水分脅迫對不同抗旱性燕麥品種光合及產量形成的影響
2020-03-05 03:55:44趙寶平徐忠山武俊英劉瑞芳劉景輝
麥類作物學報 2020年11期
關鍵詞:產量
趙寶平,任 鵬,徐忠山,武俊英,劉瑞芳,劉景輝
(1.內蒙古農業大學,內蒙古雜糧工程技術研究中心,內蒙古呼和浩特 010019;2.內蒙古呼和浩特園藝科技試驗中心,內蒙古呼和浩特 010070)
燕麥(AvenasativaL.)屬于糧飼兼用作物,同時具有降脂控糖、降低心血管疾病等功能,因而燕麥原料市場需求量較大,而且不斷增長[1-3]。我國燕麥主要分布在西北等干旱半干旱地區。這些地區年降水量約在200~400 mm,農業種植以旱作為主,生育期間的干旱是限制燕麥等作物生長及高產的主要因素[4]。不同抗旱性燕麥品種的產量潛力存在較大差異[5]。因此,在水分脅迫下探究不同抗旱性品種的生理差異,對燕麥抗旱育種和栽培具有重要意義。
作物的光合作用是產量形成的物質基礎,較高的光合同化能力有助于作物高產[6]。水分虧缺會導致作物葉片光合能力下降,影響產量[7]。拔節期到開花后10 d被認為是影響燕麥產量的臨界期[8]。水分脅迫下燕麥葉片氣孔關閉,導致凈光合速率和蒸騰速率下降,提高瞬時水分利用效率[9]。水分脅迫下不同裸燕麥品種的氣孔導度與光合速率均顯著下降,但高產品種的光合能力降低與氣孔關閉無關[10]。隨著水分脅迫程度的增大,燕麥葉片氣孔逐步關閉,葉綠體等超微結構受損傷程度加劇[11]。水分脅迫還會引發植物光系統Ⅱ(PSⅡ)的電子傳遞速率降低以及PSⅡ結構或光捕獲體損傷[12]。但Ghotbi-Ravandi等研究認為,輕度水分脅迫下氣孔導度是限制強抗旱性大麥品種光合的主要因素,而在重度水分脅迫下,除了氣孔限制,PSⅡ結構破壞、電子傳遞速率減低等均影響了光合作用[13]。此外,水分脅迫下不同小麥品種的光合氣體交換、葉綠素熒光參數[14]、葉綠素含量以及產量[15]均存在顯著差異,說明作物光合作用過程研究不僅要考慮葉片氣體交換和氣孔特征,還要考察PSⅡ的結構和功能響應特征,從而揭示水分脅迫下作物光合調控機制并以此改善作物光合產量潛力[14]。葉綠素熒光技術由于可探究葉綠素蛋白體色素、激發能傳遞以及PSⅡ光量子傳遞效率等有用信息,近年來被廣泛用于作物光合作用性能研究中[16]。在燕麥上,Marcińska等利用葉片氣體交換和葉綠素熒光動力學參數等指標分析了不同燕麥品系的抗旱性差異[17],而將這兩類指標結合分析不同水分脅迫程度下不同抗旱性燕麥品種光合和產量形成的生理差異的研究尚未見報道。為此,本試驗研究不同水分脅迫下兩個抗旱性不同的燕麥品種葉片葉綠素含量、光合特性、葉綠素熒光特性、產量及其構成因素,以期為燕麥節水高產栽培與抗旱育種提供理論依據與技術支持。
