植物乳桿菌最適生長底物解析及高密度培養工藝
2020-03-19 01:02:22王玉林黃潔崔樹茂唐鑫毛丙永趙建新張灝陳衛
食品與發酵工業 2020年4期
王玉林,黃潔,崔樹茂*,唐鑫,毛丙永,趙建新,張灝,陳衛
1(江南大學 食品學院,江蘇 無錫,214122)2(蘇州硒泰克生物科技有限公司,江蘇 蘇州,215200)
植物乳桿菌是發酵食品中常見的乳酸菌,其作為發酵劑應用于發酵食品生產時,可以避免繼代培養引起的活力退化和雜菌污染等問題[1-2],并可顯著提高生產效率和改善發酵風味。近幾十年的研究亦發現,植物乳桿菌具有很廣泛的益生作用,如降低膽固醇、降血糖、抗菌消炎、提高人體免疫力等益生功能[3-8]。隨著物乳桿菌的應用越來越廣泛,提高植物乳桿菌的增殖濃度,進而提高生產效率、降低成本,對植物乳桿菌的進一步推廣具有重要的促進意義。
營養底物與高密度培養技術是影響植物乳桿菌生長繁殖的主要因素。細胞生長所必需的成分包括碳源、氮源、無機鹽、促生長物質等,這些營養成分顯著影響乳酸菌生長速率及最大生物量。其中,碳源、氮源是乳酸菌生長繁殖不可或缺的營養物質。葡萄糖為單糖且價格低廉,是益生菌的最佳利用碳源[9-10],微生物生長代謝與產物合成需要氮源,乳酸菌對不同氮源的利用效率不同,尋求經濟、利用效果好的食品級氮源對于菌株發酵培養至關重要[11-12]。提高植物乳桿菌增殖濃度,不僅要優化培養基營養成分,而且需要改善高密度培養技術手段。高密度培養是通過改善發酵工藝,提高單位體積發酵液的生物量[13]。目前主要的高密度培養技術有恒pH培養、循環培養、固定化培養、過濾培養等[1]。恒pH培養因其操作簡單,易于控制,依然是國內乳酸菌工業化生產的主流方式,但是恒pH培養方式仍有待解決兩個問題:分批培養過程中出現的碳氮源缺乏或初始濃度過高抑制菌株生長;補料模式不科學,未根據消耗規律科學補料且非智能化控制,幾乎無法實現工業化應用[14]。
本研究通過解析植物乳桿菌的關鍵生長底物,并基于滲透壓和最適底物優化恒pH分批培養和恒pH分批自動反饋補料培養,從而提高菌株單位體積的活菌數,為提高植物乳桿菌的工業化生產效率奠定基礎。
1 材料與方法
1.1 菌株與試劑
1.1.1 菌株
L.plantarumX1、L.plantarumN8、L.plantarumN9,來源于江南大學食品生物中心菌種保藏中心。
1.1.2 試劑
胰蛋白胨、牛肉膏、酵母粉、無水葡萄糖、無水乙酸鈉、MgSO4·7H2O、MnSO4·H2O、檸檬酸氫二鈉、無水K2HPO4、吐溫80,國藥集團化學試劑有限公司;葡萄糖測定試劑盒,上海榮盛生物藥業有限公司;酵母蛋白FM103、酵母浸粉FM803、酵母浸粉FM528、酵母提取物FD00-A、大豆蛋白胨FP410、魚骨蛋白胨FP350,安琪酵母股份有限公司;胰蛋白胨AL-17445、水解植物蛋白AL-17446,上海創賽科技有限公司。
1.2 儀器與設備
