血管活性腸肽在單眼視覺剝奪性弱視幼貓外側膝狀體中的表達*
2020-03-11 09:13:30
中國現代醫學雜志 2020年4期
(川北醫學院附屬醫院 眼科,四川 南充 637100)
弱視是造成全世界兒童視力下降的主要疾病之一。近些年來,隨著分子生物學、神經生物學等多學科在弱視研究上的廣泛應用,其發病機制得到進一步揭示。目前,已知弱視的發生、發展主要與視覺神經系統(主要包括視網膜、外側膝狀體及視皮層)的異常改變緊密相關。弱視患者的視網膜和視皮層中多種神經遞質的含量改變,如神經生長因子、腦源性神經營養因子、營養神經素及多巴胺等。血管活性腸肽(vasoactive intestinal polypeptide,VIP)作為一種神經遞質,由28個氨基酸殘基組成,屬于分泌素-胰高血糖素家族,廣泛分布于中樞神經和腸神經系統中,隨后被發現也存在外側膝狀體中[1]。早前有研究表明,視覺剝奪性弱視幼貓視皮層17區的VIP 含量減少,但在外側膝狀體中的變化還未探究[2]。本實驗通過觀察視覺剝奪性弱視幼貓外側膝狀體中VIP的表達,探討VIP 對視覺發育的作用。
1 材料與方法
1.1 材料
1.1.1 實驗動物 健康3周齡幼貓30只(由川北醫學院實驗動物中心提供),雌雄、毛色不分,體重約290 ~360 g。排除屈光介質混濁及眼底異常,經檢測所有幼貓屈光度約+1.0 ~+2.0D。飼養在光照充足環境中,室內溫度維持在(27±1)℃。5周齡前幼貓不能自主攝取固態食物,每日定時喂養貓奶粉、飲用水共8 次。5周齡后保持室內存放充足食物及飲用水。本研究通過川北醫學院實驗動物倫理委員會批準并接受全程監督。
1.1.2 主要試劑及儀器 檸檬酸(pH=6.0)抗原修復液(武漢Servicebio公司),VIP抗體(美國Fitzgerald公司),辣根過氧化物酶標記的山羊抗兔抗體(武漢Servicebio公司),免疫組織化學試劑盒DAB 顯色劑(武漢Servicebio公司),焦碳酸二乙酯(美國Amresco公司),兔抗地高辛抗體(美國Jackson公司),探針序列,正向引物:5’-DIGTGCATCCGAGTGGCGCTTGATTGG-DIG-3’;反向引物:5’-DIG-CTGGTTTCCATCTTTGTACC TTGCCAAGTAGTG-DIG-3’(武 漢Servicebio 公 司),動物針電極(北京高視遠望科技有限責任公司),視覺電生理儀(重慶國特醫療設備有限公司),掌上離心機(武漢Servicebio公司),原位雜交用離心管(美國Axygen公司),成像系統(日本Nikon公司)。