1 材料與方法
1.1 試驗設計
1.1.1 供試材料
供試品種為河北省張家口農業科學院選育的水分敏感裸燕麥品種壩莜3號和內蒙古自治區農牧業科學院選育的抗旱皮燕麥品種蒙燕1號。
1.1.2 試驗設計
試驗在呼和浩特市科技園藝示范中心防雨棚內采用盆栽方式進行。設置2個品種和4個水分脅迫梯度,共8個處理,每個處理重復6次(3盆取樣測定光合色素指標,3盆用來測產考種),在拔節和抽穗兩個時期脅迫,共96盆。完全隨機排布,并每周調換一次盆的位置。4個水分梯度分別為重度脅迫[30%田間持水量(FWC)]、中度脅迫(45%FWC)、適度控水(60%FWC)和正常供水(CK,75%FWC);待各處理達到所設置的水分梯度后,分別在拔節期(出苗后41 d)、抽穗期(出苗后62 d)開始,每天采用稱重差值法實施7 d控制水分處理。處理結束之后恢復正常供水(≥75%FWC)。
盆栽土壤為沙壤土,田間持水量為16%。燕麥種植于高28 cm、直徑22 cm的塑料盆,每盆裝取自大田耕層土壤10 kg(干土重),并底施磷酸二銨和尿素各3 g,生育期不追肥。播種前每盆灌底墑水1.5 L,每盆播種50粒,三葉期每盆定苗25株。水分處理開始前和結束后維持正常供水直至成熟。
1.2 測定指標與方法
1.2.1 光合色素含量測定
用95%乙醇提取葉片光合色素,用分光光度計測定提取液在波長665、649和470 nm下吸光值,計算出葉綠素a、葉綠素b的含量及葉綠素a與葉綠素b的比值[11]。
1.2.2 葉片光合指標測定
采用Li-6400光合儀分別在燕麥拔節期、抽穗期脅迫結束后第一天測定最上部完全展開葉片的凈光合速率(Pn)、氣孔導度(Gs)、胞間CO2濃度(Ci)、蒸騰速率(Tr),并計算水分利用效率(WUE=Pn/Tr)。測定時間為上午9:00-11:00,每處理測定5株,取平均值為該處理測定值。
1.2.3 葉綠素熒光動力學參數測定
