漂洗和斬拌對海鱸魚肌球蛋白理化特性的影響
2020-03-03 11:55:00林婉玲劉芳芳李來好吳燕燕楊少玲楊賢慶
農業工程學報 2020年1期
關鍵詞:結構
林婉玲,劉芳芳,李來好,吳燕燕,楊少玲,黃 卉,楊賢慶,林 織
漂洗和斬拌對海鱸魚肌球蛋白理化特性的影響
林婉玲1,2,劉芳芳1,3,李來好1※,吳燕燕1,楊少玲1,黃 卉1,楊賢慶1,林 織4
(1. 中國水產科學研究院南海水產研究所,國家水產品加工技術研發中心,農業農村部水產品加工重點實驗室,廣州 510300;2. 韓山師范學院食品工程與生物科技學院,潮州 521041;3.上海海洋大學食品學院,上海 201306;4. 廣東順欣海洋漁業集團有限公司,陽江 529800)
為探究漂洗和斬拌對海鱸魚肌球蛋白理化特性的影響,分別提取原料、漂洗魚糜、斬拌魚糜的肌球蛋白,通過測定總巰基、活性巰基、表面疏水性、濁度等基本性質,并結合紅外光譜和原子力顯微鏡(atomic force microscope,AFM)技術對肌球蛋白二級結構和表面形貌進行研究。結果表明:相比原料,漂洗魚糜肌球蛋白總巰基含量降低19.5%,活性巰基增大63.9%,而斬拌魚糜肌球蛋白總巰基和活性巰基的質量分數相比漂洗魚糜分別降低22.6%和66.8%,漂洗魚糜的肌球蛋白變性程度最大;肌球蛋白濁度和表面疏水性在經過漂洗和斬拌均增大,漂洗對表面疏水性影響更大,斬拌對濁度影響更大;紅外光譜研究結果顯示,漂洗對二級結構影響更明顯,原料經過漂洗后,-螺旋相對含量降低了33.16%,無規則卷曲相對含量增加了79.42%,-折疊和-轉角分別增加1.11%和10.38%,斬拌后,魚糜肌球蛋白二級結構變化率較低;漂洗和斬拌都可改變肌球蛋白的表面形貌,使肌球蛋白聚集簇明顯減小,聚集高度增加。研究結果證明,漂洗和斬拌對肌球蛋白理化特性有很大的影響,是魚糜具有更好的凝膠性能的重要步驟。
蛋白;魚;檢測;肌球蛋白;變性聚集;二級結構;表面形貌
0 引 言
加熱條件下,肌原纖維蛋白在鹽的作用下水溶性增強,肌球蛋白結構展開,失去其非共價穩定結構,隨后分子間又相互締合,并結合大量的游離水分子,形成具有彈性的凝膠體[1]。肌球蛋白占肌原纖維蛋白總量的50%~55%[2],是肉中含量最高的一種蛋白質,也是肌肉中唯一可以單獨通過加熱即可形成凝膠的蛋白質[3],凝膠性、持水性等作為肉類產品重要的特性,與肌球蛋白理化性能有著密切的聯系[4],肌球蛋白在肌肉蛋白凝膠和肉的結合中發揮著主要的作用,并且肌球蛋白在體外不穩定,即使在低溫下也能在溶液中自發聚集[5-6],進而影響蛋白質凝膠性能。因此對肌球蛋白結構和變性聚集受加工條件的影響研究,有助于指導肉制品的加工。目前已有大量研究表明,肌球蛋白在加熱過程中巰基(總巰基、活性巰基)、濁度、表面疏水性以及結構都會發生變化[7-8]。活性巰基含量的變化在一定程度上可以反應肌球蛋白的變性程度[9],而濁度反應溶液中不溶懸浮顆粒的大小和數量,可以粗略地估計蛋白質的聚集程度。但是,目前關于漂洗和斬拌等工藝對肌球蛋白這些理化特性的影響的研究較少。漂洗和斬拌是魚糜加工過程中重要工藝,對魚糜品質的形成有重要的作用。已有研究表明,經過兩次漂洗,白鰱魚肌球蛋白交聯反應程度明顯增大[10]。另外也有研究發現,隨著斬拌時間延長,魚肉蛋白質粒徑先減小后增大,證明斬拌會使蛋白質變性聚集,形成微凝膠[11],同時斬拌可以使蛋白質的構象發生改變,并且魚糜凝膠性能與蛋白質構象緊密相關[12]。因此,為了研究魚糜加工過程中蛋白質凝膠特性的變化,進一步研究漂洗與斬拌對揭示肌球蛋白在魚糜加工過程中凝膠變化有重要意義。