1.2 方法
1.2.1 動物模型復制 30只幼貓隨機分為對照組和剝奪組,每組15只。用戊巴比妥鈉溶液腹腔注射麻醉剝奪組幼貓后,于右眼眶周邊對稱做4個皮膚固定縫線,每個縫線上做1個小套環(遮蓋過程中套環一直存在),使用絲線穿過小套環,再穿過黑色眼罩的4個小孔,打結固定眼罩,保證眼罩不壓迫眼球。每周剪斷絲線行圖形視覺誘發電位(pattern visual evoked potential,PVEP)檢測。對所有幼貓行PVEP 檢測時,3 根動物針電極分別刺入前額正中,雙耳連線枕部正中及耳尖背部的皮下。相應鏡片矯正屈光,調整頭位使視網膜后極部中央與屏幕中央在同一水平線上,PVEP 采用棋盤格反轉刺激,模式選用0.3 cpd,時間頻率為1 Hz,疊加64 次,分別測量剝奪組剝奪眼和未剝奪眼、對照組右眼。6周齡時,通過比較PVEP的P100波潛伏期及振幅,證實視覺剝奪組中單眼弱視形成。用戊巴比妥鈉深度麻醉剝奪組和對照組幼貓,分離出右側外側膝狀體,石蠟包埋切片,行免疫組織化學及原位雜交實驗研究。
1.2.2 實驗方法 免疫組織化學:石蠟切片,常規脫蠟,抗原修復,阻斷內源性過氧化物酶,血清封閉,分別加入一抗、二抗,DAB 顯色,復染細胞核,脫水封片,顯微鏡鏡檢,圖像采集分析陽性細胞和陽性細胞平均光密度值。原位雜交:石蠟切片,常規脫蠟,消化,阻斷內源性過氧化物酶,預雜交,雜交,雜交后洗滌,滴加封閉液,滴加兔抗地高辛抗體,DAB 顯色,復染細胞核,脫水封片,顯微鏡檢,圖像采集分析陽性細胞和陽性細胞平均光密度值。
1.3 統計學方法
數據分析采用SPSS 22.0統計軟件。計量資料以均數±標準差(±s)表示,比較用t檢驗或方差分析,進一步的兩兩比較用LSD-t檢驗,P<0.05 為差異有統計學意義。
2 結果
2.1 PVEP 結果
對照組幼貓隨著年齡增加,P100波潛伏期逐漸縮短,振幅逐漸提高。測試時控制電阻,可見各組典型PVEP 波形(見圖1)。6周齡時,對照組右眼、剝奪組右眼和剝奪組左眼潛伏期比較,經方差分析,差異有統計學意義(P<0.05);剝奪組右眼較對照組右眼及剝奪組左眼的潛伏期延長(P<0.05)。對照組右眼、剝奪組右眼和剝奪組左眼振幅比較,經方差分析,差異有統計學意義(P<0.05),剝奪組右眼較對照組右眼及剝奪組左眼振幅降低(P<0.05)。在進行單眼遮蓋3周后(即6周齡時),剝奪組幼貓右眼已形成單眼視覺剝奪性弱視。見表1。
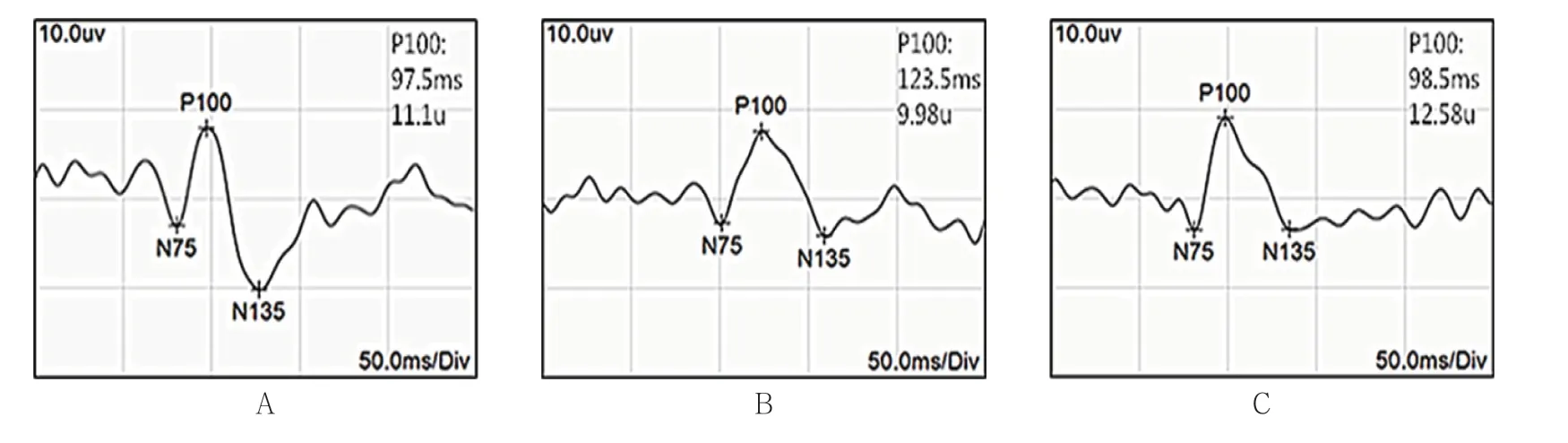