用英國漢莎公司FMS-2葉綠素熒光儀在兩個生育時期脅迫處理完成后當天進行葉綠素熒光動力學參數測定。選取植株上部受光一致的功能葉3片,葉片暗適應25 min后,測定初始熒光(Fo);隨后強閃光(6 000 μmol·m-2·s-1),脈沖時間(0.8 s)測定最大熒光(Fm);測定充分暗適應的PSⅡ最大光化學效率(Fv/Fm)。接著照射飽和脈沖光(6 000 μmol·m-2·s-1),測定光下最大熒光(Fm′)、Ft和PSⅡ實際光化學效率(ΦPSⅡ)。根據各處理數據,計算光化學猝滅系數(qP)和非光化學猝滅系數(NPQ)。
qP=(Fm′-Ft)/(Fm′-Fo)
NPQ=(Fm-Fm′)/(Fm-Fo)
1.2.4 產量及產量構成因素測定
在燕麥成熟期,收獲各處理盆中全部25株燕麥植株及籽粒,測定穗長、每穗小穗數、穗粒數(實粒數、每穗秕粒數)等,脫粒后測定穗粒重、千粒重、每盆產量等指標。
1.3 數據處理
試驗數據采用SPSS 18.0及Excel 2010進行統計分析。
2 結果與分析
2.1 水分脅迫對燕麥葉片葉綠素含量的影響
在拔節期與抽穗期,正常供水(CK,75%FWC)和適度控水(60% FWC)處理下兩個燕麥品種葉片葉綠素a(Chla)與葉綠素b(Chlb)含量均高于重度脅迫(30% FWC)處理(抽穗期蒙燕1號葉片Chla除外)(表1)。燕麥葉片葉綠素含量(Chla+b)隨水分脅迫強度的增加而逐漸降低,且重度水分脅迫處理與CK差異顯著。在拔節期,與CK相比,壩莜3號的葉片Chla+b值在重度脅迫、中度脅迫(45%FWC)、適度控水處理下分別降低了48.0%、27.6%和6.7%,蒙燕1號分別降低32.2%、21.0%和13.4%。此外,拔節期兩個品種Chla/b值隨著水分脅迫程度的增加均呈先增后降趨勢,兩個品種的Chla/b值在適度控水處理下均較高;抽穗期兩個品種Chla/b值隨水分脅迫強度的增加而增加,壩莜3號在不同水分處理間差異不顯著,蒙燕1號的重度脅迫、中度脅迫和適度控水處理較CK分別增加了28.4%、64.8%、237.5%,差異均顯著,且在重度脅迫下蒙燕1號的Chl a/b顯著高于壩莜3號。
2.2 水分脅迫對燕麥葉片光合特性影響
