèi)總蛋白質(zhì)含量分別下降 44%和46%(p<0.01),葉綠素a含量分別下降48%和56%(p<0.01),總氮含量分別下降 41%和42%(p<0.01)(圖4)。
2.4 溶藻作用下藻細(xì)胞抗氧化系統(tǒng)、氮代謝相關(guān)酶活性的變化
上清液處理2 h后,細(xì)胞內(nèi)MDA含量與0 h組無(wú)顯著差異,而處理12 h后,藻細(xì)胞內(nèi)MDA含量急劇升高,顯著高于 0 h 組(p<0.01)。處理 24 h 后,藻細(xì)胞內(nèi)MDA含量雖略有下降,但仍顯著高于0 h組(p<0.05)(圖5a)。T-SOD活性在處理12 h后升高53%,顯著高于 0 h組(p<0.05)。隨著處理時(shí)間的延長(zhǎng),T-SOD活性繼續(xù)升高,24 h后活性達(dá)到0 h組的1.8倍(圖5b)。POD活性隨處理時(shí)間的延長(zhǎng)呈逐漸升高的趨勢(shì),各處理組均顯著高于0 h組(p<0.05)且處理12 h和24 h后活性分別升高至0 h組的5.6倍和6.1倍(圖5c)。
溶藻作用下,各處理組NR、NiR的活性均顯著低于0 h組(p<0.05),而GS的活性在處理2 h時(shí)無(wú)顯著變化,但在處理 12 h及 24 h后均顯著降低(p<0.01)(圖5d-5f)。
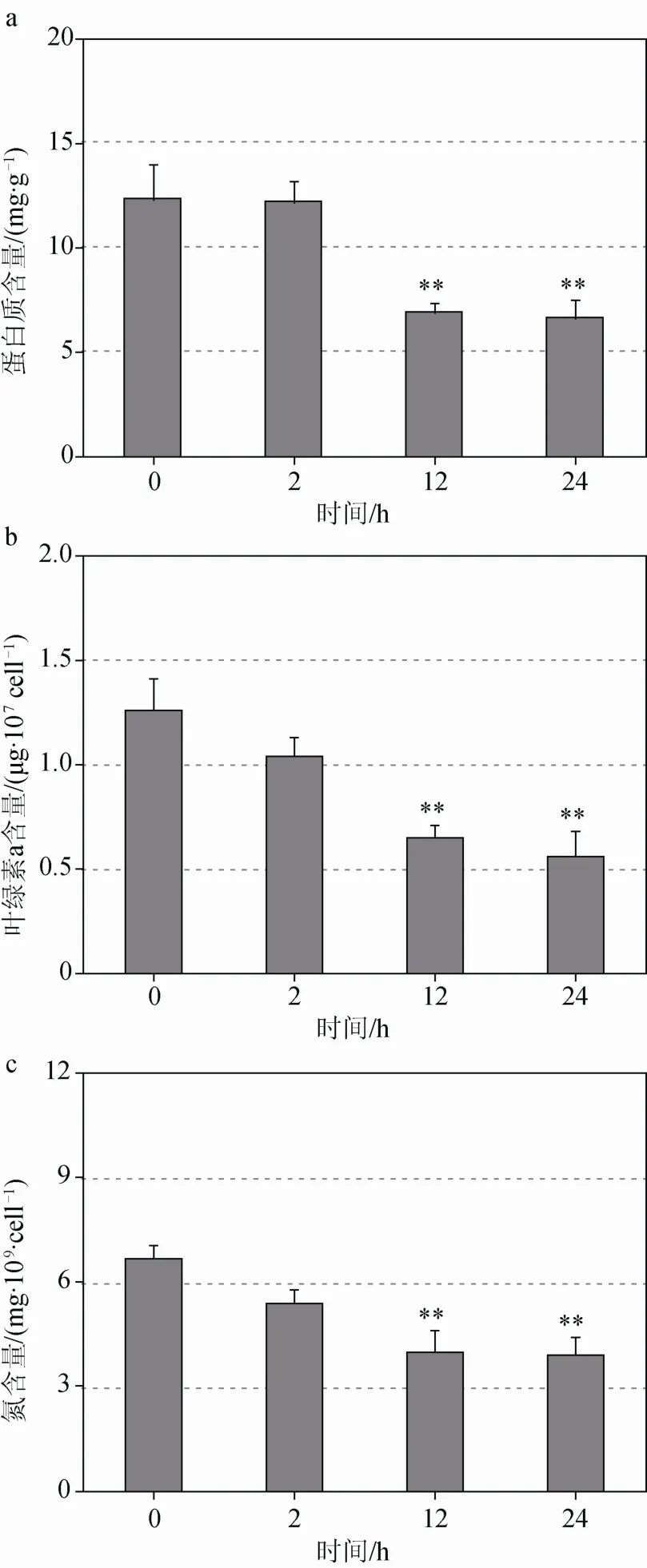
圖4 上清液對(duì)中肋骨條藻細(xì)胞蛋白質(zhì)、葉綠素 a、總氮含量的影響Figure 4 The effect of bacterial supernatant on the contents of protein, chlorophyll a and nitrogen of Skeletonema costatum cells
2.5 溶藻作用下藻細(xì)胞Fv/Fm、Y(II)的變化
溶藻作用下,各處理組藻細(xì)胞的Fv/Fm和Y(II)均顯著低于0 h組(p<0.01)。處理2 h后,F(xiàn)v/Fm和Y(II)分別下降 9%和 19%,且隨著處理時(shí)間的延長(zhǎng)持續(xù)下降,處理24 h后,F(xiàn)v/Fm和Y(II)均下降42%(p<0.01)(圖6)。
3 討論
3.1 細(xì)胞形態(tài)結(jié)構(gòu)變化
研究表明,不同種屬的骨條藻細(xì)胞在形態(tài)上存在一定的差異[19],相同種屬的骨條藻細(xì)胞長(zhǎng)度也可能隨生長(zhǎng)周期[20]或環(huán)境條件的變化而變化[21]。此外,當(dāng)骨條藻細(xì)胞進(jìn)入分裂期之后,細(xì)胞由于受到某些環(huán)境脅迫而無(wú)法分裂時(shí)細(xì)胞長(zhǎng)度也可能呈現(xiàn)增長(zhǎng)的狀態(tài)。如中肋骨條藻細(xì)胞在UVB紫外線脅迫條件下,細(xì)胞無(wú)法分裂且處于分裂之前的狀態(tài),從而導(dǎo)致細(xì)胞長(zhǎng)度變長(zhǎng)[22];Cu2+通過(guò)抑制硅藻細(xì)胞對(duì)硅的吸收利用,抑制細(xì)胞分裂,最終使細(xì)胞長(zhǎng)度增加[23]。