高速冷凍離心機,Eppendorf公司;l?ser-om806m型冰點滲透壓測定儀,德國l?ser公司;T&J-Minipod 1L×8型迷你平行發酵罐,迪必爾生物工程有限公司;GRP-9080型隔水式恒溫培養箱,上海森信實驗儀器有限公司;FE20型pH計、EL3002型電子天平,梅特勒-托利多儀器有限公司;MS 3 basic型渦旋振蕩器,德國IKA公司;MLS-3750型高溫高壓滅菌鍋,日本SANYO公司;ZHJH-C1115B型超凈工作臺,上海智誠分析儀器制造有限公司;UV-2450紫外分光光度計,日本島津公司。
1.3 實驗方法
1.3.1 植物乳桿菌的活化和培養
從冰箱取出冷凍的保菌管,置于冰盒中冰浴溶解后,使用接種環挑取種子液在固體培養基上劃線,置于37℃恒溫培養箱中培養24~48 h,使菌種復壯。挑取單菌落接種于5 mL液體MRS培養基中,在37℃ 恒溫培養箱中培養18~24 h。重復上述操作2次得到活力旺盛的種子液用于后續實驗。
1.3.2 生長曲線測定
將活力旺盛的種子液以1%接種量接種于新鮮MRS培養基中,置于37 ℃ 恒溫培養箱培養,分別取2、4、6、8、10、12 h的菌懸液測波長600 nm處的OD值。以培養時間為橫坐標,各時間點OD600為縱坐標,繪制植物乳桿菌生長曲線。
1.3.3 氮源利用分析
不同氮源培養基(g/L):氮源1(酵母蛋白FM103、酵母浸粉FM803、酵母浸粉FM528、酵母提取物FD00-A、大豆蛋白胨FP410、魚骨蛋白胨FP350、胰蛋白胨AL-17445、水解植物蛋白AL-17446)、葡萄糖6、無水K2HPO410、Na2HPO410、MgSO4·qH2O0.25、MnSO4·H2O 0.05,對照組氮源為胰蛋白胨0.4、牛肉膏 0.4、酵母粉 0.2,培養基滅菌后接入1%種子液,置于37℃恒溫培養箱中靜置培養,待菌株生長至穩定期測定OD600。
1.3.4 生長限制性無機鹽的測定
用NaCl替換MRS培養基中的無機鹽(K2HPO4、檸檬酸氫二銨和乙酸鈉),以MRS培養基作為對照,調節滲透壓保持一致。各培養基以2/3的裝樣量置于500 mL的平行發酵罐,滅菌后接入5%種子液擴大培養,以縮短發酵周期和減少染菌機會。控制發酵溫度37℃,恒pH 6.0培養,每隔2 h取樣測定發酵液的OD600和活菌數。
1.3.5 微量元素(Mn)最適濃度的測定
按最適氮源25 g/L,葡萄糖20 g/L,微量元素MnSO4質量濃度分別為0.05、0.15、0.25、0.35 g/L,配制不同MnSO4濃度的培養基,各培養基以2/3的裝樣量置于平行發酵罐,滅菌后接入5% 種子液擴大培養,控制發酵溫度37 ℃,恒pH 6.0自動反饋補糖培養, 待菌株生長至穩定期測定活菌數。
1.3.6 促生長物質分析
采用1.3.3方法,以實驗菌株的最佳氮源為培養基氮源,添加不同種類的促生長物質0.3 g/L (核苷酸(I+G)、核苷酸、腺嘌呤、復合維生素、VB族、胡蘿卜粉、番茄粉),對照組不添加促生長物質,測定菌株穩定期OD600和活菌數。
1.3.7 最適生長pH的測定