圖1 各組幼貓PVEP 波形圖
表1 各組P100 波潛伏期和振幅比較(n =15,±s)
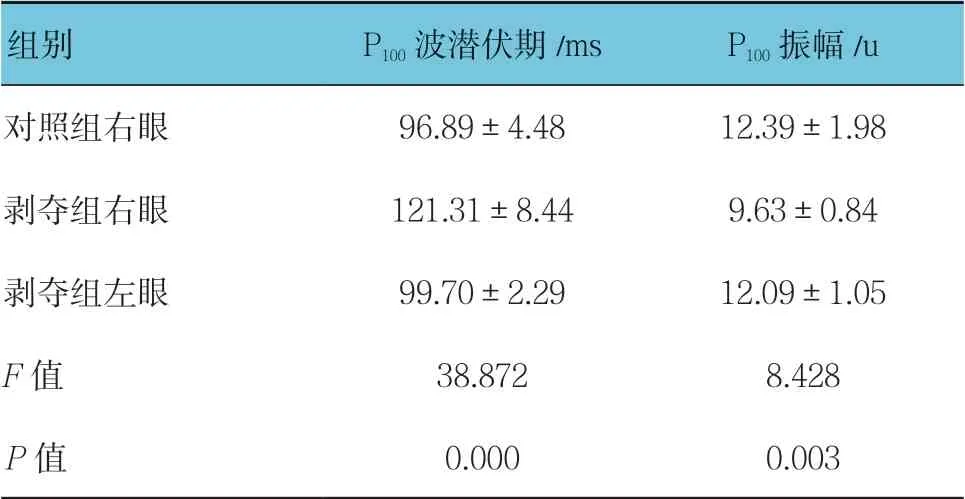
表1 各組P100 波潛伏期和振幅比較(n =15,±s)
組別 P100 波潛伏期/ms P100 振幅/u對照組右眼 96.89±4.48 12.39±1.98剝奪組右眼 121.31±8.44 9.63±0.84剝奪組左眼 99.70±2.29 12.09±1.05 F 值 38.872 8.428 P 值 0.000 0.003
2.2 免疫組織化學結果
對每張切片隨機選擇5個視野進行統計分析。可見兩組幼貓外側膝狀體神經元中VIP 呈陽性表達,位于細胞漿內,呈棕黃色-淡黃色,細胞核呈藍色。對照組大部分呈強陽性,陽性細胞數較多,剝奪組大部分呈中等陽性到弱陽性,陽性細胞數較少,可見兩組VIP 蛋白表達存在差異(見圖2)。兩組陽性細胞數比較,經t檢驗,差異有統計學意義(P<0.05),剝奪組較對照組少。兩組陽性細胞平均光密度值比較,經t檢驗,差異有統計學意義(P<0.05),剝奪組較對照組低。視覺剝奪性弱視幼貓外側膝狀體中VIP 蛋白表達較正常幼貓降低。見表2。
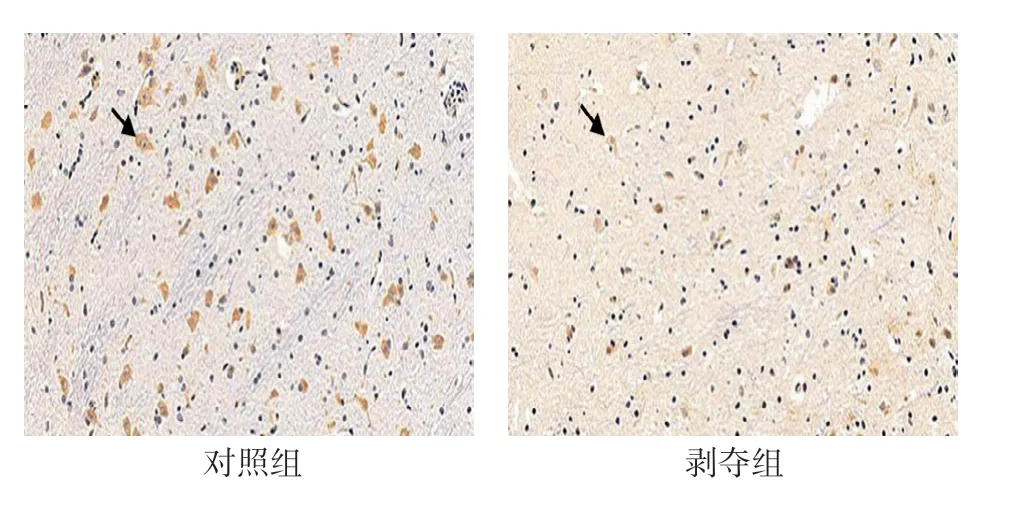
圖2 6周齡外側膝狀體神經元中VIP 蛋白的表達(DAB×200)
2.3 原位雜交結果
對每張切片隨機選擇5個視野進行統計分析。兩組幼貓外側膝狀體神經元中均有VIP mRNA 陽性表達,位于細胞漿內,呈棕黃色-淡黃色,同時與藍染的細胞核重疊。對照組大部分呈強陽性,陽性細胞數較多,剝奪組大部分呈中等陽性到弱陽性,陽性細胞數較少,可見兩組VIP mRNA表達存在差異(見圖3)。兩組陽性細胞數比較,經t檢驗,差異有統計學意義(P<0.05),剝奪組較對照組少。兩組平均光密度值比較,經t檢驗,差異有統計學意義(P<0.05),剝奪組較對照組低見表3。視覺剝奪性弱視幼貓外側膝狀體中VIPmRNA 轉錄水平較正常幼貓下降。見表3。
表2 兩組外側膝狀體中VIP 免疫組織化學結果比較(n =15,±s)
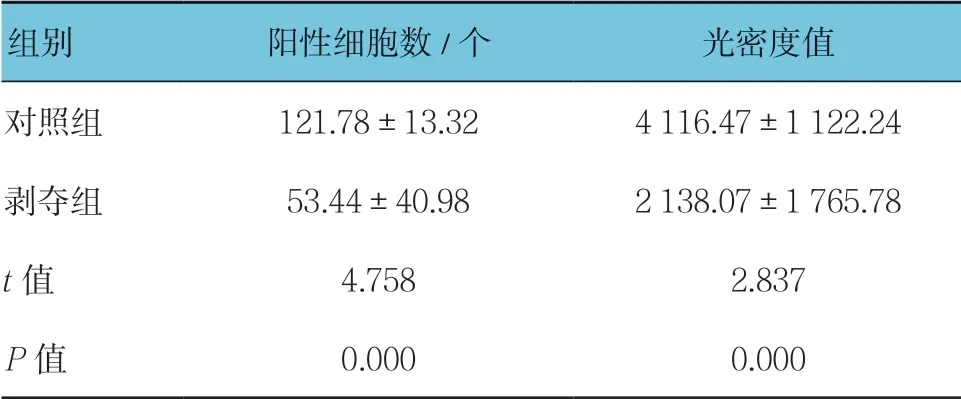
表2 兩組外側膝狀體中VIP 免疫組織化學結果比較(n =15,±s)
組別 陽性細胞數/個 光密度值對照組 121.78±13.32 4 116.47±1 122.24剝奪組 53.44±40.98 2 138.07±1 765.78 t 值 4.758 2.837 P 值 0.000 0.000
