長白山白樺山楊次生林細根形態特征和解剖結構對氮沉降的響應
2020-03-13 05:20:22洪梓明邢亞娟閆國永張軍輝王慶貴
生態學報 2020年2期
洪梓明,邢亞娟,閆國永,張軍輝,王慶貴
1 黑龍江大學農業資源與環境學院,黑龍江省寒地生態修復與資源利用重點實驗室,哈爾濱 150080 2 黑龍江省林業科學研究所,哈爾濱 150081 3 中國科學院沈陽應用生態研究所,沈陽 110016
一般認為,細根是指直徑≤2 mm的根系組分,而直徑>2 mm則為粗根。作為樹木養分吸收和水分運輸的重要器官,細根不斷地進行著呼吸作用與養分周轉,因此其在陸地生態系統碳(C)、氮(N)循環中發揮著重要作用[1- 2]。有研究顯示,在全球范圍內,如果細根每年周轉一次,約消耗陸地生態系統凈初級生產力(NPP)的33%[3]。細根作為樹木根系中最為活躍的部分,其養分和水分吸收直接影響著生態系統的能量流動和物質循環。在全球N沉降加劇背景下,外源N添加會對樹木根系生長產生復雜影響。對于細根長度,先前研究發現外源N添加可能促進[4]或抑制[5]細根生長。其中,Eissenstat和Yanai的最優化理論[6]認為N添加能增加細根長度以擴大與土壤的接觸面積,并能觸及更深層土壤以便充分吸收養分和水分來促進細根生長。而抑制細根生長的原因有兩方面:第一,當土壤N含量豐富時,就不需要消耗大量能量生產更長的細根去吸收養分[7];第二,N沉降降低了土壤pH值,使得土壤酸化[8],導致土壤中游離鋁離子(Al3+)濃度增加,可能抑制細根根尖分生組織細胞分裂,進而影響細根長度。陳海波等[9]通過研究水曲柳(Fraxinusmandshurica)苗木根系解剖結構時發現,N添加處理可能會導致單位長度上細胞數量減少,從而使根長縮短。除了細根長度,外源N添加也能夠促進細根直徑增加,其原因有二。首先,土壤中Al3+增加會促進細根直徑增粗[10];其次,由于土壤N含量增加,細根需要增加中柱直徑來促進養分和水分的運輸,與閆國永等[11]低N顯著促進興安落葉松(Larixgmelinii)前三級細根中柱直徑生長的研究結論一致。同時,谷加存等[12]研究結果顯示,無論是熱帶還是溫帶樹種,細根直徑受皮層厚度的影響較大,而受中柱直徑的影響較小。由于形態結構中的細根長度、直徑與解剖結構中的中柱直徑、皮層厚度具有緊密聯系,因此,本研究擬通過研究植物根系形態和解剖來確定N沉降對其地下過程的影響。
對于植物根系問題的研究一直處于不斷探索當中。賈淑霞等[13]通過研究興安落葉松與水曲柳細根時發現,形態結構與其生理功能緊密相連。形態特征是以往細根研究的主要內容,但其研究結果往往具有單一性。常文靜和郭大立[14]對我國溫帶、亞熱帶和熱帶45個樹種根系形態進行研究時發現,根序變化僅能解釋其直徑變異的53%;王向榮等[15]研究水曲柳前五級根時發現,根序僅能解釋直徑變異的83%。近年來,隨著細根研究的進一步深入,如果只從形態特征來探究其根系的生理功能已經不能滿足當前研究的需要。谷加存等[16]認為,細根直徑的變異主要來源于皮層厚度和維管束直徑的變化。陳海波等[8]研究水曲柳苗木根系時發現不同N濃度條件下根長度的變化可能與細胞數量的變化有關。植物形態變化決定其功能作用,而功能作用歸根到底是其生理生化的外在表現。因此,對于細根相應組分化學計量學的進一步研究才能解釋其外在形態與功能的相應變化。例如,McCormack等[17]研究發現,植物根系壽命與根系C∶N正相關。