原子力顯微鏡(atomic force microscope,AFM)與普通的顯微鏡相比具有明顯的優勢,比如原子級高分辨率,橫向分辨率達到1 nm,縱向分辨率達到0.1 nm,作為一種新型的檢測蛋白質聚集的方法,其可觀測物質表面的形貌特征,掃描圖也可反應蛋白質的聚集情況,因此可利用該技術檢測肌球蛋白聚集平面狀態下的三維圖像[13],更直觀的表示肌球蛋白在漂洗和斬拌過程中的變性聚集情況。因此,本文利用原子力顯微鏡對漂洗和斬拌過程造成的肌球蛋白聚集形態進行研究,并結合濁度、巰基、疏水性以及蛋白質二級結構變化等對肌球蛋白理化性質變化進行分析,以期為肌球蛋白的變性聚集的研究提供參考意見。
1 材料與方法
1.1 材料與試劑
海鱸魚(),購自于廣州市海珠區華潤萬家超市,平均質量700 g。
食鹽、白砂糖,食品級,廣州海珠區客村華潤萬家超市;乙二醇雙(2-氨基乙基醚)四乙酸(EGTA)、三磷酸腺苷二鈉(ATP-Na2)、5、5′-二硫代雙(2-硝基苯甲酸)(DTNB)、乙二胺四乙酸(EDTA)、8-苯胺-1-萘磺酸(ANS)以及其余常見試劑均為分析純,廣州領馭生物科技有限公司;BeyoColorTM彩色預染蛋白,購于上海碧云天生物技術有限公司;三羥甲基氨基甲烷(Tris),分析純,美國Sigma公司;-巰基乙醇、十二烷基硫酸鈉(SDS),電泳級,上海麥克林生化科技有限公司。
1.2 主要儀器與設備
絞肉機,廣東省佛山市順德區陳村鎮恒基隆五金機械廠;打漿機,湖南省長沙固利食品機械有限公司;IKA-T25組織勻漿機,德國IKA公司;HH-4快速恒溫數顯水浴箱,常州澳華儀器公司;Sunrise-basic吸光酶標儀,德國TECAN公司;Alpha1-4冷凍干燥機,德國Christ公司;IRAffinity-1紅外光譜儀,日本島津公司;3K30臺式高速冷凍離心機,德國Sigma公司。
1.3 試驗方法
1.3.1 魚糜制作
將鮮活海鱸魚在20 min內運回實驗室,冰暈后宰殺清洗并取肉,并進行漂洗(水洗1次、0.2%鹽洗1次,每次4 min,肉液比1:4),制成漂洗魚糜,然后進行擠壓脫水,保持含水率低于80%,再進行斬拌(空斬5 min后加入食鹽(1%)和白砂糖(1%)繼續斬拌10 min,斬拌溫度在4~10 ℃),制成斬拌魚糜,備用。
1.3.2 肌球蛋白提取
根據Park等[14]的方法,略微修改,為了防止蛋白質變性和蛋白質水解,所有用于制備肌球蛋白的溶液要在4 ℃下進行保存。稱取樣品,加入10倍體積的試劑A(0.1 mol/L KCl和20 mmol/L Tris-HCl緩沖液,pH值為7.5)。用分散勻漿器11 000 r/min條件下將其進行均質1 min。混合物在4 ℃下放置15 min,利用3K30高速冷凍離心機在3 000 g下離心5 min。向所得沉淀物中加入5倍體積的試劑B(含0.45 mol/L KCl、5 mmol/L-巰基乙醇、0.2 mol/L醋酸鎂(Mg(CH3COO)2)、1 mmol/L乙二醇-雙-(2-氨基乙基)四乙酸(EGTA)和20 mmol/L Tris-HCl緩沖液,pH值為6.8);再加入ATP-Na2使其成為最終濃度為5 mmol/L的懸浮液,使肌球蛋白與肌動蛋白解離,在4 ℃下放置60 min。