類似的,本研究發(fā)現(xiàn),溶藻作用下中肋骨條藻細(xì)胞單細(xì)胞長(zhǎng)度顯著增加,結(jié)合細(xì)胞計(jì)數(shù)結(jié)果顯示5%(v/v)濃度的上清液處理下,24 h內(nèi)中肋骨條藻細(xì)胞數(shù)量處于相對(duì)穩(wěn)定的狀態(tài)(圖1),表明溶藻作用下藻細(xì)胞的分裂活動(dòng)受到抑制。因此,本研究中細(xì)胞長(zhǎng)度的增加可能是由于溶藻物質(zhì)抑制了細(xì)胞內(nèi)與細(xì)胞周期相關(guān)的代謝過(guò)程,或直接作用于細(xì)胞周期調(diào)控相關(guān)過(guò)程而使細(xì)胞周期停止,細(xì)胞無(wú)法分裂而一直處于分裂前細(xì)胞增長(zhǎng)的狀態(tài)。
3.2 氧化脅迫與藻細(xì)胞完整性
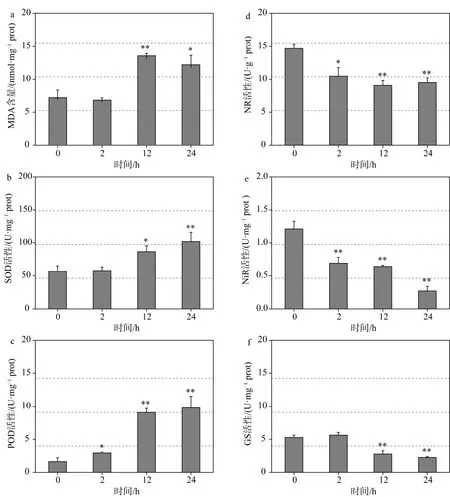
圖5 上清液對(duì)中肋骨條藻細(xì)胞抗氧化系統(tǒng)、氮代謝相關(guān)酶活性的影響Figure 5 The effect of bacterial supernatant on the activity of antioxidant and nitrogen metabolism enzymes of Skeletonema costatum cells

圖6 上清液對(duì)中肋骨條藻細(xì)胞Fv/Fm、Y(II)的影響Figure 6 The effect of bacterial supernatant on the Fv/Fm and Y(II) of Skeletonema costatum cells
活性氧(ROS)是細(xì)胞內(nèi)一類信號(hào)分子,在光合生物中主要由光合作用和呼吸作用等代謝過(guò)程產(chǎn)生,包括超氧陰離子、羥基自由基和過(guò)氧化氫等[24]。細(xì)胞內(nèi)ROS的生成和清除通常處于平衡狀態(tài),但外界條件的刺激可打破這種平衡而使細(xì)胞產(chǎn)生過(guò)量的ROS,從而引起脂質(zhì)過(guò)氧化,對(duì)細(xì)胞膜造成氧化損傷,破壞細(xì)胞膜的完整性與通透性。如 Liu等[25]發(fā)現(xiàn)添加抗生素后,中肋骨條藻細(xì)胞內(nèi)產(chǎn)生過(guò)量ROS,破壞了細(xì)胞膜的完整性與通透性,導(dǎo)致細(xì)胞內(nèi) pH增加以及細(xì)胞膨脹。Qian等[26]研究了鏈霉素對(duì)小球藻和銅綠微囊藻生理指標(biāo)的影響,結(jié)果顯示ROS含量增加的同時(shí)MDA含量以及電解液滲透量均增加,表明過(guò)量ROS會(huì)對(duì)細(xì)胞造成氧化損傷,致使細(xì)胞膜通透性增大。孔赟等[27]同樣發(fā)現(xiàn)在溶藻菌無(wú)菌濾液處理后,銅綠微囊藻細(xì)胞內(nèi) ROS含量增加,細(xì)胞膜完整性受到破壞,導(dǎo)致細(xì)胞內(nèi)K+和Ca2+外滲。吳培楓等[28]研究發(fā)現(xiàn)當(dāng)東海原甲藻受到溶藻菌 DH-e脅迫后,細(xì)胞ROS含量會(huì)急劇上升,同時(shí),MDA含量增加,SOD活性增強(qiáng)。