也有研究表明N∶P是植物生長養分限制的敏感性指標,可作為判斷生態系統健康與否的有效工具[18]。如“生長速率假說”認為:生物體為滿足快速生長階段的需要,將會投入更多富含P的rRNA來支持蛋白質的合成,rRNA作為植物的一個主要P庫,隨著含量增加將直接導致細胞中P濃度的增加,從而使植物表現出低的C∶N、C∶P和N∶P[19]。因此,在目前全球氣候變化的背景下,外源N的輸入可能改變細根化學組分的含量進而影響細根形態特征和生理功能。上述研究只考慮了某兩個或多個因子之間的耦合關系,并沒有系統性地論述細根形態特征、解剖結構、生理功能以及元素含量變化與外源N添加的耦合關系。目前,隨著細根研究的不斷深入,已經從單一的形態研究發展到形態特征、解剖結構、生理功能和元素含量耦合的系統性研究[20- 21]。
森林生態系統具有豐富的物種多樣性。有研究表明,每個樹種在遺傳、生活史、生境和物候以及生態功能方面均存在著明顯的差別[22]。相比于室內試驗,野外原位試驗更貼近N沉降現狀,能夠囊括土壤N有效性、植物根系自身特性、環境因素、土壤動物等因子的綜合作用。白樺與山楊都屬于長白山地區天然次生林典型先鋒樹種,均十分耐寒冷、瘠薄,天然更新能力強。但白樺較適宜生長在潮濕、偏酸性的土壤中,山楊喜干旱、偏中性的土壤。本試驗以長白山天然次生林白樺和山楊為研究對象,通過野外采集完整根系樣品,測定其前三級細根形態、解剖與C、N、P參數,并分別探究兩樹種各參數之間的相互耦合關系,以完善溫帶天然次生林細根對N沉降響應機制的研究。在前人研究的基礎上,我們提出以下假設:1)依據累積-穩定效應假說,長期施N可能抑制根系生長,增加細根直徑;2)由于根系直徑變異的控制因素不同,N添加下細根直徑增加可能是由中柱直徑增加引起;3)盡管白樺和山楊同為先鋒樹種,但由于白樺(喜微酸性土)和山楊(喜中性土)的生理差異,可能導致白樺細根隨著氮濃度梯度升高,呈現先促進后抑制生長的現象;而山楊細根隨著氮濃度梯度升高,呈現不斷抑制生長的現象。
1 材料與方法
1.1 試驗地點
試驗地點位于吉林省白山市撫松縣露水河林業局(42°82′49″ N,128°85′45″ E)。海拔920 m,屬典型季風性氣候,冬季寒冷干燥,春秋季短暫而干燥,夏季潮濕溫暖。年平均氣溫和降水量分別為2.78 ℃和871.6 mm。2004年至2007年該地平均大氣N沉降量為2.45 g N m-2a-1。其中濕N沉降約占75%。土壤為深棕色森林土壤,基巖為覆蓋著礫石和黃土的花崗巖,上層主要為黃土。屬于天然次生闊葉林,主要樹種有白樺(B.platyphylla)、山楊(P.davidiana)、加楊(P.koreana),伴有散生的水曲柳(F.mandschurica)、紅松(P.koraiensis)、色木槭(Acermono)、簇毛槭(A.barbinerve)、旱柳(Salixmatsudana)、楓樺(B.costata)、榆樹(Ulmusjaponica),平均樹齡約40年。主要灌木樹種是珍珠梅(Sorbariasorbifolia)、東北山梅花(Philadelphusschrenkii)、五味子(Schisandrachinensis)、溲疏(Deutziascabra)、衛矛(Euonymusalatus)。
1.2 試驗設計