上述懸浮液在12 000 g下離心10 min。取出上清液,加入5倍體積的1 mmol/L KHCO3進行緩慢稀釋,在4 ℃下保存15 min。再將此混合物在20 000 g下離心10 min所得沉淀物用2.5倍體積的試劑C(含0.5 mol/L KCl和5 mmol/L-巰基乙醇的20 mmol/L Tris-HCl緩沖液,pH值為7.5);進行懸浮。所得懸浮液在4 ℃下保存10 min,再緩慢加入2.5倍體積的1 mmol/L KHCO3。同時,加入MgCl2使其最終濃度為10 mmol/L。將形成的混合物在4 ℃下保存12 h,然后在20 000 g下離心15 min,取沉淀物,即為肌球蛋白。沉淀物用1倍體積溶液D(含0.6 mol/L NaCl的20 mmol/L Tris-HCl緩沖液,pH值為7.0)溶解,即為肌球蛋白溶液。
1.3.3 肌球蛋白純度鑒定
將提取的肌球蛋白溶液質量濃度調至2 mg/mL,并與2×上樣緩沖液1∶1混合,沸水浴5 min,然后12 000 r/min、4 ℃離心5 min得到上清液用于電泳。marker上樣量為5L,樣品上樣量為10L,分離膠為10%,濃縮膠為4%,以80 V恒壓是樣品在濃縮膠部分濃縮成一條線,然后將電壓切換至120 V,在恒壓下直至溴酚藍指示劑到達分離膠邊緣,電泳結束,染色液為0.1%的考馬斯亮蘭R-250-乙醇冰醋酸溶液;脫色液為水-乙醇冰醋酸溶液。最后用quantity one軟件進行蛋白純度分析。
1.3.4 濁度
參考Cao等[15],將肌球蛋白濃度調節至1 mg/mL,在340 nm測定吸光值,以吸光值來表達肌球蛋白濁度大小。
1.3.5 總巰基和自由巰基的測定
參考Wei等[16],略作修改。
總巰基:移取0.5 mL 4 mg/mL的肌球蛋白溶液,加入4.5 mL 0.2 mol/L Tris-HCl(含8 mol/L尿素,2%SDS,10 mmol/L EDTA,pH值為6.8)的緩沖液,震蕩混勻,并加入0.5 mL 0.1%DTNB(0.1 gDTNB溶于100 mL 0.2 mol/L Tris-HCl,pH值為8.0)緩沖液中,4 ℃避光反應1 h,于412 nm處測定吸光值。
自由巰基:移取0.5 ml 4 mg/mL的肌球蛋白溶液,加入4.5 mL 0.2 mol/L Tris-HCl不含尿素的緩沖液,震蕩混勻,其余操作同上。
巰基計算公式:
SH(mol/105g)=×÷(×)(1)
式中為412 nm處的吸光值,為稀釋倍數,本方法中為11,為摩爾消光系數,為13 600 L/(mol×cm),C為蛋白濃度,4 mg/mL。
1.3.6 表面疏水性的測定
參考Yongsawatdigul等[17],采用ANS法測定,用0.6 mol/L NaCl的20 mmol/L Tris-HCl,pH值為7.0的緩沖液將所提取的肌球蛋白溶液進行梯度稀釋,濃度為0.062 5、0.125、0.25、0.5、1 mg/mL,在激發波長380 nm,發射波長470 nm條件下利用酶標儀測定熒光強度,利用蛋白濃度-熒光強度作圖,表面疏水性S0-ANS以蛋白濃度-熒光強度曲線斜率表示。
1.3.7 肌球蛋白二級結構的測定(紅外光譜)
