弗吉尼亞櫟母樹(shù)林外生菌根的真菌多樣性*
2020-03-05 07:31:20楊預(yù)展孫海菁陳連慶袁志林
林業(yè)科學(xué) 2020年1期
靳 微 楊預(yù)展 孫海菁 陳連慶 袁志林
(1. 林木遺傳育種國(guó)家重點(diǎn)實(shí)驗(yàn)室 中國(guó)林業(yè)科學(xué)研究院 北京 100091; 2. 中國(guó)林業(yè)科學(xué)研究院亞熱帶林業(yè)研究所 杭州 311400)
弗吉尼亞櫟(Quercusvirginiana,以下簡(jiǎn)稱弗櫟)原產(chǎn)美國(guó),通常生長(zhǎng)在沿海地帶的沙質(zhì)土壤中,能耐較高的土壤鹽分,是大西洋和海灣沿岸荒島林地的優(yōu)勢(shì)樹(shù)種(陳益泰等, 2007; 2017)。自2001年在長(zhǎng)三角引種成功至今,該樹(shù)種表現(xiàn)出常綠、抗風(fēng)、耐鹽、耐寒等多種優(yōu)良特性(陳益泰等, 2007),其中在浙江上虞海涂弗櫟人工林面積達(dá)160 hm2。弗櫟在林業(yè)工程建設(shè)中具有顯著的經(jīng)濟(jì)和生態(tài)效益,目前已開(kāi)展了該地區(qū)引種表現(xiàn)(陳益泰等, 2013)、種子產(chǎn)量(陳益泰等, 2015)、耐鹽試驗(yàn)(王樹(shù)鳳等, 2010; 2011)、扦插繁殖(陳雨春等, 2007)、容器育苗和抗風(fēng)耐鹽(陳益泰等, 2007)等多項(xiàng)研究表明,與松科(Pinaceae)、樺木科(Betulaceae)和楊柳科(Salicaceae)等高等植物類似,殼斗科(Fagaceae)中的櫟屬(Quercus)樹(shù)種也形成發(fā)達(dá)的外生菌根系統(tǒng)(Warcup, 1980; Szuba, 2015; Walkeretal., 2010),從而提高樹(shù)木養(yǎng)分吸收效率和環(huán)境適應(yīng)性。外生菌根(ectomycorrhiza,ECM)是擔(dān)子菌(少數(shù)為子囊菌和低等菌)菌絲體侵染植物營(yíng)養(yǎng)根形成的一種共生體(Cairneyetal., 1999),其主要特征是菌絲在幼根表面形成菌套(mantle),并侵入到根皮層細(xì)胞間隙形成哈蒂氏網(wǎng)(Hartig net)(Smithetal., 1997)。相對(duì)于其他櫟類樹(shù)種而言,對(duì)弗櫟外生菌根真菌調(diào)查研究只有零星報(bào)道,如Howe(1964)、Kropp等(1999)和 Guevara等(2013)分別發(fā)現(xiàn)土生空?qǐng)F(tuán)菌(Cenococcumgraniforme)、酒色紫蠟?zāi)?Laccariavinaceobrunnea)和少數(shù)幾種塊菌(Tuberspp.)可以與弗櫟形成外生菌根。在菌根化育苗試驗(yàn)中表明彩色豆馬勃(Pisolithustinctorius)(Daviesetal., 1990; Marxetal., 1997)也可以作為弗櫟的外生菌根真菌。但弗櫟引種至我國(guó)東南沿海地區(qū)后在新的氣候條件和立地條件下,其形成菌根能力如何、菌根真菌群落結(jié)構(gòu)及不同采樣時(shí)期的變化特征等基礎(chǔ)資料還十分匱乏。本文以浙江上虞海發(fā)園林有限公司培育的弗櫟母樹(shù)林為研究對(duì)象,通過(guò)菌根形態(tài)學(xué)描述和真菌擴(kuò)增子(ITS基因)的群落高通量測(cè)序(amplicon-based Illumina community profiling)平臺(tái),探討其外生菌根類型、群落結(jié)構(gòu)組成及動(dòng)態(tài)變化、優(yōu)勢(shì)外生菌根真菌的遺傳多樣性,為深入認(rèn)識(shí)弗櫟外生菌根菌、挖掘優(yōu)良菌株從而提升弗櫟栽培質(zhì)量提供基礎(chǔ)。
1 材料與方法
1.1 取樣及樣品處理
試驗(yàn)點(diǎn)位于在浙江上虞海發(fā)園林引種弗櫟母樹(shù)林基地(30°10′N(xiāo),120°42′E),該地區(qū)屬于亞熱帶海洋性季風(fēng)氣候,于1996年圍墾。土壤類型: 粉泥質(zhì)濱海鹽土,平均含鹽量0.43%,pH8.95。筆者在2017年對(duì)該樣地菌根類型及子實(shí)體種類的初步調(diào)查,發(fā)現(xiàn)在4—8月之間有較明顯的變化。故選定 2017年12月26日、2018年4月17日、5月29日和8月6日等4個(gè)時(shí)期采集菌根樣品,研究外生菌根真菌群落結(jié)構(gòu)的變化特征。從中國(guó)氣象數(shù)據(jù)網(wǎng)(data.cma.cn)獲取2017年10月—2018年11月樣地月平均降水量和月平均氣溫?cái)?shù)據(jù)(圖1)。選定樹(shù)齡為14年生的母樹(shù)林(面積0.33 hm2),隨機(jī)選擇5 塊樣方(4 m×4 m),每塊樣方選2 棵樹(shù),在每個(gè)樣方的頂點(diǎn)做好固定標(biāo)記,樣方間距離10 m以上。在距樹(shù)干0.5~1 m處,去除凋落物層,小心挖取帶土的菌根樣品放入自封袋里,采樣完畢立即將樣品帶回實(shí)驗(yàn)室。挑取發(fā)達(dá)菌根樣品,在清水中清洗、震蕩。洗凈后將樣品分為2部分,其中2塊樣方的菌根樣品用于形態(tài)觀察和特征描述; 另外3塊樣方的樣品經(jīng)表面消毒(3%過(guò)氧化氫5 min,75%酒精40 s,無(wú)菌水至少?zèng)_洗3次)后液氮速凍,放置-70 ℃保存,用于高通量測(cè)序。