控制試驗共設置了9個30 m×30 m的樣方,每個樣方相距20 m,并在底部用聚乙烯塑料板隔斷至土壤母質層(約60cm),以防止相鄰樣方不同N添加的相互影響。共設置3個N添加梯度,每個3次重復,即對照(CK)、低N(TL)、高N(TH),分別按0、2.5、5.0 g N m-2a-1進行外加N(NH4NO3)處理(不包括大氣N沉降量)。根據 N添加水平,于2006年5月起,在植物生長季內(5—9月)每月月初將每個樣方所施的NH4NO3首先溶解于20 L純凈水中,再混合于180 L水中,以背式噴霧器人工均勻噴灑在林地上。每個對照樣地則噴灑同樣200 L的純凈水,保持水分一致性。于2017年7月植物生長季在每個樣方內隨機選取白樺和山楊各3株,在距離樹干1m處用平板利鍬分別挖取白樺和山楊完整根段(包含前1—5級的根序)各27束,并依序放入寫好標簽的封口袋中,再置入冷藏箱(2—3℃)運回實驗室。將根系清洗后,放入裝滿FAA固定液((FAA:70%酒精(90 mL)+100%冰醋酸(5 mL)+38%甲醛(5 mL))的塑料瓶中進行固定,于4 ℃冰箱中保存待用。
1.3 形態測定與解剖學觀察
挑選完整的根段樣品,按照Fitter和Stickland[23]的方式對根進行分級,依序放入已標記好根序標簽且裝有FAA固定液的玻璃瓶內,在最短的時間內用CanonLide 120掃描儀完成圖像掃描。然后放入65度的烘箱內烘至恒重(48 h),測定各根序生物量(m干,g),密封于1號塑料袋內以便于化學含量測定。將掃描完成的圖像采用WinRHIZO軟件(加拿大Regent Instruments公司)進行形態特征分析,包括根長(L)、平均直徑(D)、表面積(S)。
采用上述方式用70%酒精分出一、二和三級根各30個。依序裝入標記好根序標簽且盛有70%酒精的玻璃瓶內,然后進行石蠟橫切片制作,每個處理等級根序制作7個樣本。用番紅-固綠染色(木質化細胞壁和細胞核染成紅色,薄而具有纖維素的細胞壁和細胞質染成綠色),加拿大樹膠封片。使用Leica DM4000B LED顯微鏡鏡檢,每個樣本觀察10個視野。并使用Leica Application Suite Version4.6.2軟件拍照和測量皮層厚度(d皮,μm)、皮層細胞層數(n層)、中柱直徑(d中,μm)和中柱內的導管數量(n導,個)與直徑(d導,μm),計算維根比(中柱直徑/根直徑)。
1.4 C、N、P含量測定
分別將封于1號塑料袋內兩樹種各處理的前三級根序依次混合。用TOC/TN分析儀(Multi N/C 3100,Analytikjene AG,Germany)灼燒測得樣品總有機碳(TOC)含量。用消煮法稱取0.03 g根系樣品制成溶液,用TOC/TN分析儀(Multi N/C 3100,Analytikjene AG,Germany)測得樣品總N(TN)含量。使用鉬銻抗比色法測定根系總P(TP)含量。
1.5 數據分析
采用單因素方差分析不同N添加對白樺和山楊細根形態、解剖和化學含量的影響,隨后通過LSD法確定不同水平N添加1—3級根形態特征、解剖結構和根系化學成分之間的差異(P<0.05)。數據統計和作圖采用SPSS 19.0(SPSS Inc,Chicago,USA)軟件完成。
2 結果與分析
2.1 不同水平N添加對細根化學組分的影響
由表1可見,白樺、山楊TOC含量在不同N水平下均無顯著差異。白樺TN含量在TL和TH均顯著升高(P<0.05,表1),在TL下增加了約19.68%,在TH下增加了約18.80%。而山楊僅TL出現了顯著降低(P<0.05,表1),降低約11.26%。TP含量在所有處理下都無顯著差異。
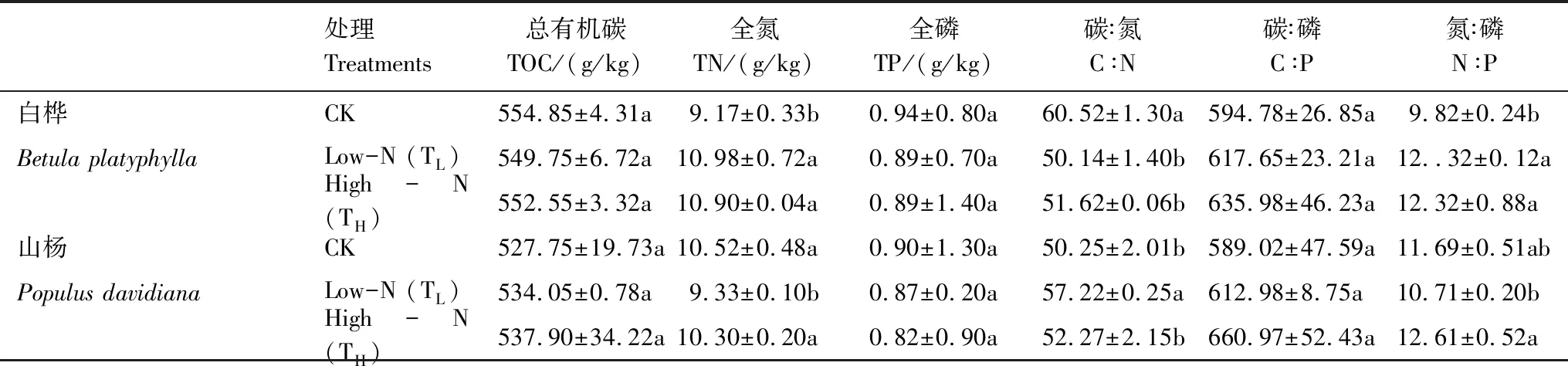