參考林婉玲等[18],首先將肌球蛋白溶液調節至相同的蛋白濃度(10 mg/mL),然后在?80 ℃冰箱中凍結4 h以上,然后置于真空冷凍干燥機中干燥直至肌球蛋白樣品完全干燥為止,即得到肌球蛋白凍干樣品。紅外光譜的測定是將肌球蛋白凍干樣品與提前干燥的無水溴化鉀以1:100充分研磨混勻,然后壓片。測定條件為分辨率4 cm-1,室溫下重復掃描24次,掃描范圍4 000~400 cm-1。掃描結果用Peak fit 4.12軟件進行去卷積,高斯擬合,分析蛋白質二級結構。
1.3.8 原子力顯微鏡(AFM)
樣品前處理:將肌球蛋白溶液稀釋至20g/mL,準確吸取10L蛋白溶液滴于直徑為1 cm的圓形云母片上,室溫條件下在超凈臺上自然干燥,形成蛋白質吸附層,然后用超純水沖洗吸附層以消除鹽分的干擾,繼續自然干燥,制成測試樣品。蛋白質顆粒的粒徑可以用干燥狀態下從云母片基底表面到蛋白質顆粒表面的垂直距離表示,原子力顯微鏡掃描圖可以反映蛋白質的聚集程度。
AFM成像:AFM探針為商用氮化硅針尖,操作模式為分子力模式,共振頻率為150~200 kHz,彈性系數為5 N/m。本試驗圖像掃描范圍均為3m×3m,每個樣品至少選取10個不同區域進行掃描,確保得到可重復的高質量圖像,所有圖像均在NanoScope Analysis離線軟件上進行,圖片先進行Flatten平整化處理,以消除慢掃描方向上的低頻噪音造成的傾斜、扭曲、跳線等的影響,對每條掃描線進行補償,使得到的圖像更加真實。
1.3.9 數據分析
所有數據均采用Origin 9.0和Excel 2010進行作圖,試驗數據用SPSS 22.0統計軟件進行分析,結果用平均值±標準差來表示,多組數據之間的差異用單因素ANOVA檢驗兩兩比較的Duncan’s比較模型,<0.05表示差異顯著。
2 結果與討論
2.1 肌球蛋白純度
提取的海鱸魚原料、漂洗魚糜、斬拌魚糜的肌球蛋白的SDS-PAGE圖如圖1所示,通過quantity one軟件進行蛋白純度分析得知,提取的肌球蛋白純度大于80%,可作為本試驗原料使用。
2.2 巰基和活性巰基
漂洗和斬拌對肌球蛋白的變性和聚集都有一定的影響,巰基作為肌球蛋白中具有最高反應活性的基團,與其結構的穩定性密切相關,并且巰基含量的變化可以反應蛋白質的氧化變性程度[19]。其中總巰基包括活性巰基和隱藏巰基[20],總巰基指暴露在蛋白表面和埋藏在蛋白分子內的巰基,而活性巰基指暴露在蛋白分子表面的巰基[21]。
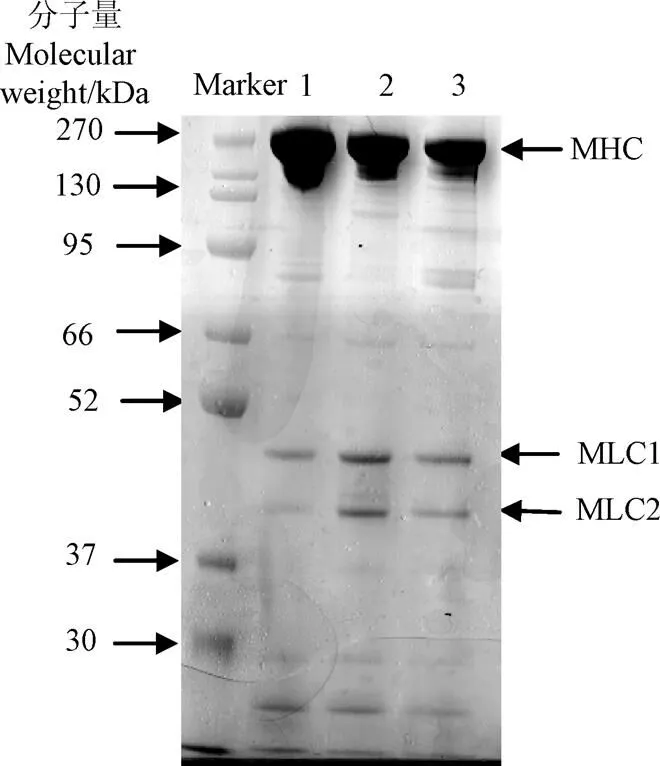
注:1、2、3分別代表海鱸原料、漂洗魚糜、斬拌魚糜肌球蛋白電泳條帶,MHC、MLC分別代表肌球蛋白重鏈和肌球蛋白輕鏈。
如圖2所示,漂洗和斬拌后魚糜肌球蛋白總巰基含量顯著下降19.5%(<0.05),從新鮮肉的8.51 mol/105g分別減少到6.85和5.3 mol/105g,而活性巰基在漂洗過程中顯著增加,從1.39增加到3.86 mol/105g,增大了63.9%,后經斬拌減小到1.28 mol/105g,活性巰基與總巰基在漂洗階段的比值為0.56,顯著大于原料和斬拌期間的比值。相比漂洗魚糜,斬拌魚糜肌球蛋白總巰基和活性巰基的質量分數分別降低22.6%和66.8%,在蛋白質中,總巰基含量下降表明活性巰基氧化生成二硫鍵,活性巰基的增加表明蛋白質結構的伸展[22],活性巰基與總巰基的比值可以反映蛋白質三級結構的解折疊程度,并且其比值與蛋白質解折疊度呈正相關關系[23]。這主要是由于漂洗過程中活性氧攻擊蛋白質,并且含有巰基的半胱氨酸屬于氧化敏感氨基酸,經過氧化易形成二硫化物。
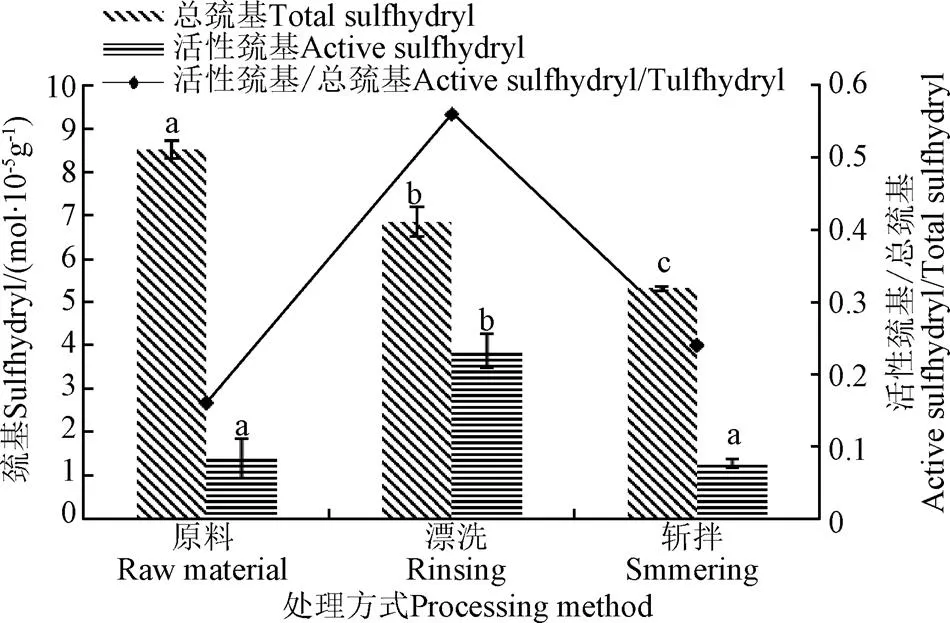