管偉城等[29]在溶藻菌 LP-10溶藻機(jī)制的研究中也得到類似的結(jié)果:LP-10能夠刺激球形棕囊藻細(xì)胞產(chǎn)生過(guò)量 ROS,細(xì)胞內(nèi)部氧化水平升高,抗氧化系統(tǒng)中的SOD和POD等酶活性出現(xiàn)不同程度的升高,與本文研究結(jié)果相似。本研究中,溶藻作用下中肋骨條藻細(xì)胞 MDA 含量、SOD和POD的活性均顯著升高。SOD和POD活性的升高表明溶藻物質(zhì)刺激中肋骨條藻細(xì)胞產(chǎn)生了過(guò)量的ROS。而MDA含量的顯著升高則說(shuō)明ROS已對(duì)細(xì)胞膜內(nèi)脂質(zhì)造成損傷效應(yīng),因?yàn)?MDA 是脂質(zhì)過(guò)氧化的產(chǎn)物之一,也是衡量細(xì)胞內(nèi)脂質(zhì)過(guò)氧化程度的重要指標(biāo),間接反映了細(xì)胞膜系統(tǒng)的受損傷程度[30]。另外,光學(xué)顯微鏡的觀察結(jié)果發(fā)現(xiàn),在溶藻作用后期,中肋骨條藻細(xì)胞原生質(zhì)體會(huì)在細(xì)胞一端形成泡狀并逐漸膨脹破裂(圖2)。因此本研究推測(cè)溶藻作用發(fā)生的原因之一可能是由于中肋骨條藻細(xì)胞的細(xì)胞膜受到過(guò)量的ROS損傷而通透性發(fā)生變化,大量胞外物質(zhì)進(jìn)入細(xì)胞膜內(nèi)而使細(xì)胞膨脹破裂死亡。相關(guān)文獻(xiàn)也有類似的報(bào)道[31]。
3.3 溶藻物質(zhì)對(duì)中肋骨條藻細(xì)胞氮代謝的影響
氮是浮游植物生長(zhǎng)發(fā)育過(guò)程中的重要營(yíng)養(yǎng)元素,而硝態(tài)氮是藻類利用的主要形式[32]。硝態(tài)氮首先通過(guò)硝酸鹽轉(zhuǎn)運(yùn)蛋白的轉(zhuǎn)運(yùn)進(jìn)入細(xì)胞[33],再通過(guò) NR和NiR的催化轉(zhuǎn)化為能被細(xì)胞直接利用的銨態(tài)氮[34],隨后在 GS等酶的催化反應(yīng)下完成氨的同化,為葉綠素、核酸和蛋白質(zhì)等富氮化合物的合成提供原料[35]。在這一系列過(guò)程中,NR、NiR和GS是關(guān)鍵的調(diào)節(jié)酶,在氮代謝過(guò)程中發(fā)揮著重要作用。本研究發(fā)現(xiàn),溶藻作用下中肋骨條藻細(xì)胞內(nèi) NR、NiR、GS的活性均顯著降低,表明溶藻物質(zhì)抑制了藻細(xì)胞對(duì)氮的吸收利用,而這一抑制作用也可能影響了葉綠素、蛋白質(zhì)等富氮化合物的合成,導(dǎo)致處理組中葉綠素a、總蛋白質(zhì)以及細(xì)胞總氮含量均顯著低于 0 h組(圖4)。此外,有研究發(fā)現(xiàn)氮是藻類細(xì)胞分裂繁殖過(guò)程中的重要元素,因此,本研究中,氮的吸收利用受到抑制也可能是溶藻作用發(fā)生的一個(gè)原因,溶藻作用下中肋骨條藻細(xì)胞氮代謝過(guò)程受阻,細(xì)胞因缺氮而無(wú)法分裂。高越等[36]和Olson等[37]研究也證實(shí)了這一點(diǎn),在氮限制條件下,藻細(xì)胞分裂受到抑制,細(xì)胞周期停滯于G1期或G2/M期。
3.4 溶藻物質(zhì)對(duì)中肋骨條藻細(xì)胞光合系統(tǒng)的影響