兩個燕麥品種的凈光合速率(Pn)對水分脅迫的響應表現相似,拔節期和抽穗期均隨水分脅迫程度的加劇而呈下降趨勢。相同水分脅迫下,兩個品種Pn除拔節期適度控水處理下蒙燕1號顯著高于壩莜3號外,其他處理下二者差異均不顯著。拔節期重度水分脅迫下壩莜3號和蒙燕1號的Pn分別較CK下降42.7%和34.2%;抽穗期分別下降27.8%和29.6%,拔節期水分脅迫對壩莜3號Pn的影響大于抽穗期(圖1A)。蒸騰速率(Tr)變化趨勢與Pn相似,隨著水分脅迫程度的增加呈下降趨勢。重度和中度脅迫下兩個品種的Tr與CK差異均顯著。拔節期水分脅迫對壩莜3號的Tr影響較大,而抽穗期水分脅迫對蒙燕1號影響較大(圖1B)。
拔節期兩個燕麥品種的氣孔導度(Gs)均隨著脅迫程度的加劇呈遞減趨勢。重度脅迫下,壩莜3號與蒙燕1號的Gs分別比CK降低55.1%和49.7%,差異均顯著(P<0.05)。抽穗期水分脅迫對Gs影響不顯著。兩個品種Gs隨著脅迫程度的增加均表現為先增后降,壩莜3號在適度控水條件下最高,蒙燕1號在中度脅迫下最高(圖1C)。抽穗期水分脅迫對兩個品種Ci影響均不顯著,拔節期水分脅迫下蒙燕1號Ci顯著低于CK,而壩莜3號的Ci值隨著脅迫程度加劇呈不斷增加趨勢(圖1D)。
隨著水分脅迫程度的增加,拔節期與抽穗期蒙燕1號水分利用效率(WUE)均呈現先增后降趨勢,適度控水處理下達到峰值,且高于壩莜3號。適度控水處理下蒙燕1號的WUE在拔節期與抽穗期分別較重度脅迫處理提高39.6%和 15.7%。在中度水分脅迫與適度控水處理下拔節期壩莜1號的WUE高于CK,重度水分脅迫下則表現相反。抽穗期壩莜3號的WUE在各水分脅迫處理下均低于CK,其中重度水分脅迫處理與CK差異達到顯著水平(圖1E)。
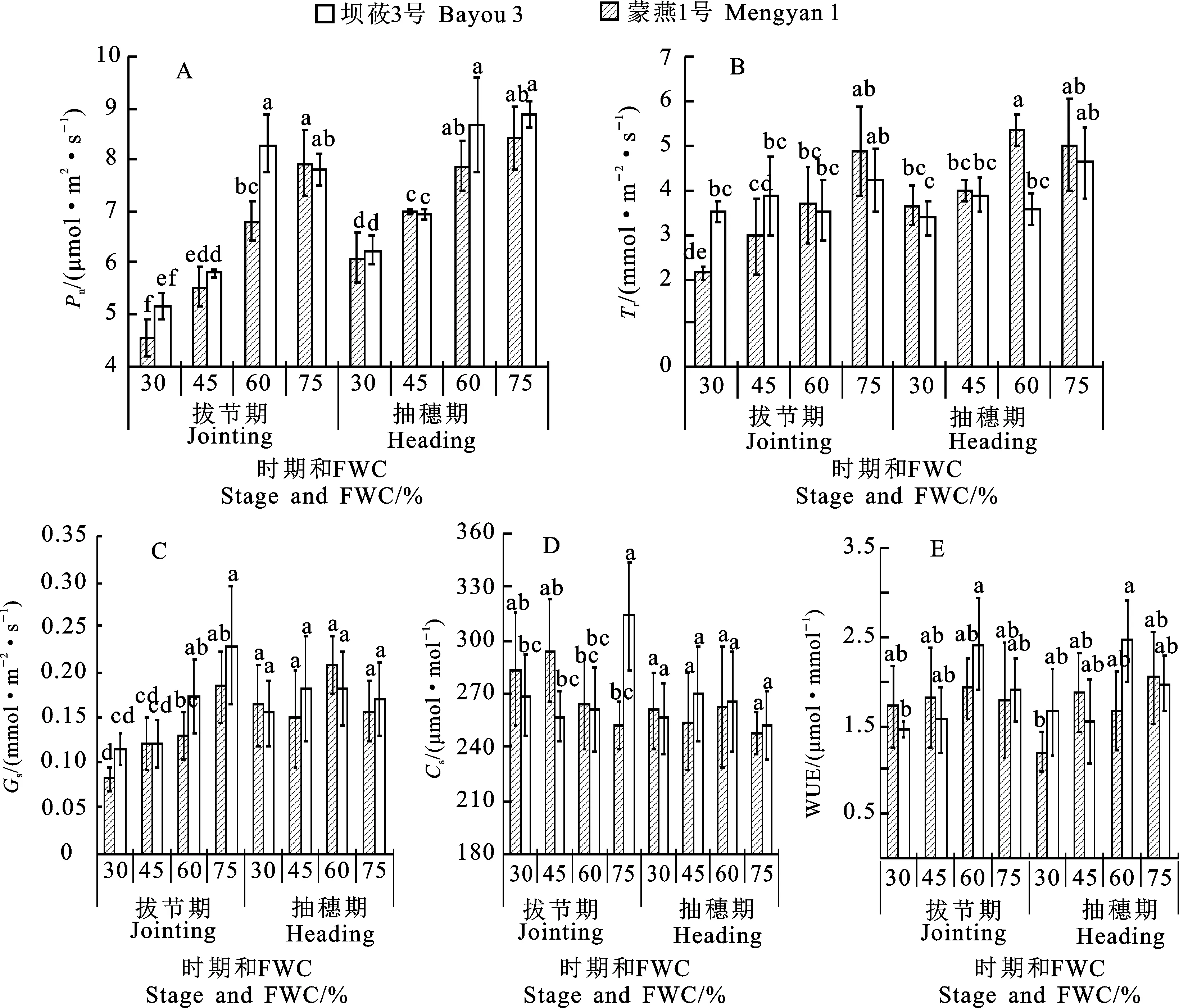
同一時期圖柱上不同字母表示不同處理間在0.05水平上差異顯著。圖2同。
2.3 水分脅迫對燕麥葉片葉綠素熒光參數的 影響
2.3.1 初始熒光(Fo)、最大熒光(Fm)及最大光化學效率(Fv/Fm)
不同生育時期,兩個燕麥品種葉片的Fo值均隨著水分脅迫程度的增加呈上升趨勢。重度脅迫下,壩莜3號和蒙燕1號的Fo值在拔節期較CK分別上升34.8%和8.2%,均顯著高于其他處理;在抽穗期分別上升39.0%和20.3%(圖2A)。Fm的變化與Fo的表現相反(圖2B),兩個燕麥品種的Fm均隨著水分脅迫程度的加劇而下降。在CK和適度控水下,壩莜3號的Fm高于蒙燕1號;在重度和中度水分脅迫下,壩莜3號的Fm急劇下降,且在重度水分脅迫下顯著低于蒙燕1號。