圖2 漂洗和斬拌對肌球蛋白巰基的影響
因此,可以推斷漂洗可以造成肌球蛋白變性伸展,使埋藏在蛋白質內的巰基暴露,從而活性巰基增加,并且部分被氧化形成二硫鍵,造成總巰基含量降低。漂洗魚糜經過斬拌后肌球蛋白活性巰基和總巰基都顯著降低(<0.05),這主要是因為隨著斬拌的進行,變性蛋白質會聚集形成微凝膠,而蛋白質聚合體的形成會掩蓋一些巰基,從而使檢測到的活性巰基顯著降低,總巰基含量的降低與二硫鍵的形成有關,而二硫鍵的形成有利于蛋白質凝膠的穩定。這一結果也與活性巰基與總巰基的比值研究結果一致,肌球蛋白在漂洗時解折疊度最大,變性最明顯,而經過斬拌肌球蛋白解折疊度顯著降低。另外巰基的氧化也會導致肌球蛋白表面疏水性的增加,變性程度增強,因此有必要進一步研究漂洗和斬拌對肌球蛋白表面疏水性的影響。
2.3 表面疏水性
蛋白質的表面疏水性是用來反映蛋白質分子內部疏水基團的暴露程度,一般情況下,這些疏水氨基酸殘基主要分布在蛋白質分子內部,并對肌球蛋白尾部的雙螺旋結構起著穩定的作用,在蛋白質變性時,分布在蛋白質內部的疏水基團就會暴露,并且疏水基團暴露越多,肌球蛋白的表面疏水性越大,因此通常將表面疏水性作為研究肌球蛋白的變性程度的另外一個指標。漂洗和斬拌對肌球蛋白表面疏水性的影響如圖3,漂洗后,魚糜肌球蛋白表面疏水性顯著增大,從395增加到759 S0-ANS,增加了47.9%,證明漂洗可以使肌球蛋白內部疏水氨基酸殘基暴露,肌球蛋白變性程度較大,但是有研究表明,0.5%鹽水漂洗,會使高白鮭魚肌原纖維蛋白表面疏水性顯著降低[24],因此可以推測造成肌球蛋白表面疏水性顯著增大是由于冷水漂洗的結果。而斬拌魚糜肌球蛋白相比漂洗魚糜,表面疏水性僅增加5%,差異不顯著,這主要是由于隨著斬拌的進行,變性展開的蛋白質會發生部分聚集,抑制了蛋白結構內疏水基團的暴露[25],與巰基研究結果一致。

圖3 漂洗和斬拌對肌球蛋白表面疏水性的影響
2.4 濁 度
通常外界環境的變化會改變蛋白質表面的微環境,從而造成蛋白質構象的改變,一般是蛋白質二級結構的轉變,-螺旋的解折疊也是肌球蛋白聚集的原因,蛋白聚集后,蛋白質顆粒增大,使光發生散射,引起溶液濁度增加。溶液的濁度與溶液中包含的單個的粒子所造成的渾濁程度有關,粒子包括懸浮物以及不溶解的顆粒,與溶液的透過率成反比關系。由圖4知,漂洗和斬拌均使肌球蛋白濁度增大,相比原料,漂洗后魚糜肌球蛋白濁度增大了12.5%,漂洗魚糜在經過斬拌后,肌球蛋白濁度增大了22.2%,而濁度反映的是肌球蛋白顆粒的大小,可以粗略估計肌球蛋白的聚集,根據前文巰基及表面疏水性分析知,漂洗和斬拌均會造成蛋白質的不同程度的變性和聚集,當肌球蛋白發生變性時,蛋白質分子之間逐漸發生了肽鏈的展開、交聯、聚集、絮凝等變化,從而使體系濁度逐漸上升[26],因此斬拌對肌球蛋白濁度的影響相比漂洗的更大,即斬拌更易造成肌球蛋白的聚集。
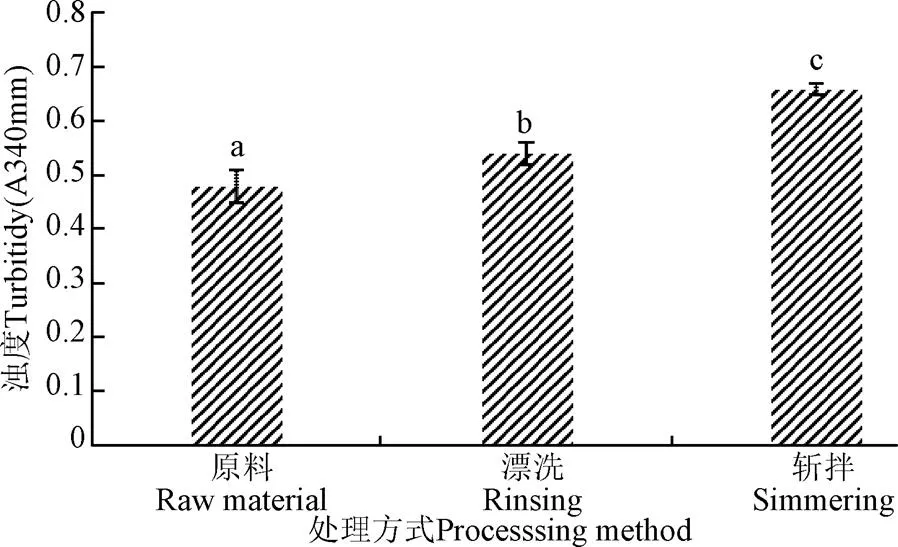
圖4 漂洗和斬拌對肌球蛋白濁度的影響
2.5 紅外光譜測定結果