表1 N添加對白樺、山楊1—3級根C、N、P含量及其比值的影響
物種間的差異均不做比較;不同字母代表在P<0.05水平上的差異顯著
2.2 不同水平N添加對細根形態特征的影響
N添加顯著增加了白樺前三級根平均直徑(P<0.05),在TL時,一級根增加34.22%,二級根增加39.65%,三級根增加29.90%;在TH時,一級根增加13.36%,二級根增加35.10%,三級根增加14.58%。但TH相比于TL,細根直徑的增加量呈現降低趨勢,其中一、三級根顯著降低(P<0.05),一級根降低15.54%,三級根降低11.79%。前三級根平均長度在N添加下并沒有一致的變化趨勢。平均表面積在不同N添加下均顯著增加(P<0.05),在TL時,一級根增加33.09%,二級根增加55.56%,三級根增加110.96%;在TH時,一級根增加22.87%,二級根增加20.55%,三級根增加69.13%,而TH相比于TL均顯著降低(P<0.05),一級根降低7.68%,二級根降低22.50%,三級根降低19.81%。由此可見,在外源施N添加下,白樺通過改變細根直徑來影響其表面對營養物質的吸收過程以適應環境的變化(圖1)。
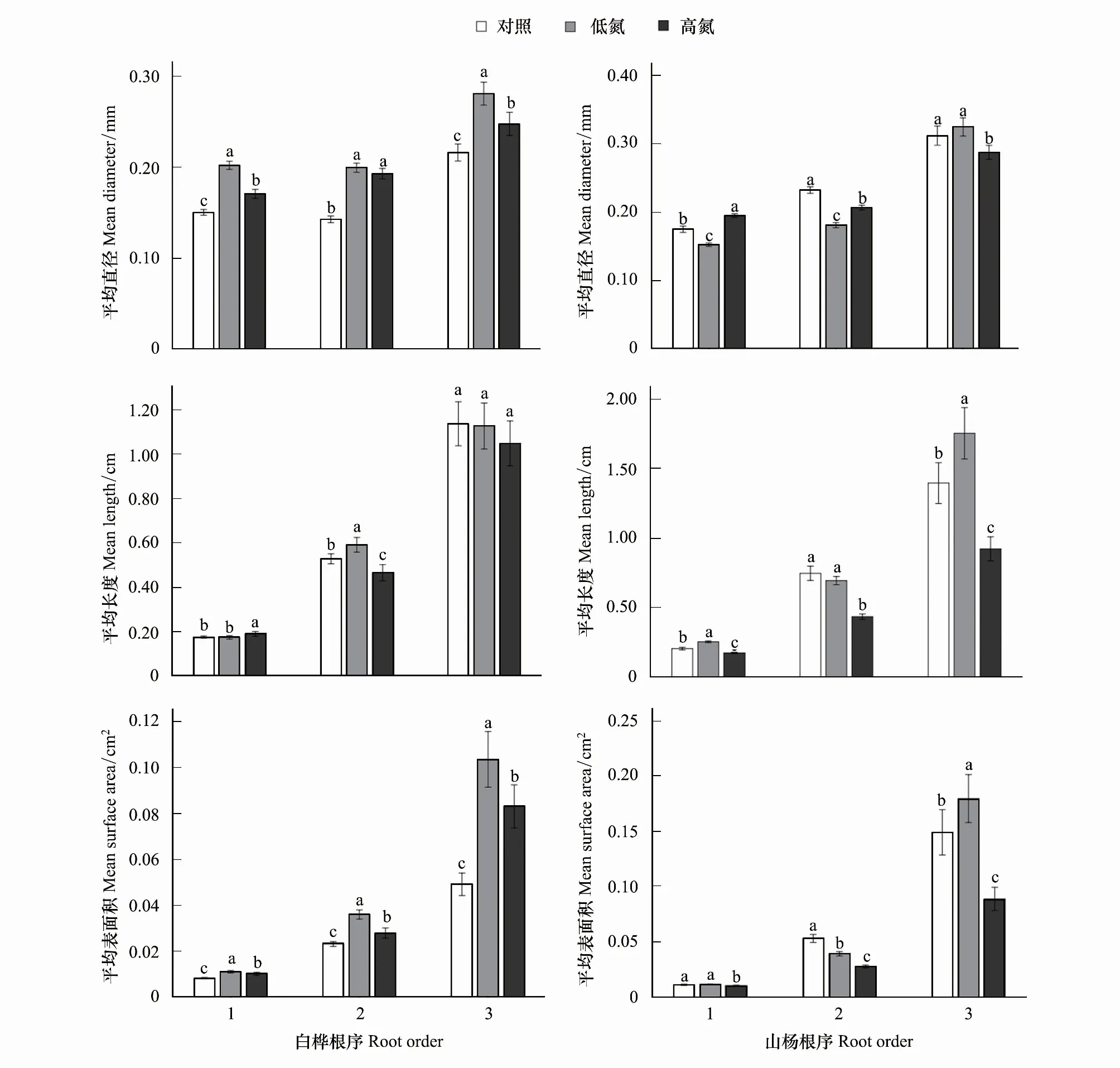
圖1 不同N水平白樺、山楊1—3級細根平均直徑、平均長度、平均表面積的變化Fig.1 Changes of average root diameter, average length and average surface area Betula platyphylla and Populus davidiana 1—3 fine roots in different nitrogen levels
山楊一、二級根平均直徑在TL時顯著降低(P<0.05),其中一級根降低12.96%,二級根降低22.18%,三級根無明顯變化;而一、二級根TH相比于TL顯著增加(P<0.05),三級根顯著降低(P<0.05),其中一級根增加28.06%,二級根增加14.27%,三級根降低11.46%。一、三級根平均長度在TL時顯著增加(P<0.05),其中一級根增加24.12%,三級根增加25.66%,二級根無顯著變化;而前三級根平均長度在TH下相比于CK和TL均顯著降低(P<0.05),相比于CK,一級根降低16.15%,二級根降低41.76%,三級根降低了33.91%;相比于TL,一級根降低32.44%,二級根降低37.36%,三級根降低47.41%。平均表面積TH相比于CK和TL均顯著降低(P<0.05)。相比于CK,一級根降低7.88%,二級根降低47.75%,三級根降低40.39%;相比于TL,一級根降低11.57%,二級根降低29.35%,三級根降低50.51%,而在TL下,一級根無顯著變化,二級根顯著降低26.05%(P<0.05);三級根顯著增加20.44%(P<0.05)。與白樺不同,山楊在外源N添加下其長度和直徑均作出積極響應以適宜其生長環境的變化(圖1)。
2.3 不同水平N添加對細根解剖結構的影響