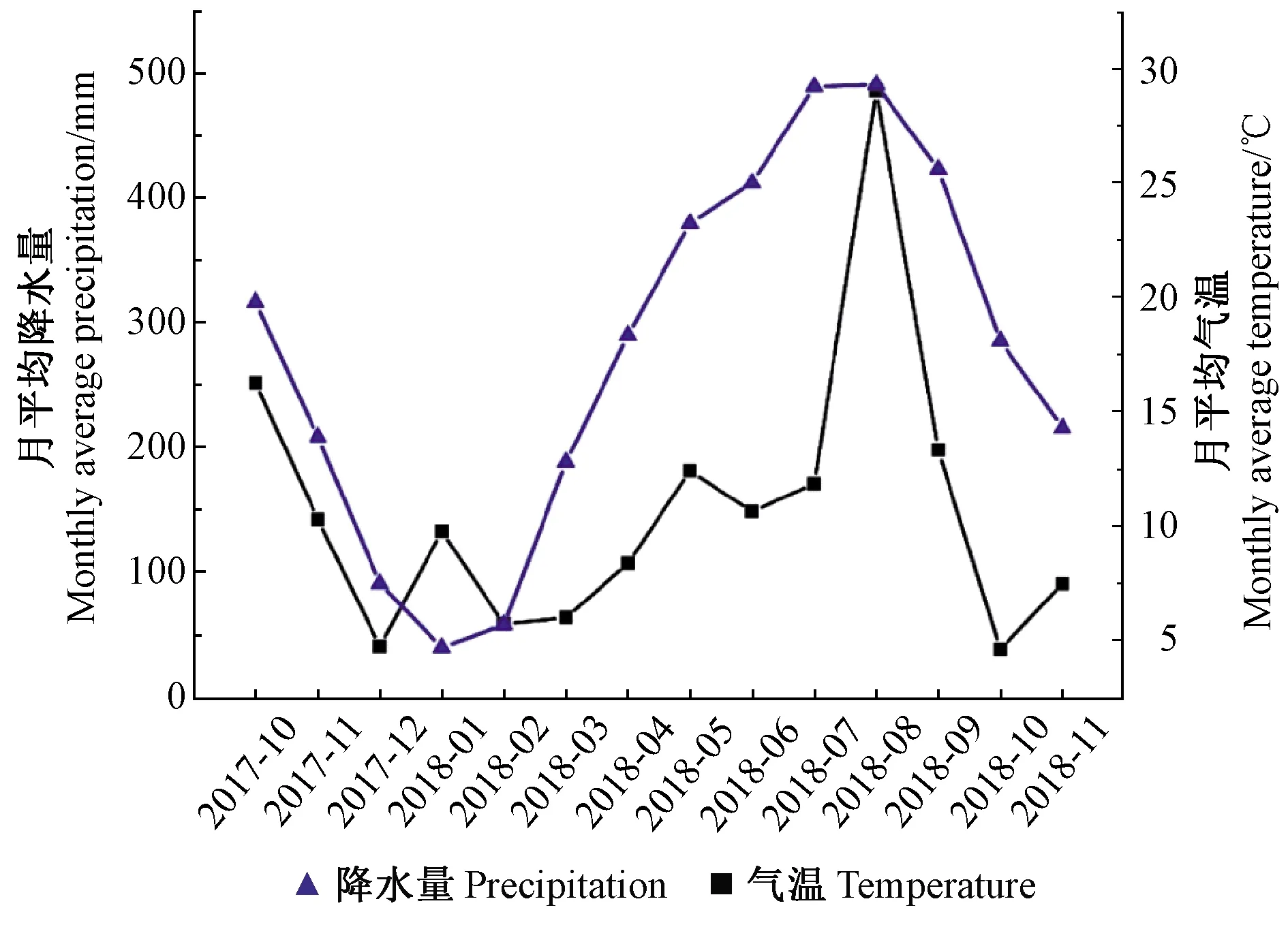
圖1 采樣地2017年10月—2018年11月的月平均降水量和月平均氣溫變化Fig.1 Monthly average precipitation and temperature from October 2017 to November 2018 in the sampling site
1.2 形態(tài)觀察
選取典型菌根在超景深顯微鏡(VHX-5000; Keyence,加拿大)下根據(jù)Agerer(2012)所提出的外生菌根形態(tài)分類要點(diǎn),觀察其分支類型、分支末端形狀、顏色、菌絲套表面質(zhì)地和外延菌絲等特征,并對(duì)其進(jìn)行拍照。用鑷子和長(zhǎng)繡花針,在體式顯微鏡下(Chinde XTL-81,中國(guó))對(duì)一個(gè)末端未分支的菌根進(jìn)行菌套剝離,在光學(xué)顯微鏡(Axio Scope A1,Zeiss,德國(guó))下觀察其菌套內(nèi)外層菌絲組織類型、菌絲排列方式、外延菌絲特征和菌索類型等解剖特征。
1.3 菌根根尖樣品擴(kuò)增子焦磷酸測(cè)序
每個(gè)采樣時(shí)期每塊樣方的菌根樣品按照不同顏色進(jìn)行分組,標(biāo)記好后用于高通量測(cè)序。大部分樣品包含3個(gè)生物學(xué)重復(fù)。個(gè)別樣品由于取樣時(shí)獲取量極少,且菌根不發(fā)達(dá),故將3個(gè)樣方的樣品混合成1份樣品,而菌根發(fā)達(dá)的樣品類型,則包含6個(gè)或12個(gè)重復(fù)。此外,在第1個(gè)采樣時(shí)期,白色菌根與少量黃色菌根交織在一起,難以分開(kāi),因此作為1種樣品類型來(lái)處理,總計(jì)共48份樣品,具體信息見(jiàn)表1。采用CTAB方法提取樣本的基因組DNA。用帶Barcode 的特異引物fITS7(5’-GTGARTCATCGAATCTTTG-3’)和ITS4(5’-TCCTCCGCTTATTGATATGC-3’) (Ihrmarketal., 2012)、Phusion 超保真 PCR 試劑盒(New England Biolabs,美國(guó))對(duì)真菌核糖體轉(zhuǎn)錄間隔區(qū)(ITS2)片段進(jìn)行擴(kuò)增。使用Qiagen Gel Extraction Kit(Qiagen, 德國(guó))試劑盒對(duì)目的條帶進(jìn)行回收純化,用TruSeq? DNA PCR-Free Sample Preparation Kit(Illumina, 美國(guó))建庫(kù)試劑盒進(jìn)行文庫(kù)構(gòu)建,構(gòu)建好的文庫(kù)經(jīng)過(guò)Qubit@ 2.0 Fluorometer(Thermo Scientific)和Agilent Bioanalyzer 2100檢驗(yàn)合格后,使用Illumina HiSeq2500平臺(tái)進(jìn)行PE250進(jìn)行雙尾測(cè)序(北京諾禾致源生物信息科技有限公司)。將測(cè)得序列共48個(gè)樣品的96份原始數(shù)據(jù)文件提交至European Nucleotide Archive數(shù)據(jù)庫(kù)(http:∥www.ebi.ac.uk/ena,登錄號(hào): RJEB28784)。
1.4 高通量測(cè)序數(shù)據(jù)處理與分析
測(cè)序原始數(shù)據(jù)去除引物接頭序列、去除各低質(zhì)量堿基后用FLASH(V1.2.7, http:∥ccb.jhu.edu/software/FLASH/)(Magoetal., 2011; Jiaoetal., 2016)進(jìn)行拼接,得到raw tags。參照QIIME(版本1.7.0, http:∥qiime.org/index.html, Caporasoetal., 2010)對(duì)數(shù)據(jù)進(jìn)行嚴(yán)格質(zhì)控得到high-quality clean tags(Bokulichetal., 2013),之后與物種注釋數(shù)據(jù)庫(kù)(UNITE,https:∥unite.ut.ee/)進(jìn)行比對(duì)檢測(cè)去除嵌合體序列(Haasetal., 2011),得到每個(gè)樣本的有效序列數(shù)據(jù)。利用Uparse 軟件(Uparse v7.0.1001, http:∥drive5.com/uparse/)(Edgar, 2013)以97%為閾值對(duì)有效序列劃分可操作分類單元(operational taxonomic unit,OTU)。