圖3 6周齡外側膝狀體神經元中VIP mRNA的表達(DAB×200)
表3 兩組外側膝狀體中VIP 原位雜交結果比較(n =15,±s)
表3 兩組外側膝狀體中VIP 原位雜交結果比較(n =15,±s)
組別 陽性細胞數/個 光密度值對照組 61.00±20.30 3 061.76±1 595.64剝奪組 35.00±13.40 1 307.08±623.24 t 值 3.024 2.897 P 值 0.009 0.012
3 討論
本實驗通過PEVP來確定剝奪組弱視的形成,然后用免疫組織化學和原位雜交來檢測VIP在外側膝狀體中的表達變化。通過實驗發現,幼貓在視覺發育敏感期內,由于雙眼視覺信息的不對等輸入,導致外側膝狀體中VIP表達降低;而VIP表達下降又對外側膝狀體正常表達的生理功能造成負面影響,從而影響視覺發育,促進弱視的發生、發展。
VIP 最早于1970年由豬小腸的甲醇抽提液中經過分離得到,其具有血管舒張活性,最初被認為是一種候選胃腸激素。后來發現其廣泛分布于大腦皮質、眼內組織中[3-6],隨后證明在外側膝狀體中也廣泛存在[1]。VIP 主要作為神經遞質起作用[7],能夠促進神經細胞的有絲分裂,增加神經細胞突起的數量[8];同時促進前體細胞向神經元分化,提高成熟神經元的數量[9]。VIP 還影響血漿中胰多肽、脂聯素及胰島素等代謝調節激素的表達水平,進而影響細胞代謝[10]。以往學者認為,外側膝狀體僅僅作為視網膜神經傳遞的接受體,將來自視網膜的神經纖維束換能后傳遞至視皮層,其并不參與視覺功能的編碼,視皮層才是弱視主要的神經系統受損部位。但后來發現外側膝狀體并不是單純的神經傳導中繼站,其參與方向性等精細視覺的形成[11]。在單眼剝奪性弱視中,與剝奪眼同側的外側膝狀體被觀察到存在神經元活動減弱[12]和神經元萎縮[13]的現象,同時經弱視側的外側膝狀體投射到視皮層17區、18區的神經元體積也減小[14]。而且單眼剝奪性弱視中,剝奪側的外側膝狀體中細胞大小的改變與同側視皮層第4 層中眼優勢柱寬度的減少密切相關[15]。有研究發現,外側膝狀體與視皮層一樣,在成年動物中仍然具有經驗依賴的可塑性,其部分功能可以在弱視后得以恢復,提示外側膝狀體在弱視治療中也應得到重點關注[16-18]。可見外側膝狀體在視覺神經系統的發育過程中不僅有神經傳導作用,還有功能編輯作用。研究異常視覺體驗對外側膝狀體中VIP表達的影響可以論證VIP在視覺發育中扮演的重要地位。