白樺前三級根皮層細胞直徑在N添加下均顯著增加(P<0.05),在TL時,一級根增加72.77%,二級根增加53.22%,三級根增加39.96%;在TH下,一級根增加57.74%,二級根增加46.33%,三級根增加30.16%。白樺皮層厚度在TL時,一級根無顯著變化,二、三級根均顯著降低(P<0.05),二級根降低13.21%,三級根降低25.41%。白樺中柱直徑在N添加時均顯著增加(P<0.05),在TL時,一級根增加54.26%,二級根增加38.32%,三級根增加49.64%,在TH時,一級根增加27.10%,二級根增加14.96%,三級根增加18.78%。而TH相較于TL均顯著降低(P<0.05),一級根降低17.61%,二級根降低16.89%,三級根降低20.62%。白樺維根比在N添加時均顯著增加(P<0.05),在TL時,一級根增加23.02%,二級根增加20.39%,三級根增加16.70%;在TH時,一級根增加16.97%,二級根增加5.79%,三級根增加9.02%。此外,TH相比于TL均顯著降低(P<0.05),一級根降低4.91%,二級根降低12.13%,三級根降低6.59%。由此前白樺細根直徑的變化可知,其主要通過皮層厚度與中柱直徑的協同作用來促使細根直徑發生改變(圖2)。
山楊前三級根皮層細胞直徑在TL時僅一、二級根顯著下降(P<0.05),一級根下降40.80%,二級根下降28.17%,三級根無顯著變化;而TH時,二、三級根均顯著增加(P<0.05),二級根增加17.37%,三級根增加103.45%;TH相比于TL均顯著增加(P<0.05),一級根增加76.54%,二級根增加63.39%,三級根增加139.27%。山楊皮層厚度在TL時,僅一、二級根顯著下降(P<0.05),一級根降低32.45%,二級根降低33.06%,三級根無顯著變化;在TH時,一、三級根顯著增加(P<0.05),一級根增加14.02%,三級根增加110.58%;TH相比于TL均顯著增加(P<0.05),一級根增加68.78%,二級根增加50.81%,三級根增加88.53%。山楊中柱直徑在TH時顯著下降(P<0.05),相比于CK,二級根下降12.01%,三級根下降41.08%;相比于TL,一級根下降11.81%,三級根下降41.37%。山楊維根比在TL時,一、二級根顯著增加(P<0.05),一級根增加46.02%,二級根增加14.67%,三級根無顯著變化;TH相比于TL均顯著降低(P<0.05),其中一級根降低30.58%,二級根降低13.31%,三級根降低31.99%。結合此前山楊細根直徑的變化可知,山楊主要通過改變皮層細胞直徑來促使其皮層厚度發生變化以適應外源N添加改變(圖2)。
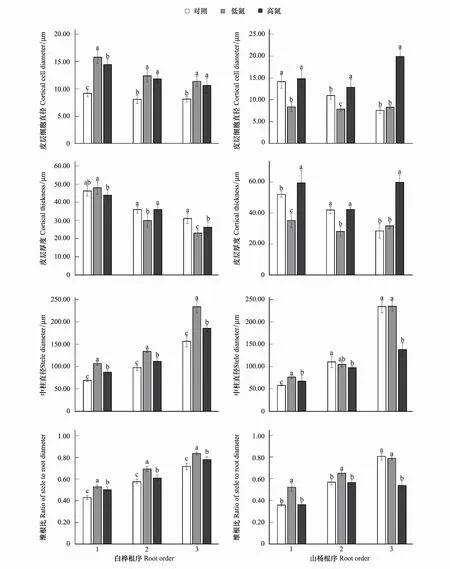
圖2 不同N水平白樺、山楊1—3級細根皮層細胞直徑、皮層厚度、中柱直徑、維根比的變化Fig.2 Changes of cortical cell diameter, cortical thickness, stele diameter, ratio of stele to root diameter of fine roots of Betula platyphylla and Populus davidiana 1—3 fine roots in different nitrogen levels
白樺導管個數和導管平均直徑在TL時均顯著增加(P<0.05),導管個數增加130%,導管平均直徑增加25.05%;TH相比于TL均顯著降低(P<0.05),導管個數降低60.14%,導管平均直徑降低17.85%。
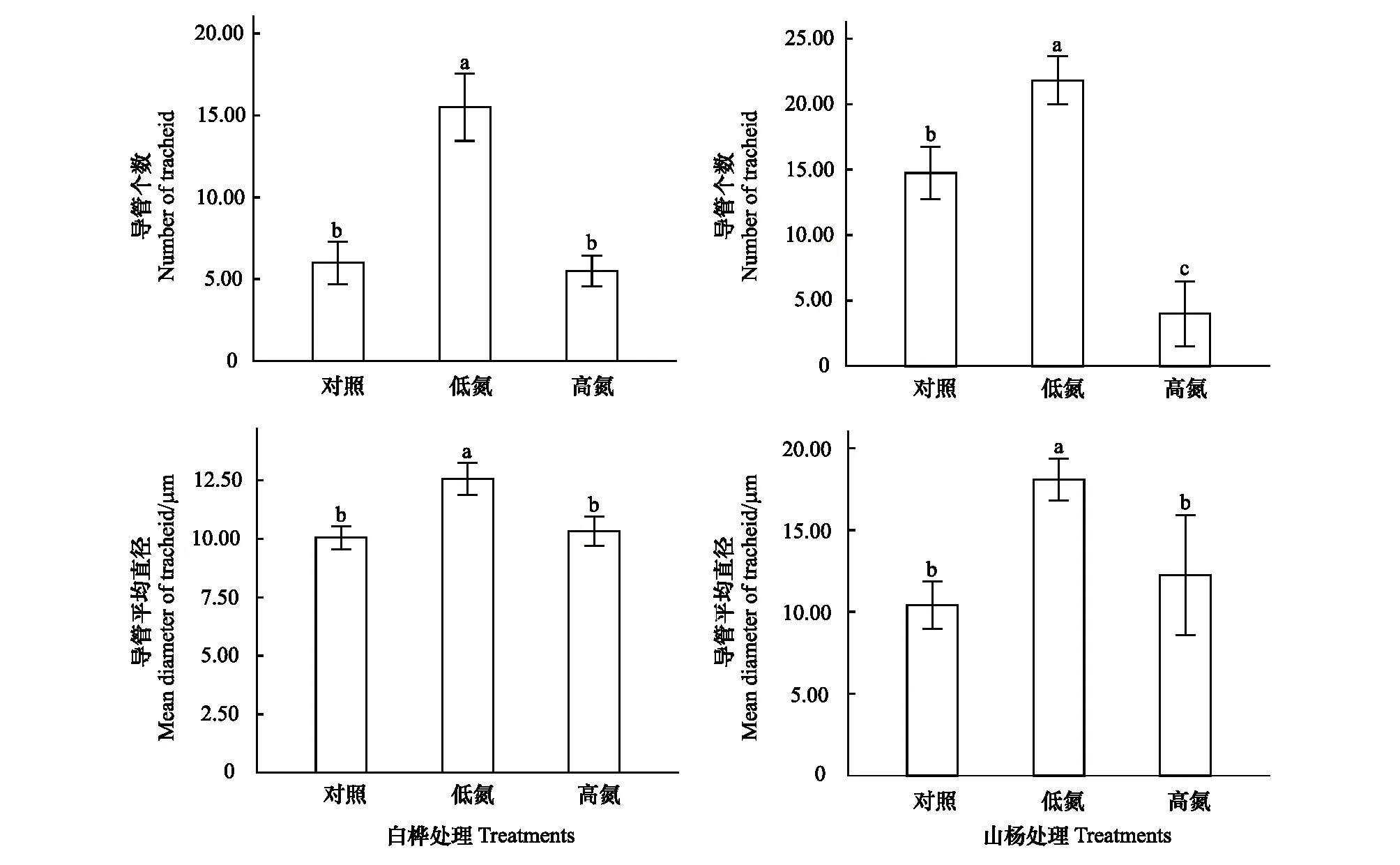