選擇OTUs中出現(xiàn)頻數(shù)最高的序列作為OTUs的代表序列,并基于 QIIME平臺(tái)(版本1.9.1)的BLAST工具在UNITE數(shù)據(jù)庫(kù)中對(duì)OTU進(jìn)行物種注釋,并分別在目和屬的水平上統(tǒng)計(jì)各樣本的群落組成。以樣本中數(shù)據(jù)量最少的為標(biāo)準(zhǔn)對(duì)OTUs進(jìn)行均一化處理,后續(xù)的分析都基于均一化處理后的結(jié)果。不同采樣時(shí)間和不同顏色的外生菌根真菌群落組成的差異的非度量多維標(biāo)度(NMDS),采用R語(yǔ)言(版本 2.15.3)的vegan包實(shí)現(xiàn)。用QIIME(版本1.9.1)計(jì)算加權(quán)和非加權(quán)Unifrac距離,并用R語(yǔ)言進(jìn)行α-多樣性指數(shù)和β-多樣性指數(shù)組間差異分析。α-多樣性指數(shù)主要包括以下4個(gè)指標(biāo): 菌群豐度(Chao1和ACE)和菌群多樣性指數(shù)(Shannon和Simpson)。由于在OTU注釋結(jié)果中并未發(fā)現(xiàn)塊菌等子囊菌類群,因此在本研究中的所有結(jié)果均是基于擔(dān)子菌分析得到。
使用ClustX 1.81軟件進(jìn)行ITS2基因序列比對(duì),用GeneDoc軟件對(duì)序列進(jìn)行適當(dāng)編輯,去除頭尾不整齊的序列。最后在Phylip 軟件包(版本3.68)中使用Neighbor-Joining算法構(gòu)建系統(tǒng)發(fā)育樹(shù)。利用DnaSP(版本6.12.01,http:∥www.ub.edu/dnasp)計(jì)算幾個(gè)外生菌根真菌類群的種間核苷酸多樣性。

表1 弗櫟外生菌根采樣時(shí)間及高通量測(cè)序樣品信息Tab.1 Information of ectomycorrhizal root tips samples used for pyrosequencing

圖2 弗櫟白色菌根形態(tài)結(jié)構(gòu)與解剖結(jié)構(gòu)Fig.2 External and anatomic structures of white-colored ectomycorrhiza of Q. virginianaA-B: 外部形態(tài)特征Branching pattern of ectomycorrhiza; C: 外層菌套Outer layer of mantle; D: 內(nèi)層菌套Inner layer of mantle; E: 外延菌絲Emanating mycelium; F: 菌索Rhizomorph.
2 結(jié)果與分析
2.1 弗櫟不同顏色菌根形態(tài)和解剖特征
白色菌根大多不規(guī)則羽狀分支,0~3級(jí),少數(shù)單軸狀,疏水。末端變細(xì)。表面性狀為密羊毛狀,具大量棉絮狀外延菌絲,菌索透明。菌套內(nèi)外層均為密絲組織,菌套類型為A型。外延菌絲表面光滑,具鎖狀聯(lián)合。菌索未分化,松散纏繞(圖2)。
黑色菌根大多呈羽狀和塔狀,0~3級(jí),親水。少數(shù)呈未分支末端棒狀,頂端圓鈍。外層菌套為擬薄壁組織,表面光滑,菌絲細(xì)胞排列呈石堆狀,菌絲細(xì)胞三角至多角形,未見(jiàn)圓形細(xì)胞堆積,菌套類型為L(zhǎng) 型。內(nèi)側(cè)菌套為擬薄壁組織,菌絲細(xì)胞為擬薄壁細(xì)胞。外延菌絲極少且具有鎖狀聯(lián)合。菌索淺黃色未分化,表面光滑(圖3)。
黃色菌根大多不規(guī)則羽狀分支,偶見(jiàn)塔狀分支。0~2級(jí),親水。淺黃色,成熟菌根為黃色,未分支末端棒狀,頂端圓鈍。外層菌套由短刺狀菌絲構(gòu)成,菌套接近D型,內(nèi)層菌套為密絲組織。偶見(jiàn)外延菌絲,未見(jiàn)鎖狀聯(lián)合。菌索透明無(wú)色,松散排列未分化。多和弗櫟葉片凋落物粘連在一起,剝離時(shí)會(huì)將一部分菌套留在落葉組織中(圖4)。
在第3個(gè)采樣時(shí)期,除發(fā)現(xiàn)以上3種顏色的菌根外,還觀察到中間色(乳白色和深紅色)菌根(圖5)。從形態(tài)結(jié)構(gòu)看,深紅色菌根與黑色菌根較接近,乳白色菌根與黃色菌根較接近,但菌套外表平滑,無(wú)短刺狀菌絲。考慮到該2種顏色菌根可能代表不同的外生菌根真菌種類,因此在高通量測(cè)序分析時(shí),也將上述菌根樣品分成獨(dú)立的2組(表1)。3種主要菌根類型的形態(tài)學(xué)特征描述與鑒定結(jié)果見(jiàn)表2。

圖3 弗櫟黑色菌根形態(tài)結(jié)構(gòu)與解剖結(jié)構(gòu)Fig.3 External and anatomic structures of black-colored ectomycorrhiza of Q. virginianaA-C: 菌根不同的分支類型Different branching pattern of ectomycorrhiza; D: 外延菌絲Emanating mycelium; E: 外層菌套Outer layer of mantle; F: 內(nèi)層菌套Inner layer of mantle.