將MRS中氮源替換成最佳氮源配制培養基,各培養基以2/3的裝樣量置于平行發酵罐,滅菌后接入5%種子液擴大培養,控制發酵溫度37℃,在不同pH條件下(pH 5.0、5.5、6.0、6.5)恒pH培養至穩定期,測定發酵液活菌數。
1.3.8 最適生長滲透壓及完全抑制滲透壓的測定
MRS培養基滲透壓為350 mOsm/kg,1 L MRS中添加3 g NaCl,滲透壓可增加100 mOsm/kg;添加NaCl調節MRS培養基的滲透壓,配制350、500、600、700、800、900、1 200、1 500、2 400、2 700、3 000 mOsm/kg不同滲透壓條件的培養基。各培養基滅菌后接入1%種子液,置于37℃恒溫靜置培養;采用方法1.3.2,以時間為橫坐標,lg OD600為縱坐標繪制植物乳桿菌在不同滲透壓環境下生長曲線。
1.3.9 碳氮消耗比的測定
采用1.3.3方法的培養基配方,將最佳氮源作為培養基氮源。在發酵過程中不斷監測發酵液中葡萄糖濃度和OD600,計算菌株生長速率被抑制時糖消耗量(碳氮比)。根據碳氮消耗比,配制不同梯度的碳氮比培養基,各培養基以2/3的裝樣量置于平行發酵罐,滅菌后接入5%種子液擴大培養,控制發酵溫度37℃,恒pH 6.0自動反饋補糖培養, 待菌株生長至穩定期測定活菌數。
1.3.10 恒pH分批培養
根據培養基營養底物分析,確定菌株培養基最優底物組成。根據可耐受滲透壓與最佳碳氮比,推算培養基中氮源含量,培養基中碳氮源初始濃度低于抑制生長滲透壓的原則進行設定3個滲透壓梯度。各培養基以2/3的裝樣量置于平行發酵罐,滅菌后接入5%種子液擴大培養,控制發酵溫度37℃,以最優pH進行恒pH培養至穩定期,測定發酵液活菌數。
1.3.11 恒pH自動反饋補料培養[15-18]
根據培養基營養底物分析、最適碳氮比、可耐受滲透壓與完全抑制滲透壓,優化恒pH反饋補料工藝;(1)推算恒 pH 培養時菌體被完全抑制時消耗的氮源,配制滲透壓為(350±20) mOsm/kg的培養基,補料液為400 g/L葡萄糖;(2)根據碳氮比配制環境滲透壓為(350±20) mOsm/kg培養基,補料液為碳氮混合液(葡萄糖質量濃度都為200 g/L,氮源濃度按照最佳碳氮消耗比添加)。培養基以2/3的裝樣量置于平行發酵罐,滅菌后接入5%種子液擴大培養,控制發酵溫度37℃,在最適pH條件下恒pH培養至穩定期,測定發酵液活菌數。
1.3.12 分析方法
測定生物量,采用光密度法;測定葡萄糖濃度,采用葡萄糖氧化酶-過氧化物酶法;活菌數,采用平板菌落計數法。
1.3.13 數據統計與分析
試驗中每個實驗值均為3次平行實驗的平均值,在后期的統計分析中不再重復強調。
2 結果與分析
2.1 植物乳桿菌氮源利用選擇性分析
由圖1可知,3株植物乳桿菌的生長趨勢相同,菌株發酵均在8~12 h達到穩定期,此時生物量最大且基本保持穩定,測定穩定期活菌數。

a-植物乳桿菌生長曲線圖;b-植物乳桿菌穩定期活菌數圖1 植物乳桿菌生長曲線和最大生物量Fig.1 Growth curve and viable counts of L.plantarum in MRS medium注:不同小寫字母表示具有顯著性差異(P<0.05)(下同)