兩個生育時期兩個品種的Fv/Fm值均隨水分脅迫程度增加均呈下降趨勢(圖2C)。與CK相比,適度控水處理的降幅較小,中度和重度脅迫處理的降幅較大,且各水分脅迫處理下抽穗期的Fv/Fm值均低于拔節期;在重度和中度水分脅迫處理下兩個品種Fv/Fm值的降幅不同,蒙燕1號在拔節期和抽穗期重度水分脅迫下Fv/Fm值分別較CK下降4.8%和6.0%,而壩莜3號分別下降11.9%和14.5%;蒙燕1號的Fv/Fm值在兩個時期均顯著高于壩莜3號。
2.3.2 實際光化學效率(ΦPSⅡ)和電子傳遞速率(ETR)
不同生育時期燕麥葉片實ΦPSⅡ和ETR值均隨水分脅迫程度的增加而下降。拔節期和抽穗期水分脅迫下,兩個燕麥品種葉片的ΦPSⅡ均顯著低于CK(圖2D);重度水分脅迫下壩莜3號和蒙燕1號在拔節期、抽穗期分別較CK降低63.2%、65.0%和34.8%、29.2%,且相同水分條件下蒙燕1號的ΦPSⅡ均顯著高于壩莜3號。兩個品種的ETR隨著水分脅迫程度的加劇均呈下降趨勢,且壩莜3號的降幅大于蒙燕1號(圖2E)。在拔節期和抽穗期重度水分脅迫下,蒙燕1號的ETR分別較壩莜3號高108.0%和122.8%。
2.3.3 光化學淬滅系數(qP)和非光化學淬滅系數(NPQ)
兩個生育時期的水分脅迫均導致燕麥葉片qP下降,且降幅隨著水分脅迫程度的增加而增大(圖2F)。在抽穗期隨著水分脅迫程度的加劇,兩個品種的qP差異逐漸增大,在重度水分脅迫下蒙燕1號較壩莜3號高25.0%。而NPQ表現出與qP相反的趨勢,其值隨著水分脅迫加劇而逐漸增加(圖2G),且在各水分脅迫處理下壩莜3號均高于蒙燕1號。
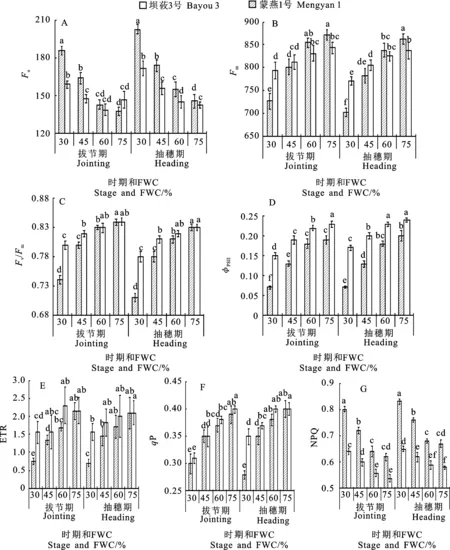
圖2 不同水分脅迫下燕麥葉片葉綠素熒光參數
2.4 水分脅迫對燕麥產量及其構成因素的影響
兩個生育時期的水分脅迫均對燕麥籽粒產量及其構成因素產生顯著影響(表2)。隨水分脅迫程度的增加,壩莜3號和蒙燕1號的穗長、單株小穗數均顯著下降。拔節期重度水分脅迫下壩莜3號、蒙燕1號穗長分別較CK(75%FWC)減少26.3%、21.8%;抽穗期重度和中度水分脅迫下,壩莜3號穗長分別較CK下降25.3%和21.4%,而蒙燕1號分別下降20.3%和8.0%。水分脅迫對壩莜3號單穗小穗數的影響大于蒙燕1號。拔節期適度控水、中度和重度水分脅迫下壩莜3號單穗小穗數較CK分別下降26.4%、30.7%和 39.0%;而蒙燕1號分別下降10.0%、21.1%和 20.5%;抽穗期適度控水、中度和重度水分脅迫下壩莜3號較CK分別下降20.8%、32.9%和 41.7%;而蒙燕1號分別下降5.3%、17.2%和 29.0%。