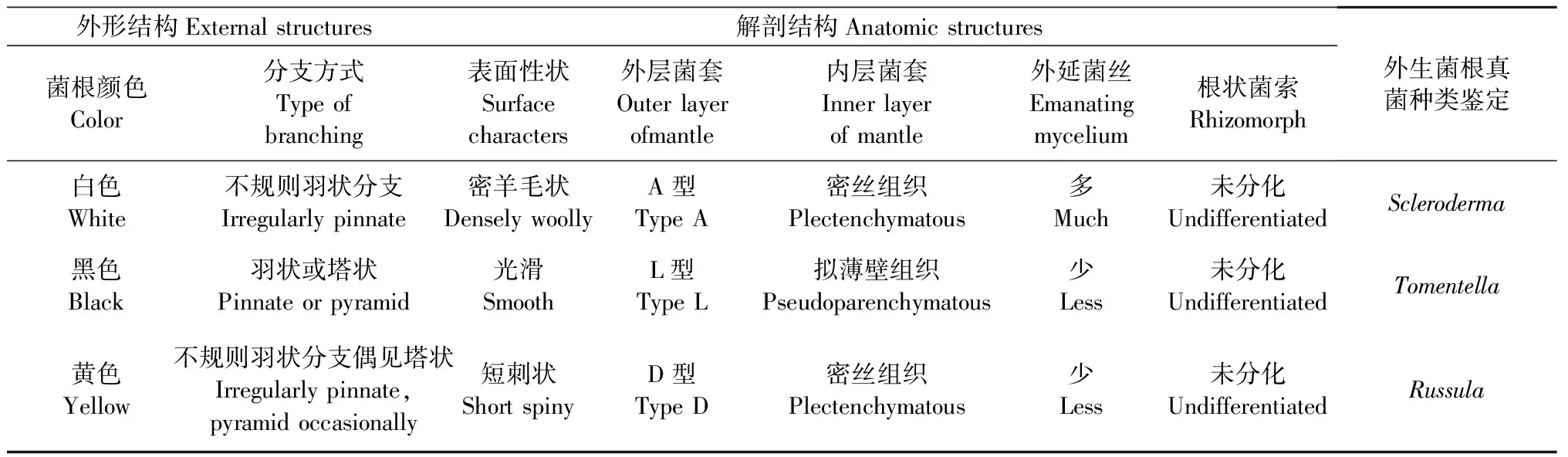
表2 弗櫟不同顏色外生菌根形態(tài)結(jié)構(gòu)、解剖結(jié)構(gòu)與鑒定Tab.2 Morphological descriptions of Q. virginiana-associated ectomycorrhizas and the corresponding fungal identities

圖4 弗櫟黃色菌根形態(tài)結(jié)構(gòu)與解剖結(jié)構(gòu)觀察Fig.4 External and anatomic structures of black-colored ectomycorrhiza of Q. virginianaA-B: 菌根形態(tài)及分支Branching pattern of ectomycorrhiza; C-D: 外層菌套的短刺狀結(jié)構(gòu)Short-spiny surface of mantle; E: 外層菌套Outer layer of mantle; F: 內(nèi)層菌套Inner layer of mantle.

圖5 弗櫟深紅色菌根(A、B)和乳白色菌根(C、D)形態(tài)結(jié)構(gòu)Fig.5 Morphological characteristics of the ectomycorrhizas with carmine and oyster white colored appearances in Q. virginiana
2.2 Illuminar HiSeq測(cè)序數(shù)據(jù)分析結(jié)果
基于擴(kuò)增子高通量測(cè)序平臺(tái),對(duì)弗櫟外生菌根真菌物種組成進(jìn)行分析鑒定。48份樣品的稀釋性曲線(rarefaction curve)均趨于平緩或達(dá)到平臺(tái)期,說(shuō)明測(cè)序數(shù)據(jù)量足夠,可真實(shí)反映弗櫟菌根真菌群落結(jié)構(gòu)(數(shù)據(jù)未列出)。從48份樣品共獲得5 720 430條序列,除去子囊菌和少數(shù)低等菌后,以97%序列相似性為閾值,共得到88個(gè)擔(dān)子菌OTUs。
2.3 弗櫟外生菌根真菌物種組成及種間遺傳多樣性
在UNITE真菌ITS數(shù)據(jù)庫(kù)進(jìn)行Blast比對(duì),得到OTU注釋結(jié)果。結(jié)果表明在弗櫟菌根中檢測(cè)到以下外生菌根真菌類群: 棉革菌(Tomentella)、硬皮馬勃(Scleroderma)、紅菇(Russula)、鵝膏菌(Amanita)、黏滑菇(Hebeloma)、豆馬勃(Pisolithus)、乳菇(Lactarius)和蠟殼耳菌(Sebacinales)(圖6)。根據(jù)各種顏色菌根樣品中OTUs的相對(duì)豐度并在屬的水平繪制柱狀統(tǒng)計(jì)圖,可以看出上述前3個(gè)屬分別對(duì)應(yīng)了黑色菌根、白色菌根和黃色菌根(圖7)。需要指出的是,除了白色菌根中硬皮馬勃占有極高的豐度外(均值達(dá)到92.25%),其余2種顏色菌根棉草菌和紅菇占有的比列相對(duì)較低,均值分別為52.69%和74.57%。在采樣過(guò)程中發(fā)現(xiàn)白色菌根易分離且菌套發(fā)達(dá)、菌絲生長(zhǎng)旺盛; 同時(shí)這3種顏色的菌根往往交織在一起。因此,在黑色和黃色菌根樣品中可檢測(cè)到一定比例的硬皮馬勃。此外,乳白色和深紅色菌根中,硬皮馬勃、棉草菌和紅菇均占有一定比例,但基于加權(quán)的聚類分析表明,乳白色菌根的真菌群落結(jié)構(gòu)組成與黃色菌根相似,深紅色菌根則與黑色菌根更為接近(圖8)。因此,推測(cè)這是同一菌根類型的不同發(fā)育階段。由圖6還可看出,某些豐度較低的外生菌根真菌類群表現(xiàn)出一定的季節(jié)或菌根類型的特異性,如OTU_14[沼生黏滑菇(Hebelomahelodes)]只在第4個(gè)采樣時(shí)期的黑色菌根中存在,OTU_133[彩色豆馬勃(Pisolithusarhizus)]主要在第1個(gè)采樣時(shí)期的黑色菌根中檢測(cè)到,OTU_7[蠟味紅菇(Russulacerolens)]只存在于第2個(gè)采樣時(shí)期的黃色菌根中。