植物乳桿菌在發酵培養過程中容易受到酸和碳源不足的抑制,發酵結束后發酵液中仍有氮源剩余,從而無法比較同等質量氮源對菌株的增殖差異。氮源篩選培養基可以提高培養基中碳氮比、增強無機鹽緩沖能力,從而緩解發酵過程中的酸抑制及彌補發酵體系中碳源不足。
采用氮源篩選方法1.3.3,分析單位質量氮源對植物乳桿菌增殖效果。從表1可以看出,3株植物乳桿菌對微生物類氮源的利用具有偏好性,特別是對酵母蛋白FM103、酵母浸粉FM803與酵母浸粉FM528的利用。植物乳桿菌X1、N8、N9的最佳利用氮源分別為酵母浸粉FM803、酵母浸粉FM 528、酵母浸粉FM 803,植物乳桿菌利用1g最佳氮源的增殖效果(OD600)分別達到1.48±0.02、1.56±0.01、1.82±0.07。
為了進一步探究不同種類氮源條件下菌株增殖差異是否因氮源含有生長因子,研究復合氮源對菌株的增殖效果。將增殖效果差的氮源和可能含有生長因子的酵母類氮源按照1∶1比例復配,分析對3株植物乳桿菌的增殖效率。通過計算得知,增殖效果差的氮源與含生長因子的酵母復配并未提高最大生物量。
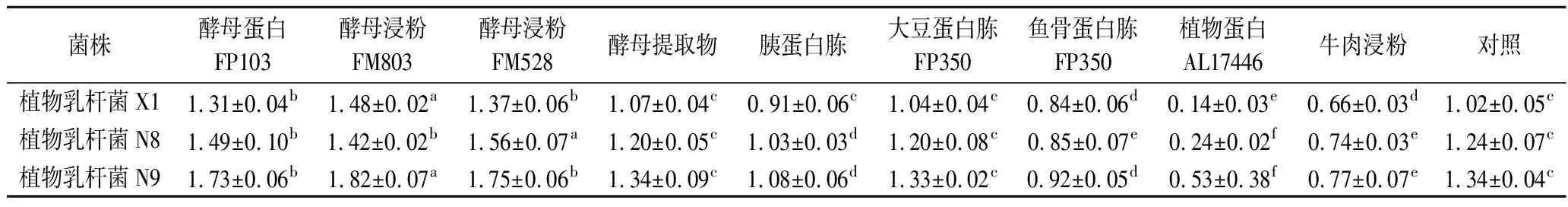
表1 不同氮源發酵液分批培養植物乳桿菌的最高生物量(OD600)Table 1 Maximum biomass of L. plantarum in batch culture with different nitrogen sources
注:不同小寫字母表示具有顯著性差異(P<0.05)(下同)
綜上,植物乳桿菌對微生物氮源利用具有偏好性,對其他種類氮源利用較差。即使在增殖效果較差的氮源中添加增殖效果好酵母粉也并沒有促進菌株對其利用,說明植物乳桿菌對不同種類氮源利用效果的不同,不是由氮源中生長因子導致的,而是因氮源中有效成分。該結果進一步證明氮源中肽含量是乳酸菌生長的限制因素[19-20]。
2.2 生長限制性無機鹽
不同微生物生長繁殖需要的無機鹽不同,不同無機鹽在發酵培養中作用不同。乙酸鹽、檸檬酸鹽與磷酸鹽具有緩沖作用,可以避免乳酸菌在發酵過程中因 pH 迅速下降而死亡[21]。微生物的胞內代謝反應需要高鉀低鈉生態環境,防止細胞水腫,勢能儲備[22]。
由圖3可知,在培養基中添加緩沖鹽,僅僅起到緩沖劑的作用,對植物乳桿菌生長無促進作用。因此,植物乳桿菌在恒pH培養時培養基中無需添加無機鹽。
2.3 微量元素(Mn)最適濃度
不同乳桿菌的代謝途徑不同,不同代謝途徑中關鍵酶也不同,因此,乳桿菌生長繁殖所需要的微量元素不同。植物乳桿菌代謝屬于同型發酵,葡萄糖發酵利用糖酵解途徑(embden-meyerhof-parnas,EMP),主要代謝產物是乳酸。在EMP途徑中,丙酮酸與NADH在乳酸脫氫酶作用下生成乳酸,Mn不僅對NADH氧化有巨大的刺激作用[23],還是乳酸脫氫酶的組成成分[24],因此Mn是EMP途徑中關鍵代謝酶的激活劑[25]。