圖3 不同N水平白樺、山楊3級細根導管個數和導管平均直徑的變化Fig.3 Changes of mean diameter of grade 3 fine roots in Betula platyphylla and Populus davidiana in different nitrogen levels
山楊導管個數和導管平均直徑在TL時顯著增加(P<0.05),其中導管個數增加47.92%,導管平均直徑增加73.60%。在TH時,山楊導管個數相較于CK和TL均顯著降低(P<0.05),相較于CK降低72.88%,相較于TL降低81.67%;而導管平均直徑相較于CK無顯著變化,但相較于TL顯著降低32.27%(P<0.05)(圖3)。
3 討論
樹木的形態結構與其生理功能具有緊密聯系[24],而N沉降能顯著影響植物細根的生存策略[25]。先前研究主要將根系分為粗根(直徑>2 mm)和細根(直徑≤2 mm)兩部分[26],認為細根主要起水分和養分吸收的功能,而粗根起儲藏、運輸、支撐的作用,但目前有研究認為這種結論存在一定的不足。近期研究顯示,假如采用直徑大小對細根進行劃分,將會在某種程度上忽視個體根在整個根序上的差別。由于不同根序上的個體根在解剖、形態和生理功能上都存在明顯的差異性,因而采用直徑閾值來對根系組分進行劃分,缺乏對根系功能方面的充分考慮,這將會阻礙對細根結構和功能的準確理解[17,27- 29]。谷加存等[16]認為由于1—3級根都具有初生構造和完整的皮層組織結構,主要起水分和養分吸收的功能,因此被定義為吸收根,并建議采用功能劃分的方法來充分判定樹木細根系統內結構和功能的異質性,進而更加客觀地評價根系所蘊含的生態學過程及其影響。通過以上觀點,本研究分析了不同N添加對白樺和山楊1—3級細根的形態特征和解剖結構的影響,并探討了與元素含量之間的耦合關系。
3.1 N沉降對細根C、N、P元素的影響
本研究表明外源N輸入顯著改變了細根N含量,但并沒有明顯改變C、P含量(表1)。進一步分析表明:白樺細根C∶N在TL和TH均顯著降低,其原因在于外源N輸入促進細根呼吸[30],使細根周轉加快,細根N含量升高,導致細根C∶N減小。白樺和山楊C∶P隨著N濃度增加均無顯著變化,說明土壤P含量并沒有隨著外源N的輸入而處于緊缺狀態,符合前人的研究結論:在寒冷的氣候下,由于生長季較短,物種生長速率較快,溫帶物種組織中的P含量比熱帶物種豐富[31]。此外,Koerselman和Meuleman[32]通過對濕地植被施肥試驗的研究表明,N∶P<14可以認為是受N限制,N∶P>16是受P限制。本實驗中N∶P均小于14,沒有受到P元素限制。此外,白樺和山楊C∶P與N∶P隨著N濃度升高成增加趨勢,與“生長速率假說”認為植物應表現出低的C∶P和N∶P[19]不相符。但有學者研究發現,高等植物能夠儲存額外的養分從而改變生長速率和N∶P之間的關系[19,33- 35],本研究恰好驗證了這一觀點。
3.2 N添加下細根形態特征與解剖結構的耦合關系
目前有大量研究顯示,在溫帶森林中,土壤N的可用性能夠顯著影響細根的形態特征[36- 38]。許多研究發現通過外源添加N顯著改變了細根的長度[4- 5]和直徑[10,14]。本實驗中白樺細根在N添加下平均直徑均顯著增加(符合我們的假設1),山楊細根在N添加下平均直徑和平均長度均發生顯著變化(圖1)。物種間變化趨勢的不同可能反映了不同樹種的營養需求不一致,也可能反映了樹種間為適應變化的環境而采取不同的營養覓食策略[39- 40]。此外,細根的形態特征與其解剖結構有著十分緊密的聯系。