為了評(píng)估硬皮馬勃、棉草菌和紅菇 3個(gè)重要類群的遺傳多樣性,提取了相應(yīng)OTU的序列,基于鄰接法(Neighbor-Joining,NJ)構(gòu)建了系統(tǒng)發(fā)育樹(shù)(圖9)。結(jié)果發(fā)現(xiàn),共有16個(gè)OTU屬于硬皮馬勒、23個(gè)OTU屬于棉草菌、14個(gè)OTU屬于紅菇屬。多數(shù)OTU形成了獨(dú)立的進(jìn)化分支并具有較長(zhǎng)的分支長(zhǎng)度,可能代表了一些新的分類單元。通過(guò)DnaSP軟件計(jì)算得到3個(gè)屬的OTUs核苷酸多樣性(π)值分別為0.061、0.129和0.117,表明棉草菌的種間遺傳多樣性最高。
2.4 3不同采樣時(shí)期弗櫟外生菌根真菌群落的變化特征
將每個(gè)采樣時(shí)期所有菌根樣品的OTUs數(shù)據(jù)進(jìn)行整合,分析不同采樣期弗櫟外生菌根真菌群落的變化特征。首先,在目的水平上比較了4個(gè)時(shí)期的群落組成變化(圖10),群落結(jié)構(gòu)呈現(xiàn)較明顯的演替,主要表現(xiàn)為: 硬皮馬勃(牛肝菌目,Boletales)在初始階段豐度較高,隨后呈逐漸下降趨勢(shì),但在所有時(shí)期豐度仍最高; 相反紅菇(紅菇目,Russulales)在初始階段比例極低,隨后呈較快增長(zhǎng),但豐度仍未超過(guò)硬皮馬勃; 棉草菌(革菌目,Thelephorales)在第2個(gè)時(shí)期的豐度躍升,其余時(shí)期較穩(wěn)定。實(shí)地采樣中也發(fā)現(xiàn),在冬季母樹(shù)林能觀察到硬皮馬勃子實(shí)體,而在5月份至少鑒定出2種紅菇子實(shí)體。該變化特征與硬皮馬勃和紅菇子實(shí)體發(fā)生時(shí)間基本吻合。
進(jìn)一步比較各個(gè)時(shí)期外生菌根真菌群落的β-多樣性指數(shù)(加權(quán)和非將加權(quán))表明,第1個(gè)時(shí)期與第4個(gè)時(shí)期的群落結(jié)構(gòu)組成差異最顯著(圖11)。通過(guò)計(jì)算4種α-多樣性指數(shù),表明第2個(gè)采樣時(shí)期菌根樣品的Chao1、ACE、Shannon和Simpson數(shù)值最高,且分別與第3和第4個(gè)時(shí)期的樣品之間的差異達(dá)到極顯著水平(Wilcoxon秩和檢驗(yàn),圖12)。基于OTUs的Venn圖顯示,雖然4個(gè)時(shí)期樣品共有25個(gè)相同的OTUs,但第2個(gè)采樣時(shí)期樣品種檢測(cè)到7個(gè)特有的OTUs(圖13)。非度量多維度(non-metric multidimensional scaling,NMDS)分析表明,除第3個(gè)時(shí)期和第4個(gè)時(shí)期的部分樣點(diǎn)聚在一起,其余時(shí)期的樣點(diǎn)基本能很好地分開(kāi),stress值為0.157(<0.2的閾值),說(shuō)明群落結(jié)構(gòu)組成仍存在較明顯差異(圖14)。
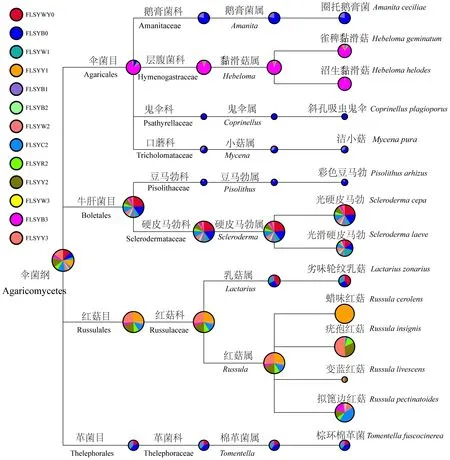
圖6 弗櫟外生菌根真菌物種組成及相對(duì)豐度Fig.6 Relative abundance of ectomycorrhizal fungal communities at different taxonomic levels in Q. virginiana圓圈的大小代表該分類單元的相對(duì)豐度。沒(méi)有鑒定到種水平的OTUs沒(méi)有納入分析。The relative abundance is expressed as a percentage. The size of pie chart indicates the relative abundance of corresponding taxonomic level in samples. OTUs not assigned to species level are not included for analysis.