由圖4可知,植物乳桿菌最大生物量與低濃度微量元素呈正相關。當質量濃度高于0.25 g/L,持續增高并未顯著提高活菌數。因此,植物乳桿菌的MnSO4最適添加量為0.25 g/L。

a-植物乳桿菌X1;b-植物乳桿菌N8;c-植物乳桿菌N9;圖2 復合氮源發酵液分批培養植物乳桿菌的最高生物量(OD600)Fig.2 Maximum biomass of L. plantarum in batch culture with different composite nitrogen sources注:1-酵母浸粉803+大豆蛋白陳;2-酵母浸粉803+酵母提取物;3-酵母浸粉803+牛肉浸粉;4-酵母浸粉803+水解植物蛋白;5-酵母浸粉803+胰蛋白腺;6-酵母浸粉803+魚骨蛋白膝;7-酵母浸粉803

a、b、c-植物乳桿菌X1、N8、N9的生長曲線;d-植物乳桿菌X1、N8、N9利用無機鹽恒pH培養的活菌數圖3 植物乳桿菌利用無機鹽恒pH培養生長曲線和活菌數Fig.3 Growth curve and viable counts of L. plantarum in mediums containing different inorganic salts under pH-controlled condition
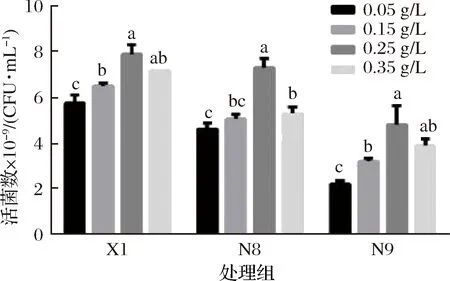
圖4 植物乳桿菌利用不同濃度微量元素MnSO4恒pH補料培養的活菌數Fig.4 Proliferation concentration of L.plantarum cultured in mediums with different manganese concentrationsunder pH-controlled condition
2.4 促生長物質
促生長因子是可以刺激微生物的生長活性、調節細胞生長和細胞生理功能的多肽物質。本實驗以酵母粉為基礎氮源,添加生長因子分析植物乳桿菌的促生長物質。從圖5可知,以酵母浸粉類為氮源時,不需要額外添加其他促生長物質。
2.5 最適生長pH值
在培養基或環境中,pH從多方面影響著植物乳桿菌生命活動,pH不僅影響胞內外酶的生物活性,還影響影響植物乳桿菌對營養物質的吸收以及改變培養基中化合物離子化程度,由而促進或抑制乳酸菌的繁殖生長。不同菌株的最適生長pH條件不同,由圖6可知,植物乳桿菌X1、植物乳桿菌N8最適生長pH條件為 5.5,植物乳桿菌N9最適生長pH條件為6.0。
2.6 最適生長滲透壓和完全抑制滲透壓
滲透壓是影響微生物生長繁殖的環境因子之一,通過影響細胞內水分活度、細胞膜通透性、蛋白質分子結構與細胞生理功能,從而影響菌株生長繁殖。乳桿菌在生長繁殖過程中產生代謝產物會引起滲透壓升高,因此菌株高密度培養最大生物量與乳桿菌環境耐滲性有關[2, 26]。