由于在蛋白質中,表面疏水性和活性巰基含量的改變都意味著蛋白質二級結構可能發生了變化[27],因此為進一步研究漂洗和斬拌對肌球蛋白變性和聚集的影響,有必要對其二級結構變化進行研究。傅里葉紅外光譜在測定蛋白質二級結構中發揮著重要的作用,蛋白內部不同的結構會選擇性吸收不同的紅外波長,從而產生不同的特征吸收峰。如圖5,是不同處理條件下海鱸肌球蛋白的紅外光譜圖。傅里葉變換紅外光譜圖可提供蛋白質酰胺I帶、酰胺II帶、酰胺III帶信息以及蛋白質結構中的C-C振動和C=O等振動信息,酰胺I帶吸收峰是由蛋白多肽骨架的伸縮振動而引起的,位于之間1 700~1 600 cm-1波段之間,對蛋白質二級結構變化較敏感,通常用于分析蛋白質二級結構相對含量的變化。對酰胺I帶去卷積,進行高斯擬合得到各個子峰,其中各子峰與蛋白質二級結構的對應關系為:1 600~1 640 cm-1是-折疊;1 640~1 650 cm-1是無規卷曲;1 650~1 660 cm-1是-螺旋;1 660~1 695 cm-1是-轉角。利用相對面積變化來計算蛋白質二級結構相對含量的變化,漂洗和斬拌對肌球蛋白二級結構相對含量的變化如圖6所示。
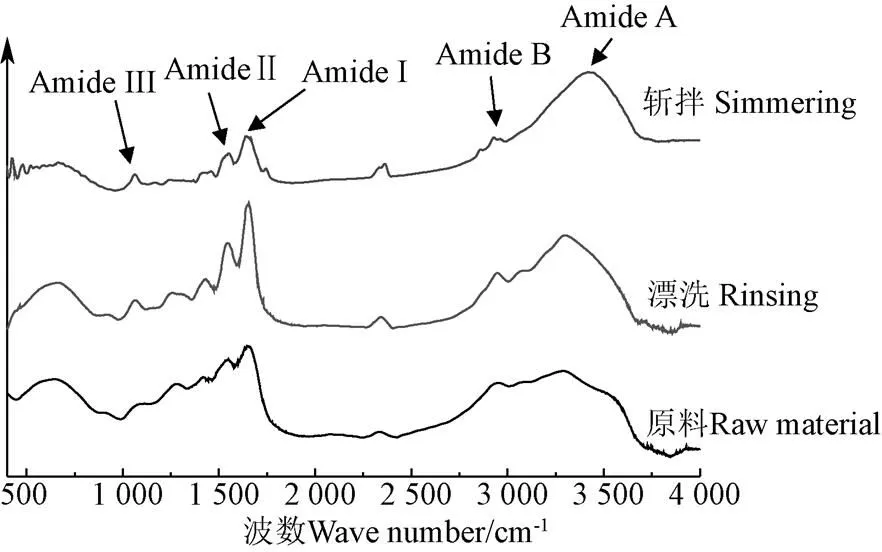
圖5 肌球蛋白紅外光譜圖
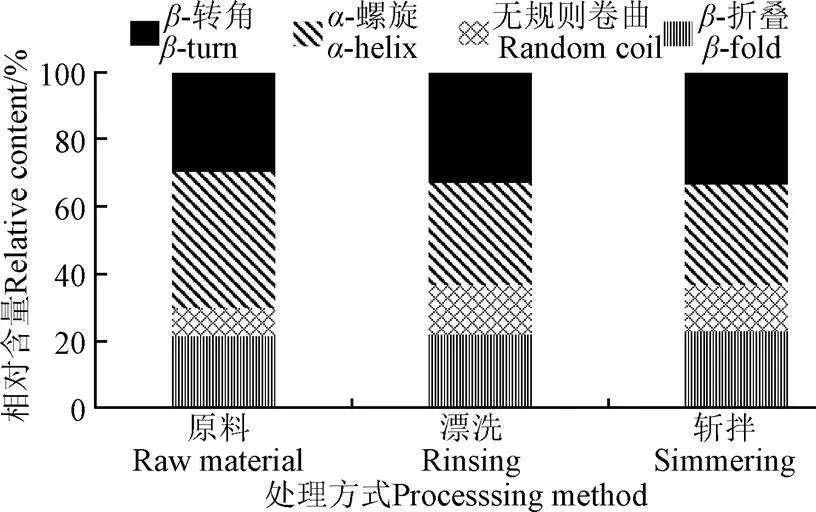
圖6 肌球蛋白二級結構相對含量變化
不同處理方式對肌球蛋白二級結構相對含量的影響如圖6,-螺旋和-折疊是主要的二級結構,又因為-螺旋和-折疊中含有較多的氫鍵,是較有序、規則的結構,因此可以推測魚肉經過漂洗和斬拌處理后,肌球蛋白結構仍相對穩定。相比原料,漂洗魚糜肌球蛋白-螺旋相對含量從40.20%降低到30.19%,降低了33.16%,無規則卷曲相對含量從8.41%增加到15.09%,增加了79.42%,-折疊和-轉角變化率相對較小,分別增加1.11%和10.38%;斬拌對肌球蛋白二級結構的影響相比于漂洗較不明顯,-螺旋和無規則卷曲相對含量分別下降1.16%和7.95%,-折疊和-轉角分別增加4.16%和1.92%,并且-折疊和-轉角結構是凝膠形成的基礎,兩者含量的增高有利于良好凝膠結構的形成[28],同時-螺旋含量的下降有利于蛋白質的聚集,肌球蛋白作為唯一可以單獨加熱形成凝膠的蛋白,對蛋白質凝膠的形成起著至關重要的作用,因此漂洗和斬拌對于蛋白質的凝膠的形成有著重要的作用。
2.6 漂洗和斬拌對肌球蛋白表面形貌的影響
2.6.1 緩沖液AFM成像
圖7為緩沖液即肌球蛋白溶解液(pH值為7.0,0.6 mol/L NaCl的20 mmol/L Tris-HCl緩沖液),由圖可知,緩沖液表面無明顯雜質,證明在超凈臺自然干燥時未被塵埃污染,其對應的3維圖進一步說明云母片表面較為平整,溶液顆粒細小,無直徑較大的聚集體。

圖7 緩沖液AFM圖像
2.6.2 不同處理條件下肌球蛋白AFM圖像
AFM圖像可以提供蛋白質表面形態的高分辨率成像,這是研究肌球蛋白聚集特征的重要因素,通過2D圖像、3D圖像和橫截面圖像觀察海鱸魚原料以及經過漂洗和斬拌的魚糜的肌球蛋白聚集體的表面形態(圖8)。
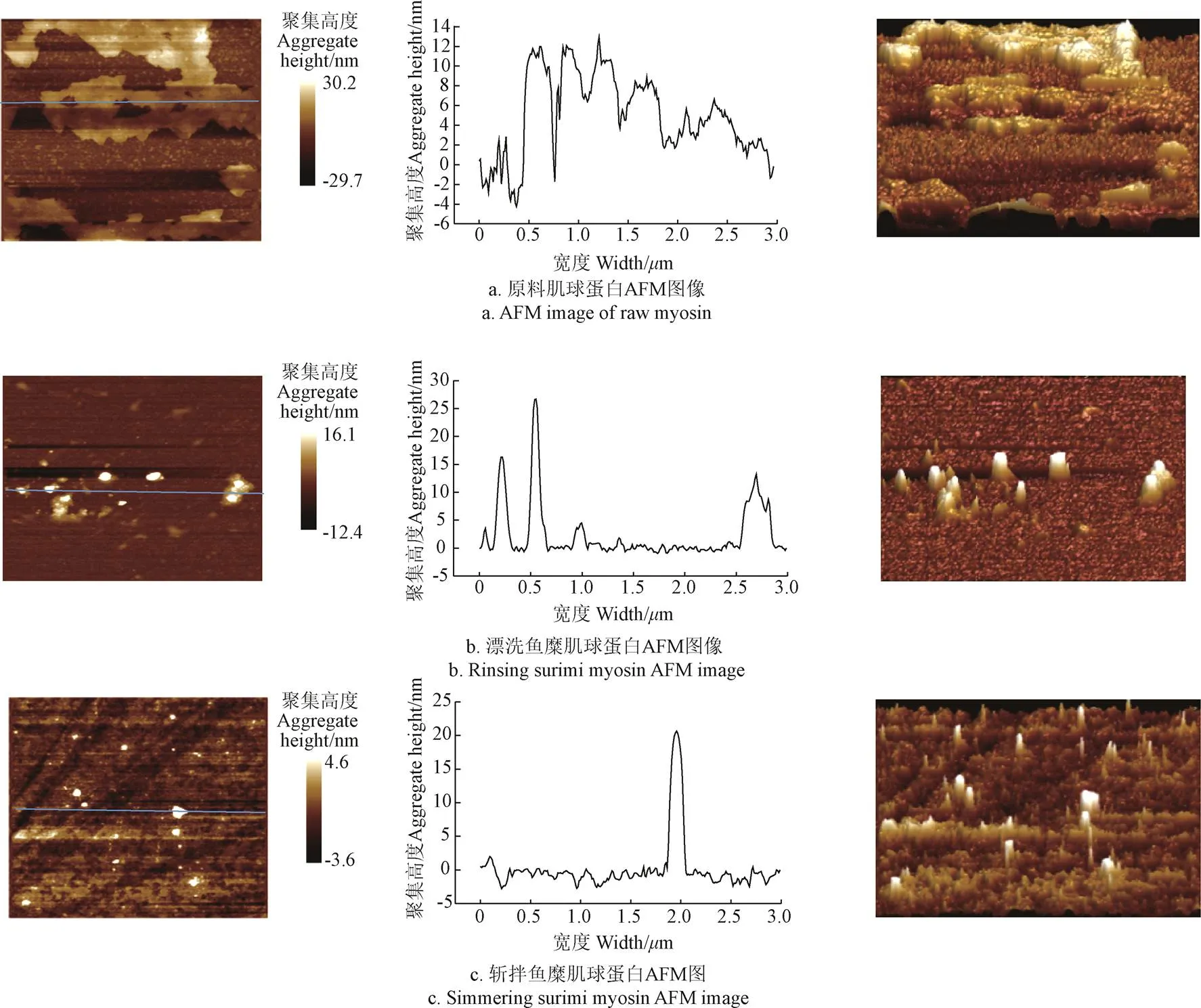
注:從左至右依次為:2D圖、左圖沿線中相應的截面分析、3D圖。 Note: From left to right are 2D image, section analyses along the corresponding lines, 3D graph.