兩個時期的水分脅迫對各品種的穗粒數影響均顯著(表2)。拔節期各水分脅迫處理下蒙燕1號的穗粒數均顯著高于壩莜3號,而在抽穗期除了重度脅迫下蒙燕1號的穗粒數較壩莜3號高32.1%之外,在其他水分處理下兩個品種間差異均不顯著。同一水分處理下蒙燕1號單穗實粒數顯著高于壩莜3號;在抽穗期重度水分脅迫下壩莜3號和蒙燕1號的單穗實粒數分別較CK下降24.0%和26.0%,其他水分處理間差異不顯著。拔節期重度水分脅迫下壩莜3號和蒙燕1號單穗秕粒數分別較CK增加134.2%和61.3%,其他處理間差異不顯著;而在抽穗期壩莜3號單穗秕粒數在不同處理間差異不顯著,但蒙燕1號的單穗秕粒數在重度脅迫下較CK增加72.8%。
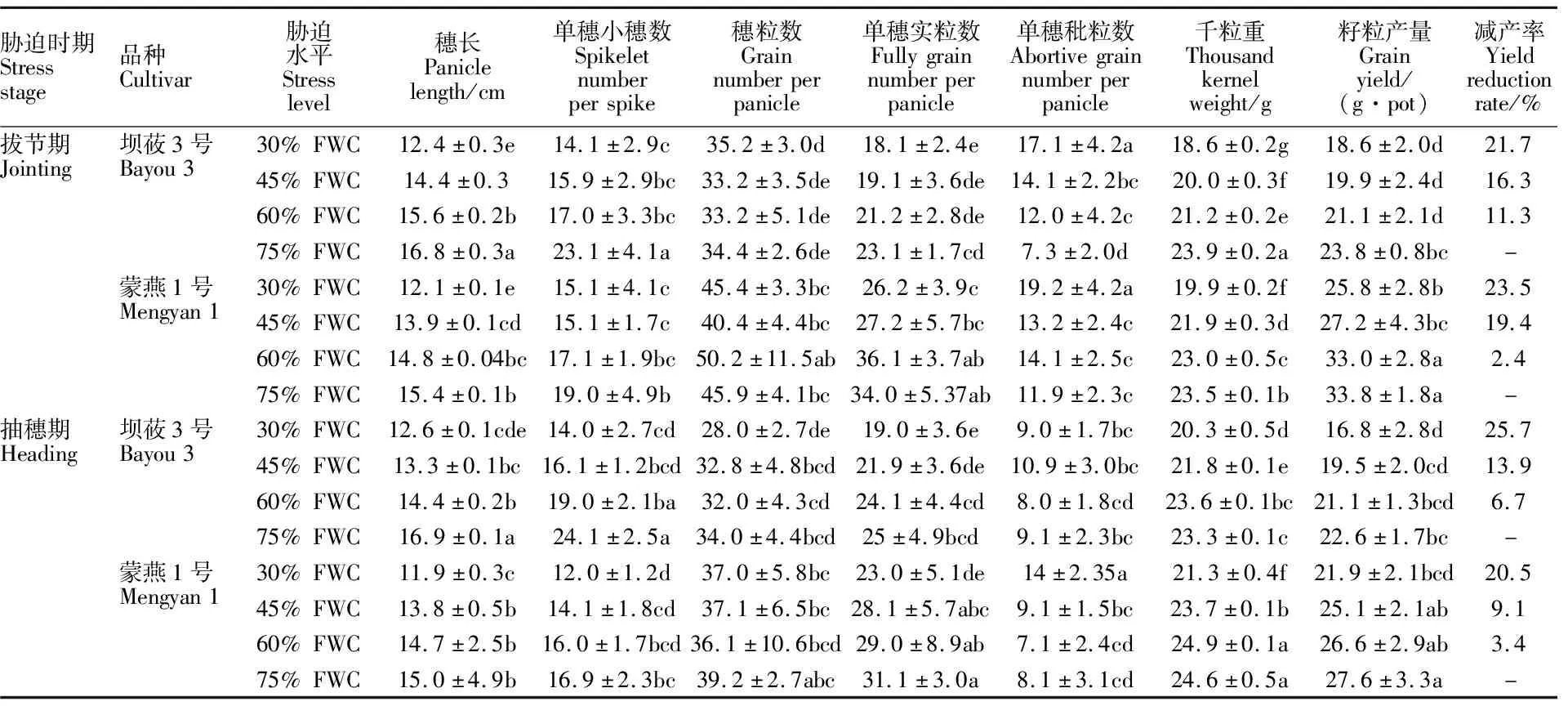
表2 水分脅迫下不同品種燕麥產量及其構成因素
兩個生育時期隨著水分脅迫加劇,千粒重均呈不斷下降趨勢。拔節期壩莜3號的千粒重在適度控水、中度脅迫和重度脅迫下分別下降 11.2%、16.5%和22.2%,而蒙燕1號分別下降 2.0%、6.9%和15.2%;在抽穗期,適度控水處理的千粒重與CK差異不顯著,重度水分脅迫下蒙燕1號和壩莜3號的千粒重分別較CK下降 13.6%和 12.9%。兩個生育時期隨水分脅迫程度的加劇,兩個品種的產量降幅逐漸增大,在拔節期適度控水處理下,壩莜3號和蒙燕1號分別減產 11.3%和2.4%;抽穗期壩莜3號在中度和重度水分脅迫下分別減產13.9%和25.7%;而蒙燕1號減產率分別為 9.1%和20.5%,且中度水分脅迫下產量與CK差異不顯著。
3 討 論
本研究表明,不同生育時期、不同水分脅迫程度下兩個燕麥品種光合特性、葉綠素熒光參數和產量形成的響應均存在差異。葉綠素含量的高低在一定程度上能反映葉片的光合能力[13]。在本試驗中,中度和重度水分脅迫下,兩個品種葉片Chla、Chlb、Chla+b值均顯著低于CK,且水分脅迫對水分敏感型品種壩莜3號影響大于抗旱品種蒙燕1號,表明干旱條件下抗旱燕麥品種的葉片光合色素具有較強保持能力。Chla/b值反映了植物對干旱脅迫的敏感性及抗旱性[18]。研究表明,干旱脅迫下金花茶幼苗葉片Chla/b值顯著降低[19]。本研究中,在拔節期水分脅迫下兩個品種的Chla/b值均降低,說明此期水分脅迫燕麥通過減少捕光蛋白色素復合物含量,保護PSⅡ以適應水分脅迫[20];在抽穗期重度水分脅迫下蒙燕1號Chla/b值顯著高于壩莜3號,說明抗旱性強的品種可能具有更小的捕光蛋白色素復合體,以減輕水分脅迫對PSⅡ的損傷[21]。