由圖7可知,植物乳桿菌X1在培養基滲透壓超過800 mOsm/kg時生長速率被抑制,滲透壓達到2 700 mOsm/kg時菌株生長完全被抑制; 植物乳桿菌N8在培養基滲透壓超過800 mOsm/kg時生長速率被抑制,滲透壓達到3 000 mOsm/kg時菌株生長完全被抑制;植物乳桿菌N9在培養基滲透壓超過1 200 mOsm/kg時生長速率被抑制,滲透壓達到2 700 mOsm/kg時菌株生長完全被抑制。

a-分別以植物乳桿菌X1;b-植物乳桿菌N8;c-植物乳桿菌N9圖5 植物乳桿菌分析促生長物質(OD600)Fig.5 Maximum biomass of L. plantarum in mediums with different composite nitrogen sources注:1-酵母浸粉803;2-酵母浸粉803+I+G;3-酵母浸粉803+VB族;4-酵母浸粉803+番茄粉;5-酵母浸粉803+復合核苷酸;6-酵母浸粉803+復合維生素;7-酵母浸粉803+胡蘿卜粉;8-酵母浸粉803+腺嘌呤
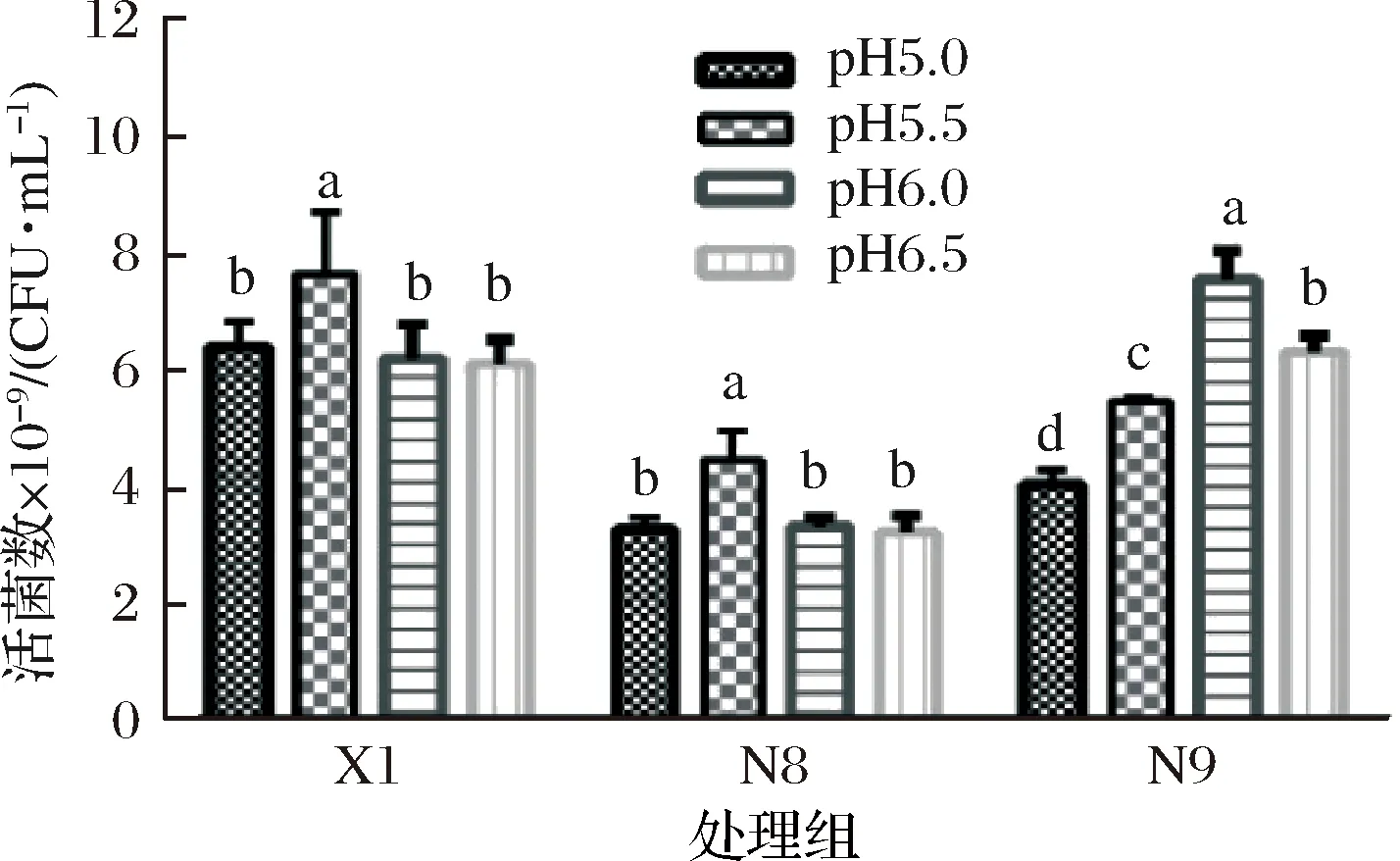
圖6 植物乳桿菌在不同pH條件下恒pH培養的活菌數Fig.6 Proliferation concentration of L. plantarumcultured under different pH-controlled condition
2.7 碳氮消耗比
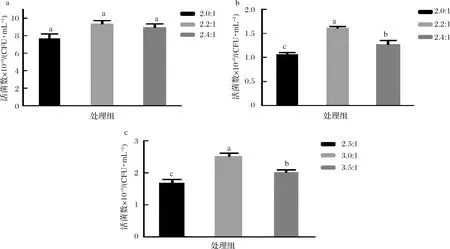
基于氮源優化結果,選擇增殖效果最佳的氮源,采用快速篩選氮源的培養基和方法,測定植物乳桿菌生長速率拐點的葡萄糖消耗量,即碳氮消耗比,結果如表2所示。
根據碳氮消耗比,設置不同梯度的碳氮比,使培養基初始滲透壓低于影響各自菌株生長速率的滲透壓值。由圖8可知,過高的碳氮比沒有促進菌株的生長,反而會對菌株的生長速率有一定影響,因此植物乳桿菌X1、N8、N9的最佳碳氮比分別為2.2 ∶1、2.2 ∶1、3.0 ∶1;
a-植物乳桿菌X1;b-植物乳桿菌N8;c-植物乳桿菌N9圖7 植物乳桿菌在不同滲透壓條件下的生長曲線Fig.7 Proliferation concentration of L.plantarum in different osmotic pressure

表2 植物乳桿菌生長速率被抑制時碳氮消耗比Table 2 Consumption ratio of carbon to nitrogen when the growth rate of L.plantarum is inhibited
2.8 恒pH分批培養
葡萄糖為單糖且價格低廉,是益生菌的最佳利用碳源[9-10],理論上菌株生長繁殖最大生物量與碳源含量呈正相關,但當發酵液的滲透壓高于最適生長滲透壓時,菌株的生長速率受到抑制滲透壓,因此探究恒pH培養工藝的最適底物濃度至關重要(圖9)。
a-分別為植物乳桿菌X1;b-植物乳桿菌N8;c-植物乳桿菌N9圖8 不同碳氮比恒 pH 培養的活菌數Fig.8 The viable counts with different carbon and nitrogen ratios under pH-controlled condition