對于幼貓,視覺剝奪的影響程度在出生后1個月達到峰值,到3個月齡時,視覺剝奪對視覺發育的影響幾乎為零[19]。本實驗在視覺發育關鍵期內進行單眼視覺剝奪,通過PVEP 檢測弱視的形成,這與目前普遍認為的弱視形成機制和診斷方式相一致。對照組右眼、剝奪組左眼與剝奪組右眼的P100波潛伏期和振幅比較有差異,說明實驗組單眼弱視已形成,其中P100波潛伏期延長更能提示視路的傳導存在問題;同時創新性的遮蓋方式降低因傳統眼瞼縫合而增加皮損處感染的風險,避免定期PVEP 檢測對眼瞼皮膚的反復縫合造成諸如角膜刺激、縫線斷裂等影響測試的不良后果。
本文分別從蛋白和基因層面證實視覺剝奪對外側膝狀體中VIP表達的影響。免疫組織化學結果表明,在幼貓視覺發育敏感期內的視覺剝奪,使外側膝狀體接受視網膜的視覺傳入信號減弱,致使外側膝狀體中表達VIP的神經元得不到足夠的視覺刺激信號,導致神經元的功能發育異常,蛋白表達能力降低。原位雜交實驗結果則證明,視覺剝奪還影響外側膝狀體內源性VIP的產生,使神經元的細胞器編碼、加工蛋白質的能力減弱。已知VIP 參與細胞內一系列磷酸化偶聯反應,與受體結合后,通過環磷酸腺苷(cyclic adenosine mono phosphate,cAMP)依賴的蛋白激酶途徑,促進葡萄糖的磷酸化和糖原分解,增強神經元的營養代謝。VIP 還可以促進腺苷酸環化酶的活化從而增加cAMP的作用[20]。在大腦皮層中,VIP和去甲腎上腺素相互協同,刺激皮層中cAMP的合成;同時VIP 還加速皮層神經元放電[21]。cAMP 作為細胞內重要的第二信使,是參與細胞內生物學功能和物質代謝的重要物質,其含量降低會影響細胞內磷酸化等多種胞內活動;同時VIP 缺乏導致視交叉上核神經元活動節律紊亂[22]。由此推測,視覺剝奪通過影響外側膝狀體中神經元的發育,致使VIP表達下降,而VIP 減少又使神經元的信號通路發生障礙,細胞內的能量代謝、糖代謝等活動均受到影響;同時神經元間的電傳導受阻。所以VIP表達下降會導致外側膝狀體神經元功能發育不良,而代謝異常的神經元又影響外側膝狀體功能表達,從而促進弱視的形成。
綜上所述,弱視的神經系統病理改變也發生在外側膝狀體;VIP在視覺發育中扮演著不可或缺的角色;異常視覺體驗導致外側膝狀體中VIP表達下降是弱視的重要發病機制之一。此結果為下一步對VIP 外源性干預視覺剝奪性弱視幼貓,探索VIP 治療弱視的藥理效果提供實驗基礎。由于方法和實驗條件的限制,本實驗只研究VIP在外側膝狀體中表達的變化,而外側膝狀體中神經元的微觀改變有待進一步觀察。研究神經元相關細胞器的變化可以進一步明確VIP 對弱視形成的影響。