光合作用是藻細(xì)胞中的重要代謝過(guò)程之一,為藻細(xì)胞提供有機(jī)物質(zhì)和能量,光合作用的強(qiáng)弱直接影響著藻細(xì)胞的正常生長(zhǎng)[38]。葉綠素a在光能收集和能量轉(zhuǎn)換過(guò)程中起著重要的作用,常被用來(lái)評(píng)估藻細(xì)胞對(duì)環(huán)境脅迫反應(yīng)的程度。Fv/Fm表示藻細(xì)胞PS II中的潛在最大光合能力,反映藻細(xì)胞最大光能轉(zhuǎn)換效率,可作為反映光合作用變化的重要指標(biāo)[39],而 Y(II)表示藻細(xì)胞的實(shí)際光合效率。傅麗君等[40]研究發(fā)現(xiàn)在溶藻細(xì)菌 BS03上清液處理后,塔瑪亞歷山大藻細(xì)胞內(nèi)葉綠素a含量和Fv/Fm隨處理時(shí)間的延長(zhǎng)和濃度的增加呈逐漸下降趨勢(shì),與本文研究結(jié)果相似。本研究中,溶藻作用下,藻細(xì)胞的葉綠素a 含量(圖4b)、Fv/Fm 和 Y(II)均顯著下降(圖6),表明溶藻物質(zhì)對(duì)藻細(xì)胞的光合系統(tǒng)可能造成了一定的損傷。研究表明,光化學(xué)反應(yīng)和電子傳遞主要依賴于類囊體上的電子載體以及相關(guān)酶的移動(dòng),光合作用受類囊體結(jié)構(gòu)和流動(dòng)性的影響[41],而過(guò)量的 ROS會(huì)對(duì)光合系統(tǒng)包括類囊體等造成氧化損傷,從而影響藻細(xì)胞的光合作用[42-43]。因此,本研究中 Fv/Fm和Y(II)被顯著抑制可能與類囊體膜受氧化損傷而不完整有關(guān),MDA含量的顯著升高也間接地印證了這一推斷。Yang等[24]也認(rèn)為靈菌紅素是通過(guò)破壞銅綠微囊藻類囊體膜的完整性而使光合作用功能受損。此外,氮對(duì)藻細(xì)胞光合作用中光反應(yīng)和暗反應(yīng)的一系列酶的活性也有重要影響,在一定范圍內(nèi),藻細(xì)胞的光合速率與氮的營(yíng)養(yǎng)水平呈正相關(guān)[44]。因此,溶藻作用下,中肋骨條藻細(xì)胞內(nèi)氮含量的降低也可能是藻細(xì)胞光合速率下降的原因之一。
4 結(jié)論
本研究以中肋骨條藻為研究對(duì)象,通過(guò)顯微觀察以及對(duì)相關(guān)生理參數(shù)及酶活性的測(cè)定,探討了嗜鹽桿菌HSQAY1對(duì)中肋骨條藻的溶藻作用機(jī)理。根據(jù)本研究結(jié)果推測(cè)嗜鹽桿菌上清液中的溶藻物質(zhì)可顯著抑制中肋骨條藻對(duì)氮的吸收利用,細(xì)胞內(nèi)蛋白質(zhì)、葉綠素a等重要生命物質(zhì)的合成受阻,細(xì)胞的正常代謝過(guò)程受到抑制,最終細(xì)胞分裂活動(dòng)停止。同時(shí)溶藻物質(zhì)的脅迫刺激中肋骨條藻細(xì)胞產(chǎn)生過(guò)量的活性氧,細(xì)胞膜系統(tǒng)受到氧化損傷而使其通透性發(fā)生變化,大量胞外物質(zhì)透過(guò)細(xì)胞膜進(jìn)入細(xì)胞內(nèi),細(xì)胞最終膨脹破裂死亡。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
國(guó)際商務(wù)財(cái)會(huì)(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