2.8.1 植物乳桿菌X1恒pH分批培養
由上述結果可知,菌株的最佳氮源為酵母浸粉FM803、最適MnSO4濃度為0.25 g/L、最佳碳氮比2.2 ∶1、影響菌株生長的初始滲透壓為800 mOsm/kg以及完全抑制滲透壓為2 700 mOsm/kg。通過計算,配制酵母浸粉FM803 30、35、40 g/L,碳氮比2.2 ∶1,MnSO40.25 g/L,培養基環境滲透壓分別為(600±5)、(700±7)、(800±9)mOsm/kg,恒pH 5.5分批培養。發酵培養12h達到穩定期,發酵液滲透壓均低于1 800 mOsm/kg。碳氮比同等條件下,酵母浸粉FM803 40 g/L,葡萄糖88 g/L,MnSO40.25 g/L對植物乳桿菌X1增殖效果好,活菌數達到3.1×1010CFU/mL。
2.8.2 植物乳桿菌N8恒pH分批培養
配制酵母浸粉FM528 30、35、40 g/L,碳氮比2.2 ∶1,MnSO40.25 g/L,3種培養基的滲透壓分別為(585±5)、(695±7)、(784±9) mOsm/kg,恒pH 5.5分批培養,發酵培養12 h達到穩定期,發酵液滲透壓低于2 200 mOsm/kg,沒有達到完全抑制滲透壓。碳氮比同等條件下,酵母浸粉FM528 40 g/L,葡萄糖88,MnSO40.25 g/L分批培養對植物乳桿菌N8增殖效果好,葡萄糖幾乎沒有剩余,最終的活菌數可以達到2.9×1010CFU/mL。
2.8.3 植物乳桿菌N9恒pH分批培養
配制酵母浸粉FM 528 35、40、45 g/L,碳氮比3∶1,MnSO40.25 g/L,3種培養基的滲透壓分別為(875±7)、(1 000±6)、(1 125±9) mOsm/kg,恒pH 6.0分批培養。發酵培養12 h至穩定期,發酵后環境滲透壓低于2 200 mOsm/kg,沒有達到完全抑制滲透壓。碳氮比同等條件下,氮源45 g/L,葡萄糖135 g/L對植物乳桿菌X1增殖效果好,最終的活菌數可以達到6.2×1010CFU/mL。
2.9 恒pH自動反饋補料培養
由圖10可知,植物乳桿菌X1最佳補料工藝為酵母浸粉FM803 40 g/L,葡萄糖15 g/L,MnSO40.25 g/L,持續補糖36 mL,葡萄糖補料液濃度為400 g/L,活菌數達到4.1×1010CFU/mL。植物乳桿菌N8補糖補料工藝與補糖氮補料工藝的活菌數沒有顯著差別,均可達到2.2×1010CFU/mL。植物乳桿菌N9的補糖氮工藝活菌數比補糖工藝顯著增高,最佳補料工藝為酵母浸粉FM803 15 g/L,葡萄糖45 g/L,MnSO40.25 g/L,持續補料150 mL,該補料液為碳氮比為3∶1(酵母浸粉FM803 50 g/L),活菌數可以達到5.4×1010CFU/mL。