一般認為,皮層厚度的降低使得營養物質進入中柱運輸部位的距離縮短[41],而導管平均直徑和個數顯著增加引起的中柱直徑增粗[28]使得皮層吸收的營養物質能夠更有效地進行運輸,進而改變了細根的形態結構。本實驗中白樺皮層厚度在N添加下呈現出下降的趨勢(圖2),中柱直徑在施N處理下均顯著增加(符合我們的假設2)。顯然,白樺通過增加中柱直徑和降低皮層厚度來提高維根比,從而滿足外源N添加處理后的生長促進狀態。與谷加存等[12]的研究結論:根徑受皮層厚度的影響較大,而受中柱直徑的影響較小不一致,其主要原因是谷加存等關注物種之間比較,并未關注施加外源N后其動態變化。另外,在木本植物中已經發現了根直徑與中柱大小的正相關關系[11,42]。白樺在TH與TL比較時,中柱直徑的變化與細根直徑的變化相契合(顯著降低),而二、三級根皮層厚度顯著增加。由此,我們推測白樺在TH時處于“營養過剩”狀態。山楊在施N處理下皮層厚度與中柱直徑無一致變化趨勢。白樺、山楊次生林細根在不同N添加下形態特征和解剖結構的適應性仍需進一步研究(圖2)。
一般而言,在吸收根中皮層厚度的變化可以用來評估植物適應變化的環境[43],而皮層厚度的變異來源有皮層細胞直徑和皮層層數兩個方面。本實驗中白樺皮層厚度在TL時顯著降低(一級根除外),而皮層細胞直徑顯著增加。可見,TL通過減少皮層層數來促進白樺皮層厚度的降低使得營養和水分吸收效率增加,促進細根生長。而山楊在TH相較于TL時,皮層厚度與皮層細胞直徑均顯著增加,在TL時又具有相同的變化趨勢(圖2)。由此可見,山楊主要通過影響皮層細胞直徑來改變皮層厚度。
3.3 N添加對細根功能的影響
有研究證實,吸收根可以通過改變根形態和解剖特征來優化其功能,以適應土壤條件諸如水分[7,44- 45]和重金屬含量[46- 48]的變化。在根直徑方面,先前很多研究[49- 51]認為:N供應不足,會對葉片CO2同化產生影響,導致分配到根系中的光合產物減少,從而增加細根衰老和死亡的概率,而直徑增粗可以降低這種反應的危險系數。本研究中山楊一、二級根平均直徑在TL時(N供應充足)顯著降低,三級根有增加趨勢,這充分驗證了上述研究結論。在根長度方面,許多研究表明根長度是體現根吸收效率的重要指標[52- 53]。山楊在TL時平均長度顯著增加以充分吸收養分,在TH時又顯著降低從而減小養分吸收速率[6](圖1)。可見,植物能夠通過調節自身形態以適應外界的環境變化。
還有部分研究顯示導管與細根運輸能力顯著相關。Senock和Leuschner[54]觀察到樹液流速和柳葉桉根部的最大液流速率分別與平均導管直徑和總導管橫截面積成正相關。在鱷梨根部也驗證了液流速率與總導管橫截面積的正相關性[55]。本實驗中兩樹種三級細根在TL時導管個數和導管平均直徑(導管橫截面積)均顯著增加(圖3)。表明了導管的形成能夠積極感受土壤環境中N含量的可用性。有實驗證實,在N含量較高的情況下植物會增加他們的運輸能力[43]去平衡植物吸收和運輸之間的關系[56- 57],避免營養物質積累在根系系統中[58]。另一方面,本實驗中白樺中柱直徑(圖2)符合其導管的變化趨勢,顯然,其中柱直徑的增加主要來源于導管的直接作用。山楊在TH時中柱直徑的變化符合其導管變化規律。在TL時中柱直徑無顯著變化,但導管平均直徑和導管個數均顯著增加。我們猜測TL使得山楊不再需要大量的韌皮部來積極運輸有機物和礦質元素離子,反而需要更多的導管來支持植物體內不需要能量的水分運輸的充分需要。然而,具體變化原因還需進一步探索。