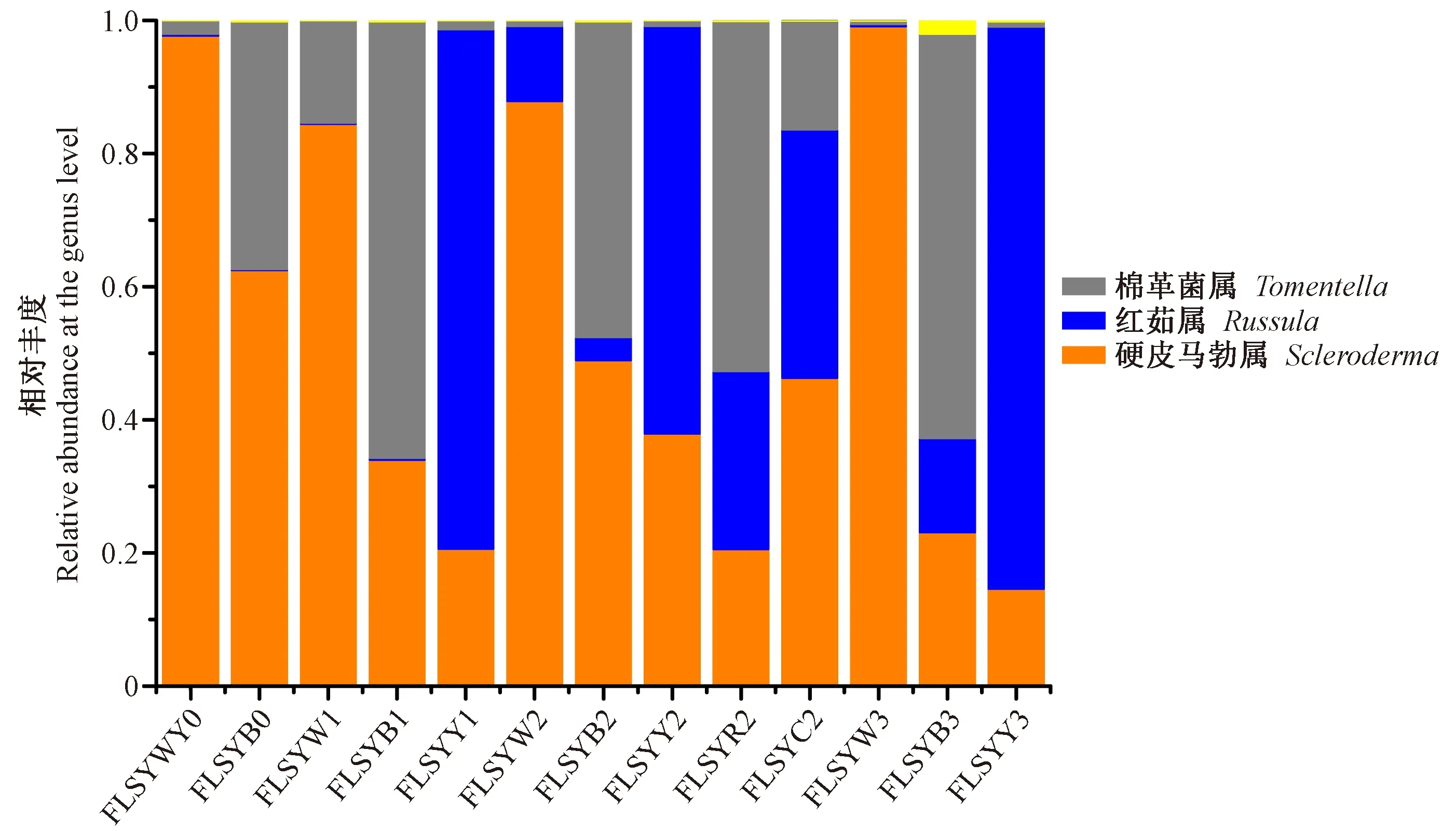
圖7 硬皮馬勃、棉草菌和紅菇等外生菌根真菌在4個(gè)采樣時(shí)期的相對(duì)豐度Fig.7 Relative abundances of the ectomycorrhizal Scleroderma, Tomentella and Russula in Q. virginiana root samples across four time points柱狀圖的樣品編號(hào)與表1對(duì)應(yīng),即4個(gè)采樣時(shí)期不同菌根類型樣品。下同。The sample names are the same as described in Tab.1. The same below.

圖8 基于加權(quán)的聚類分析比較4個(gè)采樣時(shí)期不同顏色菌根類型的外生菌根真菌群落結(jié)構(gòu)Fig.8 Comparison of difference of the ectomycorrhizal fungal communities from different colored mycorrhizal root samples of Q. virginiana across four time points based on weighted pair-groups method average (WPGMA) cluster analysis
3 討論
形成櫟樹(shù)外生菌根的真菌包括有擔(dān)子菌中的革菌科(Thelephoraceae)、紅菇科(Russulaceae)、蠟殼耳科(Sebacinaceae)、牛肝菌科(Boletaceae)、絲膜菌科(Cortinariaceae)(Cairney and Chambers, 1999)。