影響植物光合作用的因素可分為氣孔因素和非氣孔因素[22]。本試驗中,兩個時期不同水分脅迫處理下燕麥葉片Pn均低于CK;蒙燕1號拔節期各水分脅迫處理的Ci均低于CK,而抽穗期均高于CK,表明拔節期蒙燕1號Pn的下降是由氣孔因素引起的,而抽穗期可能是由非氣孔因素引起的;而壩莜3號兩個生育時期的Pn在水分脅迫下均下降,Ci均高于CK,說明該品種兩個時期水分脅迫Pn的下降可能受到非氣孔因素影響;同時,水分脅迫下PSⅡ實際光化學效率下降,說明其光合羧化效率降低[23]。本研究發現,適度控水下抗旱性強的蒙燕1號具有較高的葉片水分利用效率(WUE),這與Yang得出適度干旱或干濕交替灌溉有利于提高作物WUE的研究結果相似[24]。
本研究發現,不同水分脅迫下兩個品種葉片葉綠素熒光動力學參數表現不同。在中度與重度水分脅迫下,壩莜3號葉片Fo顯著高于蒙燕1號,而Fm和Fv/Fm均顯著低于蒙燕1號,Fo的增加說明水分敏感型燕麥品種在水分脅迫下 PSⅡ 反應中心受破壞程度和光化學反應受抑制程度更大[9],導致植株光合速率降低(圖1A);Fv/Fm的降低說明光捕獲體向PSⅡ的電子傳遞能力下降,可能與上述水分脅迫下Chla/b值降低、捕光蛋白色素復合物含量下降有關;而蒙燕1號在脅迫下Fv/Fm和Chla/b值更高,說明其抗旱性主要體現在光捕獲蛋白含量較高和PSⅡ活性較強[18,25]。同時,水分脅迫下ETR下降,導致激發能過量,使得光抑制作用加強[26]。此外,抽穗期各水分脅迫處理下Fv/Fm均低于拔節期,表明水分脅迫下抽穗期葉片PSⅡ活性中心受損程度大于拔節期。
水分脅迫下壩莜3號葉片實際光化學效率ΦPSⅡ顯著低于蒙燕1號,表明水分脅迫導致水分敏感性品種ΦPSⅡ下降幅度更大[27]。光化學淬滅系數qP反映了PSⅡ的電子傳遞活性,qP的減少意味著光抑制作用增加,因此逆境條件下作物維持較高的qP值以避免光抑制的作用至關重要[14]。本試驗試中兩個燕麥品種隨著水分脅迫程度的加劇,葉片qP逐漸降低,但蒙燕1號的qP在重度水分脅迫下顯著高于壩莜3號,說明抗旱品種能夠在水分脅迫下減輕光抑制作用。非光化學猝滅系數NQP反映了葉片吸收的光能以熱耗散的形式散失的程度。本研究發現,水分脅迫加劇使得NQP值逐漸增加,而且壩莜3號顯著高于蒙燕1號,可能是由于水分敏感型品種吸收的光能有很大比例沒有被光合利用,而以熱能形式(NPQ)散失[28]。