a-植物乳桿菌X1分批培養活菌數;b-植物乳桿菌N8分批培養活菌數;c-植物乳桿菌N9分批培養活菌數圖9 植物乳桿菌恒pH分批培養的活菌數Fig.9 Proliferation concentration of L.plantarum during constant pH feeding culture
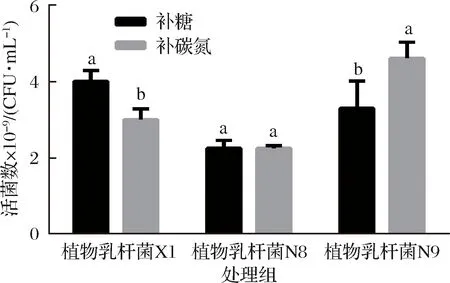
圖10 植物乳桿菌恒pH自動反饋補料工藝Fig.10 Proliferation concentration of L.plantarumduring pH-controllde using the automatic feedbackfeeding method
3 結論
植物乳桿菌對不同種類氮源的利用具有偏好性,其可高效利用酵母類氮源,對胰蛋白胨和大豆蛋白胨利用較差,幾乎不利用植物水解蛋白胨。
植物乳桿菌對氮源的偏好性歸因去菌體自身不能利用非酵母類氮源的肽,與促生長因子無關。且以酵母浸粉類為氮源時,不需要額外添加其他促生長物質。
緩沖鹽在分批培養時通過減緩pH的降低提高乳酸菌的生物量,但在恒pH培養時緩沖鹽對對植物乳桿菌的生長沒有促進作用。
植物乳桿菌生長濃度與微量元素濃度呈正相關,當培養基中MnSO4濃度低于0.25 g/L時,濃度越高菌株能達到的最高生物量越高,但過高的微量元素濃度會抑制菌株生長。
植物乳桿菌在恒pH培養時滲透壓是其主要抑制因素,植物乳桿菌X1在培養基滲透壓超過800 mOsm/kg時生長速率被抑制,滲透壓達到2 700 mOsm/kg時菌株生長完全被抑制;植物乳桿菌N8在培養基滲透壓超過800 mOsm/kg時生長速率被抑制,滲透壓達到3 000 mOsm/kg時菌株生長完全被抑制;植物乳桿菌N9在培養基滲透壓超過1 200 mOsm/kg時生長速率被抑制,滲透壓達到2 700 mOsm/kg時菌株生長完全被抑制。
植物乳桿菌培養過程中最適碳氮比為對數生長期生長速率被抑制時的碳氮消耗比,植物乳桿菌X1和植物乳桿菌N8最適碳氮比均為2.2 ∶1,植物乳桿菌N9最適碳氮比為3.0 ∶1,菌株最適碳氮比對培養工藝策略優化具有重要的指導意義。
植物乳桿菌的培養應控制發酵體系滲透壓在各自的可耐受滲透壓范圍內,植物乳桿菌X1最適培養策略:酵母浸粉FM803 40 g/L,葡萄糖15 g/L,MnSO40.25 g/L,恒pH 5.5恒溫37℃自動反饋補料36 mL葡萄糖溶液(質量濃度為400 g/L),活菌數提高約10倍,達到4.1×1010CFU/mL;植物乳桿菌N8最適培養策略為酵母浸粉FM528 40 g/L,葡萄糖88 g/L,MnSO40.25 g/L恒pH 5.5恒溫37℃分批培養,活菌數提高約5倍,達到2.9×1010CFU/mL。植物乳桿菌N9最適培養策略為酵母浸粉FM803 45 g/L,葡萄糖135 g/L,MnSO40.25 g/L,恒pH 6.0恒溫37℃分批培養,活菌數提高約12倍,達到6.2×1010CFU/mL。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