此外,子囊菌中的塊菌(Tuber)、空?qǐng)F(tuán)菌(Cenococcum)和低等菌中的內(nèi)囊菌目(Endogonales)真菌也能與櫟樹(shù)形成菌根(Yamamotoetal., 2016)。早期對(duì)外生菌根真菌資源的研究主要通過(guò)描述外生菌根及其形成的子實(shí)體的多種形態(tài)特征進(jìn)行鑒定。然而,該方法不能真實(shí)、全面反映外生菌根真菌的群落結(jié)構(gòu)。近十年來(lái),基于真菌ITS基因的環(huán)境PCR方法(如克隆庫(kù)測(cè)序、Roche 454焦磷酸測(cè)序和Illuminar HiSeq等平臺(tái))助推了對(duì)菌根真菌結(jié)構(gòu)組成及多樣性的認(rèn)識(shí)。比如在厚葉櫟(Q.crassifolia)的根尖組織中就檢測(cè)到多達(dá)44個(gè)外生菌根真菌類群(Morrisetal., 2008)。
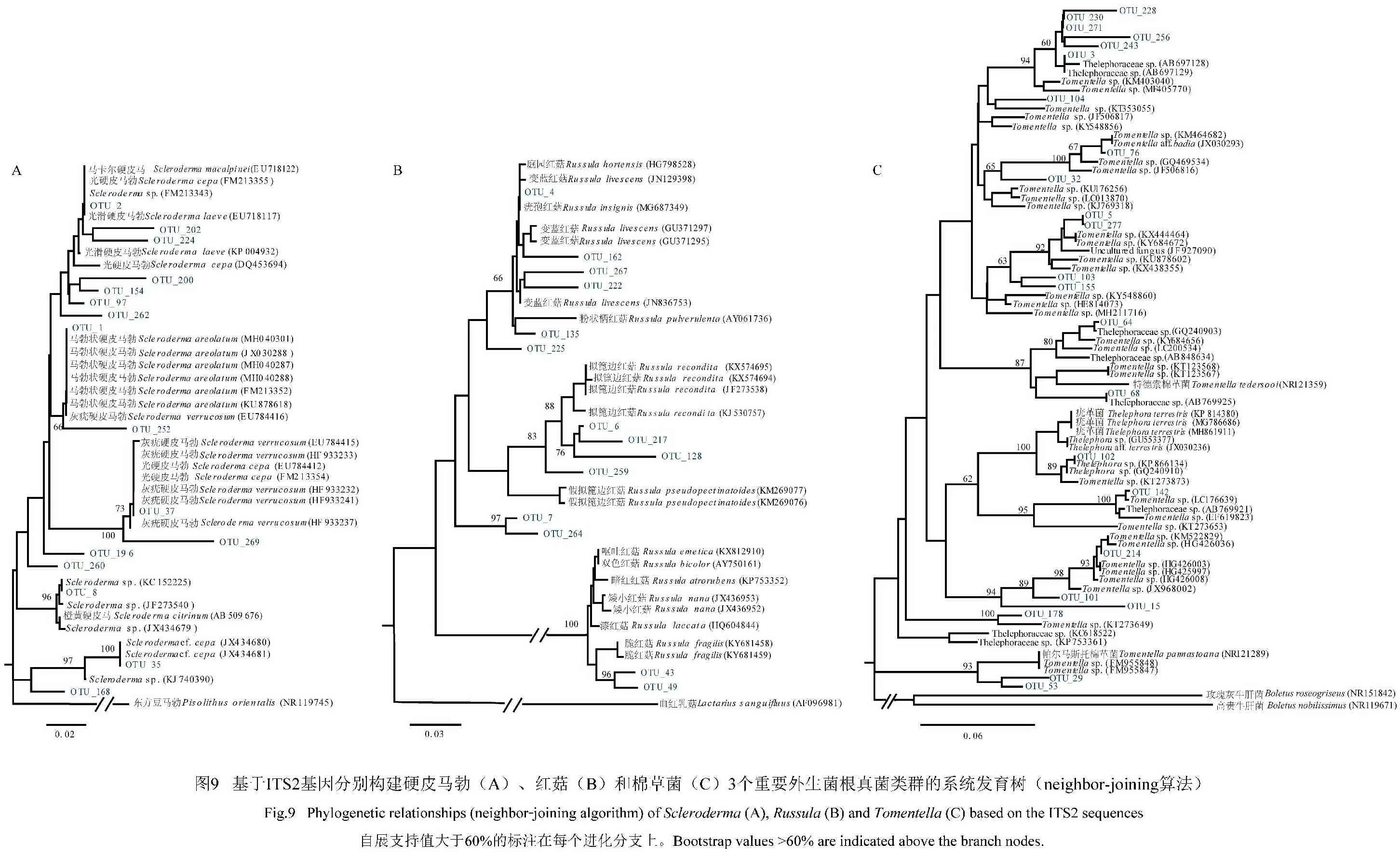
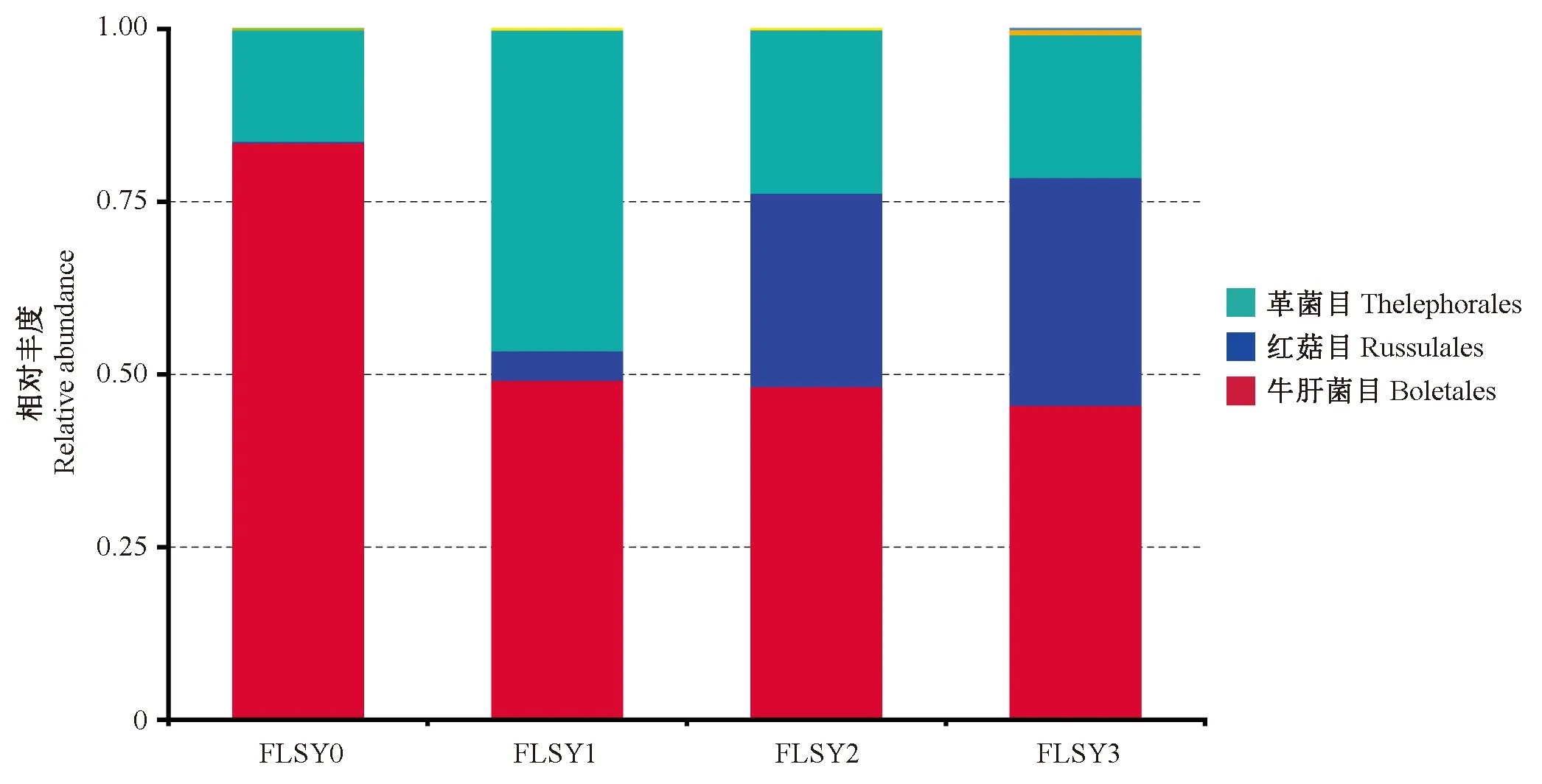
圖10 4個(gè)采樣時(shí)期弗櫟外生菌根真菌相對(duì)豐度(在目的水平上)Fig.10 Relative abundance of the detected fungal orders across four time points of pooled samples of Q. virginiana柱狀圖樣品編號(hào)代表每個(gè)采樣時(shí)期所有菌根類型的樣品(下同)。The sample names indicate the pooled samples across four time points (the same below).