已有研究證明,肌球蛋白在生理條件下呈現單體或可溶性低聚物[29],由圖8a可知,海鱸魚原料在未經任何處理的條件下表現出完整的肌球蛋白束,并且肌球蛋白在懸浮液中的均勻分布,無明顯顆粒狀存在,與雞胸肌肌原纖維蛋白外觀一致[30]。然而王艷敏等[31]研究表明,未加熱的鰱魚天然肌球蛋白以單體或小聚集體的形式吸附在云母片的表面,Guo等[32]研究表明,金線魚肌球蛋白在天然狀態下呈現珠狀,因此可以推測不同魚種肌球蛋白在天然狀態下結構有所差異。蛋白在漂洗(圖8b)和斬拌(圖8c)后,肌球蛋白開始以低聚物的形式存在,通過高度圖像的截面分析,肌球蛋白懸浮液中聚集體的高度從14 nm經過漂洗明顯增高到高于25 nm,斬拌后,肌球蛋白聚集體高度略有下降,但不明顯,如圖8所示。另外經過斬拌,肌球蛋白顆粒聚集簇明顯減小,且分布更加均勻。因此在漂洗和斬拌過程中,肌球蛋白是變性和聚集共同存在的過程,又因為蛋白質解聚是加熱進一步聚集成簇狀聚合物的前提,通過原子力顯微鏡的研究,進一步證實了漂洗和斬拌是蛋白質加熱形成具有良好凝膠性能的重要前處理步驟。
3 結 論
漂洗和斬拌作為魚糜加工過程中的重要步驟,相比原料,漂洗魚糜肌球蛋白總巰基減少了19.5%,活性巰基、濁度和表面疏水性分別增大了63.9%、12.5%和47.9%,肌球蛋白變性程度較大;而斬拌過程中活性巰基減少了66.8%,肌球蛋白變性程度減弱,漂洗對表面疏水性影響更大,斬拌對濁度影響更大;漂洗對魚糜肌球蛋白二級結構影響更顯著,尤其是對-螺旋和無規則卷曲結構,-螺旋相對含量降低了33.16%,無規則卷曲相對含量增加了79.42%,斬拌后肌球蛋白二級結構變化相對不明顯;漂洗和斬拌均可以造成肌球蛋白表面形貌發生變化,漂洗后,肌球蛋白聚集簇明顯減小,而聚集高度顯著增大,斬拌后肌球蛋白聚集簇進一步減小,聚集高度變化不明顯。因此,漂洗和斬拌對肌球蛋白的基本理化性質以及變性和聚集有很大的影響,是魚糜加工過程中重要的前處理過程。
[1]Xu X L, Han M Y, Fei Y, et al. Raman spectroscopic study of heat-induced gelation of pork myofibrillar proteins and its relationship with textural characteristic[J]. Meat Science, 2011, 87(3): 159-164.
[2]Samejima K, Hashimoto Y, Yasui T, et al. Heat gelling properties of myosin, actin, actomyosin and myosin-subunits in a saline model system[J]. Journal of Food Science, 2010, 34(3): 242-245.
[3]Samejima K, Hashimoto Y, Yasui T, et al. Heat gelling properties of myosinactin, actomyosin and myosin-subunits in a saline model system[J]. on Academic, 1969, 34(3): 242-245.
[4]Hayakawa T, Yoshida Y, Yasui M, et al. Heat-induced gelation of myosin in a low ionic strength solution containing L-histidine[J]. Meat Science, 2012, 90(1): 77-80.