此外,還發現一個有趣的現象。山楊皮層厚度在TH相較于TL時顯著增加(圖2),而導管個數和導管平均直徑(導管橫截面積)顯著降低(圖3)。同時,其平均表面積也相應地顯著降低(圖1),根據營養吸收-運輸假設[43],推測,山楊在TH時明顯感受到了環境的改變所產生的脅迫壓力,因此,其改變了自身的營養吸收和轉運策略來適應當前環境變化[39- 40,56- 57],以降低其破壞力。山楊皮層厚度的改變恰好驗證了我們對其全N的實驗結果:TL時為顯著低值(表1)。這符合植物組織解剖研究結果[59]:皮層作為初生組織,主要由薄壁細胞構成。其特點是養分濃度高(大部分是以蛋白質形式存在的N),代謝速率快(主要是通過離子吸收、同化及轉運到其他組織)(圖3,圖4)。
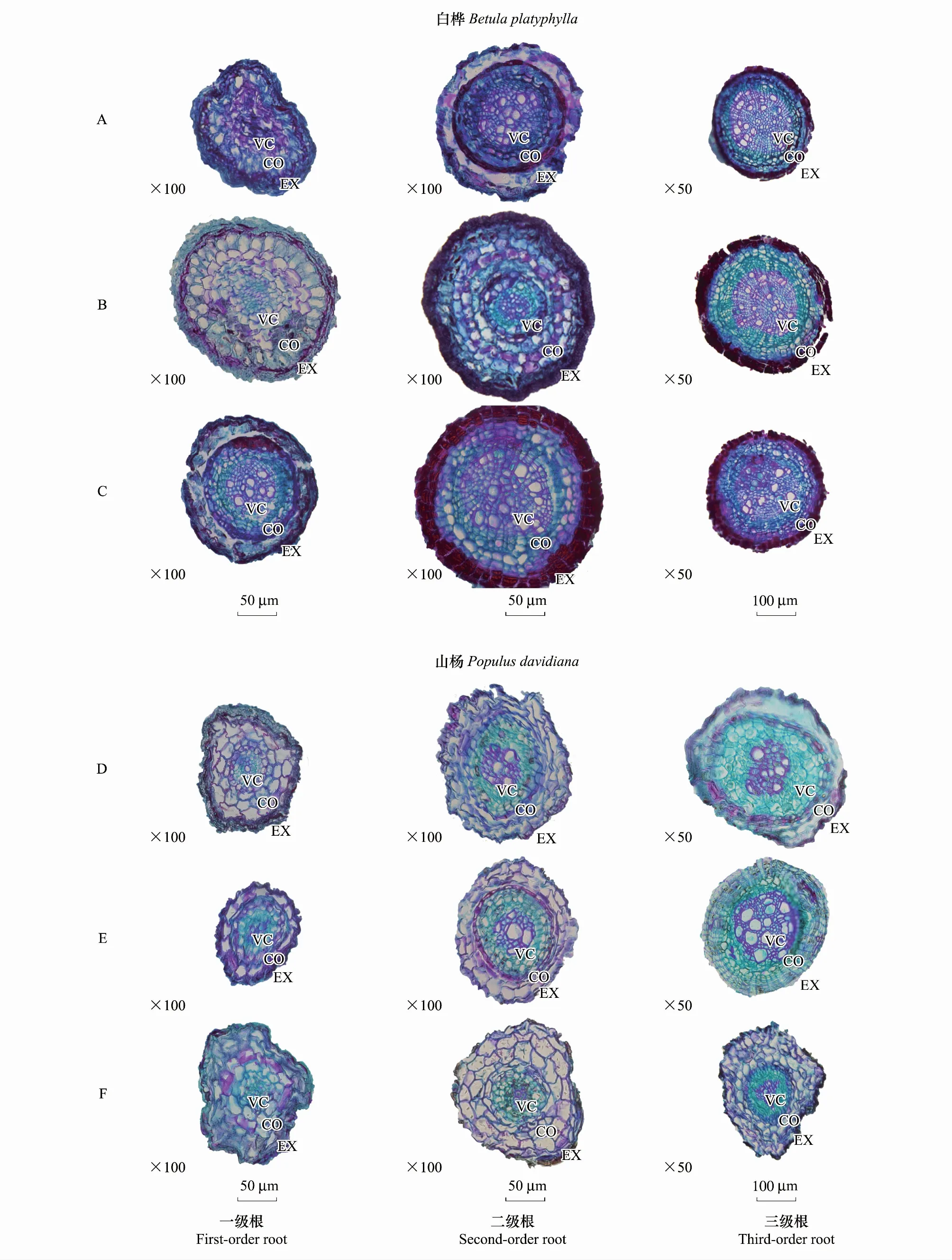
圖4 不同N水平白樺(A-CK、B-LN、C-HN)、山楊(D-CK、E-LN、F-HN)1—3級細根解剖結構圖片Fig.4 Anatomy and structure picture of Betula platyphylla (A-CK, B-LN, C-HN) and Populus davidiana (D-CK, E-LN, F-HN) 1—3 fine roots in different nitrogen levelsVC,中柱;CO,皮層;EX,外皮
白樺、山楊都屬于先鋒樹種,均十分耐寒。白樺通常生長于濕潤并呈酸性的土壤環境中,山楊偏愛排水良好且呈中性的土壤。由于人為添加N,改變了白樺和山楊細根的生存環境,使得其性狀發生改變。鑒于白樺自身的喜酸性特征,其在TL時長度無明顯變化而直徑顯著增加,可能原因是酸性土壤中Al3+濃度增加進而抑制細根根尖分生組織細胞分裂[8,10]。在TH時土壤酸性進一步增強,Al3+濃度過高,產生了鋁毒效應,影響其生長,因此呈現出TH直徑增加量明顯小于TL且長度逐漸降低的現象。相反,由于山楊呈現出較為“中性”的特征,在TL時山楊直徑生長受到抑制,而長度顯著增加,土壤中Al3+濃度效果不顯著,其原因可能是白樺作為“緩沖樹種”,在TL時生長加快而吸收了大量人為添加的N,進而調節土壤pH值,使其酸性減弱。然而在TH時,白樺自身生長也受到抑制,從而導致“緩沖效應”消失,因此鋁毒效應顯著,致使山楊細根長度顯著下降而皮層厚度顯著增加(符合我們的假設3)。據此,隨著目前全球氣候變化背景下的N沉降進一步延續,長白山地區白樺山楊天然次生林會持續展現出其自身的適應機制,從而為完善植物根系的響應研究提供數據支持。
4 結論
本研究對長白山試驗樣地內白樺和山楊兩樹種前三級細根進行形態、解剖和化學組分測定后,得到結論如下:白樺和山楊的細根對N沉降的反饋機制不同,白樺主要通過細根中柱直徑和皮層厚度來體現,而山楊通過細根長度和皮層厚度兩方面來體現;TL時,兩物種都是通過抑制前三級細根皮層厚度生長來加速水分與營養物質的吸收,不同之處在于白樺通過增加皮層細胞直徑但減少皮層層數來達到皮層厚度的降低,而山楊主要是通過削弱皮層細胞直徑的生長而促使皮層厚度的降低;TH時,山楊細根生長受到抑制,只能通過增加皮層厚度降低導管橫截面積來抵御過高的N濃度對其自身生長的危害。因此,植物能夠通過感知潛在的外界環境變化而改變其根系解剖結構,從而影響根系外在的形態特征進而優化其生理功能。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