圖11 基于UniFrac 距離的β-多樣性熱圖Fig.11 β-diversity heatmap based on the UniFrac distance數(shù)值(上下兩個(gè)值分別代表加權(quán)和非加權(quán))表示兩組樣品之間外生菌根真菌群落的相異系數(shù)。Here the weighted Unifrac (upper) and unweighted Unifrac distances are used as the measure index. The value indicates discrepancy coefficient between two group samples.

圖12 4個(gè)采樣時(shí)期弗櫟外生菌根真菌群落的α-多樣性指數(shù)Fig.12 α-diversity (based on ACE, Chao1, Shannon and Simpson) in ectomycorrhizal fungal communities across four time points of pooled samples of Q. virginiana利用Wilcoxon 秩和檢驗(yàn)進(jìn)行組間物種多樣性差異分析。Differences between the paired samples were performed by Wilcoxon test.

圖13 Venn圖分析不同采樣時(shí)期弗櫟外生菌根真菌OTUs組成及共有、特有OTUsFig.13 Venn diagram showing all OTUs detected in, shared by and exclusive to each sample type in Q. virginiana

圖14 基于NMDS分析的不同采樣時(shí)期弗櫟外生菌根真菌群落組成差異Fig.14 Difference of ectomycorrhizal fungal communities associated with Q. virginiana roots across four time points based on NMDS analysis4種顏色代表4個(gè)采樣時(shí)期,每個(gè)樣點(diǎn)代表1個(gè)重復(fù)樣品。Colors indicate the different sampling dates and data points represent individual samples
本研究利用擴(kuò)增子高通量測(cè)序技術(shù),對(duì)弗櫟外生菌根真菌的群落結(jié)構(gòu)組成及不同采樣期的變化特征進(jìn)行了較為細(xì)致的分析。從結(jié)果看,在不同采樣時(shí)期弗櫟能夠形成多種菌根類型,主要以硬皮馬勃、棉草菌和紅菇等外生菌根真菌為主,與已報(bào)道的其他櫟樹(shù)較為相似。上述3個(gè)類群還表現(xiàn)出豐富的種間遺傳多樣性。無(wú)論是種內(nèi)還是種間遺傳多樣性,均一定程度反映了其功能的多樣性,對(duì)于提高樹(shù)木的環(huán)境適應(yīng)性可能有重要的生態(tài)學(xué)意義(Hazardetal., 2018)。
土壤理化因子和氣候可能是影響外生菌根真菌群落差異的驅(qū)動(dòng)因素(陳云等, 2016; Castaoetal., 2017)。比如土壤水分含量對(duì)外生菌根真菌組成有顯著影響,高濕土壤能顯著增加塊菌的豐度,而革菌科在中等水分條件下維持較高活力(Cavender-Baresetal., 2009)。從降雨和氣溫變化的數(shù)據(jù)來(lái)看(圖1),從4月份開(kāi)始有明顯的增溫和降雨過(guò)程,而此時(shí)棉草菌的豐度最高,這一結(jié)果與上述研究結(jié)論是一致的。有研究表明,櫟類林中大部分外生菌根真菌類群的豐度在夏季最高(Voí?kováetal., 2014)。本研究結(jié)果顯示只有紅菇表現(xiàn)出該動(dòng)態(tài)變化規(guī)律,硬皮馬勃反而在冬季更占優(yōu)勢(shì),而棉草菌則在早春更為普遍。其他一些不常見(jiàn)的種類如Pisolithus和Hebeloma則只出現(xiàn)在某個(gè)采樣時(shí)期(圖7)。這些結(jié)果表明弗櫟不同的外生菌根真菌種類對(duì)氣候和土壤水分的響應(yīng)有較大差異,種類和豐度也存在較明顯的變化特征。這可能與真菌本身的生物學(xué)性狀和生態(tài)習(xí)性有關(guān),如有性無(wú)性繁殖權(quán)衡策略的不同、在逆境條件下代謝活力的差異等(Richardetal., 2011)。在對(duì)北美紅櫟(Q.rubra)和橡樹(shù)櫟(Q.prinus)混交林、蒙古櫟(Q.mongolica)外生菌根真菌季節(jié)動(dòng)態(tài)變化的研究中也得出相似的結(jié)論(Walkeretal., 2008; Kimetal., 2016)。另外,本研究還發(fā)現(xiàn)雖然第2次和第3次采樣時(shí)間只間隔40多天,但真菌群落結(jié)構(gòu)組成、多樣性和主要類群的豐度均經(jīng)歷了一次劇烈的變化(圖10和圖12),預(yù)示著在這個(gè)時(shí)間段內(nèi)降雨或者氣溫也發(fā)生了顯著改變,樹(shù)木光合生理、根系活力也處于較活躍狀態(tài),直接或間接造成了真菌群落各成員之間的此消彼長(zhǎng)。圖1數(shù)據(jù)看出,4月中旬至5月底,溫度逐漸升高,5月份的降水量在2018年上半年處于最高值。因此可理解該2個(gè)采樣時(shí)期弗櫟外生菌根真菌群落的明顯差異。本研究結(jié)果對(duì)深入認(rèn)識(shí)弗櫟外生菌根真菌多樣性、挖掘優(yōu)良菌株,從而提升弗櫟的栽培質(zhì)量具有科學(xué)意義和應(yīng)用價(jià)值。
4 結(jié)論
弗櫟引種到我國(guó)華東灘涂鹽堿地后,同樣能形成發(fā)達(dá)的外生菌根。棉革菌、硬皮馬勃和紅菇是3種主要類群且種間遺傳多樣性豐富,其中硬皮馬勃在各個(gè)時(shí)期均占優(yōu)勢(shì)。弗櫟不同的外生菌根真菌種類對(duì)氣候和土壤水分的響應(yīng)有較大差異,種類和豐度存在較明顯的變化特征。