作物產量損失程度受水分脅迫時期、程度影響很大[29-30],并且不同基因型品種對水分脅迫的生理響應不同[31]。本試驗中,拔節期適度控水條件下壩莜3號減產達11.3%,在抽穗期中度和重度水分脅迫下壩莜3號減產13.9%~25.7%。說明壩莜3號在拔節期對水分更加敏感。拔節期正值燕麥幼穗分化期,水分脅迫影響小穗分化、小花發育,進而導致小穗數減少、秕粒數增加甚至籽粒敗育[32];水分脅迫下壩莜3號在該生育時期Pn及ΦPSⅡ、Fv/Fm均顯著下降,說明水分脅迫主要通過非氣孔因素限制葉片凈光合速率,抑制PSⅡ活性,光合產物減少導致小穗數、粒重下降,最終影響產量。而蒙燕1號在兩個生育時期適度控水處理下減產2.4%~6.7%,說明抗旱強蒙燕1號可適應適度干旱而不影響產量,在中度或重度水分脅迫下減產9.1%~23.5%,減產率低于壩莜3號(表2),在抽穗期中度脅迫下減產9.1%。這可能與其具有較高的Fv/Fm和Chla/b值,減輕了水分脅迫對光系統Ⅱ的損傷和光抑制作用,提高電子傳遞能力,進而提高其光合羧化效率[13],說明其在水分脅迫下光抑制作用較輕,可為穗部提供光合同化物,以保證小穗小花發育和結實,進而減少產量損失[33]。
4 結 論
兩個生育時期水分脅迫均顯著影響了燕麥葉綠素含量、光合速率以及熒光動力學參數,并且兩個品種間存在顯著差異。水分敏感品種壩莜3號葉片Pn及ΦPSⅡ、Fv/Fm受水分脅迫抑制作用顯著大于抗旱品種蒙燕1號,導致穗長、小穗數、千粒重顯著減少,顯著增加秕粒數,減產幅度較大,甚至在拔節期適度控水即顯著減產。抗旱品種蒙燕1號Fv/Fm和Chla/b值在水分脅迫下相對較高,減輕了水分脅迫對PSⅡ的損傷和光抑制作用,在適度控水下產量下降不顯著,中度和重度脅迫減產幅度小于壩莜3號。這說明抗旱性品種通過維持較高的光化學效率,以促進穗粒發育,進而減少產量損失。不同生育時期水分脅迫對兩個品種燕麥光合及產量影響效果不同,整體來看拔節期受水分脅迫影響程度更大。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