[5]高霞,曹立偉,熊善柏,等. 不同質量濃度鰱魚肌球蛋白低溫自組裝動力學及理化性質[J]. 食品科學,2019,40(1):1-8.
Gao Xia, Cao Liwei, Xiong Shanbai, et al. Self-assembly kinetics and physicochemical properties of silver carp myosin under conditions with different concentrations and low temperature[J]. Food Science, 2019, 40(1): 1-8. (in Chinese with English abstract)
[6]Brenner T, Johannsson R, Nicolai T. Characterization of fish myosin aggregates using static and dynamic light scattering[J]. Food Hydrocolloids, 2009, 23(2): 296-305.
[7]Chan J K, Gill T A, Thompson J W, et al. Herring surimi during low temperature setting, physicochemical and textural properties[J]. Journal of Food Science, 1995, 60(6): 1248-1253.
[8]Gill T A, Chan J K, Phonchareon K F, et al. Effect of salt concentration and temperature on heat-induced aggregation and gelation of fish myosin[J]. Food Research International, 1992, 25(5): 333-341.
[9]Ding Y, Liu R, Rong J, et al. Heat-induced denaturation and aggregation of actomyosin and myosin from yellowcheek carp during setting[J]. Food Chemistry, 2014, 149: 237-243.
[10]王金余,劉承初,趙善貞,等. 白鰱魚糜肌球蛋白交聯反應和凝膠化最適條件的研究[J]. 食品科學,2008,29(11):223-227.
Wang Jinyu, Liu Chengchu, Zhao Shanzhen, et al. Optimal conditions for maximal cross-linkage of myosin heavy chain (MHC) and gelation of surimi product from silver carp (Hypophthalmichtys molitrix)[J]. Food Science, 2008, 29(11): 223-227. (in Chinese with English abstract)
[11]Ducept F, De Broucker T, Souliè J M, et al. Influence of the mixing process on surimi seafood paste properties and structure[J]. Journal of Food Engineering, 2012, 108(4): 557-562.
[12]李賢. 破碎方式對魚糜凝膠特性的影響[D]. 武漢:華中農業大學,2014.
Li Xian. Effect of Broken Methods on Surimi Gel Properties[D]. Wuhan: Huazhong Agricultural University, 2014. (in Chinese with English abstract)
[13]Ikeda S, Morris V J, Nishinari K. Microstructure of aggregated and nonaggregated κ-carrageenan helices visualized by atomic force microscopy[J]. Biomacromolecules, 2001, 2(4): 1331-1337.
[14]Park J, Lanier T. Scanning calorimetric behavior of tilapia myosin and actin due to processing of muscle and protein purification[J]. Journal of Food Science, 1989, 54: 49-51.
[15]Cao H, Jiao X, Fan D, et al. Microwave irradiation promotes aggregation behavior of myosin through conformation changes[J]. Food Hydrocolloids, 2019, 96: 11-19.
[16]Wei L, Cao L, Xiong S, et al. Effects of pH on self-assembly of silver carp myosin at low temperature[J]. Food Bioscience, 2019, 30: 100420.
[17]Yongsawatdigul J, Park J W. Thermal denaturation and aggregation of threadfin bream actomyosin[J]. Food Chemistry, 2003, 83(3): 409-416.
[18]林婉玲,楊賢慶,李來好,等. 浸漬凍結對調理草魚凍藏過程中肌原纖維蛋白特性的影響[J]. 南方水產科學,2016,12(3):67-73.
Lin Wanling, Yang Xianqing, Li Laihao, et al. Immersion freezing effect on myofibrillar protein characteristics of prepared grass carp () fillets during frozen storage[J]. South China Fisheries Science, 2016, 12(3): 67-73. (in Chinese with English abstract)
[19]He X H, Liu H Z, Liu L, et al. Effects of high pressure on the physicochemical and functional properties of peanut protein isolates[J]. Food Hydrocolloids, 2014, 36: 123-129.
[20]Benjakul S, Sutthipan N. Muscle changes in hard and soft shell crabs during frozen storage[J]. LWT- Food Science and Technology, 2009, 42(3): 723-729.
[21]Hamada M, Ishizaki S, Nagai T. Variation of SH content and kamaboko-gel forming ability of shark muscle protein by electrolysis[J]. Journal of Shimonoseki University of Fisheries, 1994, 42: 131-135.
[22]Ko W C, Yu C C, Hsu K C. Changes in conformation and sulfhydryl groups of tilapia actomyosin by thermal treatment[J]. LWT-Food Science and Technology, 2007, 40(8): 1316-1320.
[23]馮芳,劉文豪,陳志剛. 大豆7S、11S蛋白的結構與熱致凝膠特性的研究[J/OL].食品科學:1-11[2019-07-03]. http://kns.cnki.net/kcms/detail/11.2206.ts.20190409.1521.064. html.
[24]高澤磊. 防止蛋白質氧化引起高白鮭肌肉劣變的技術研究[D]. 石河子:石河子大學,2017.
Gao ZeLei. Study on the Technology to Prevent Protein Oxidation Caused the Deterioration of Coregonus Peled Muscle[D]. Shihezi: Shihezi University, 2017. (in Chinese with English abstract)
[25]李婷,周春霞,馮瑞,等. 硫酸葡聚糖對低離子強度下羅非魚肌球蛋白熱變性聚集的抑制及其機制[J]. 食品與機械,2018,34(12):5-10.
Li Ting, Zhou Chunxia, Feng Rui, et al. Inhibition of heat-induced aggregation of Tilapia myosin by dextran sulfate at low ionic strength[J]. Food & Machinery, 2018, 34(12): 5-10. (in Chinese with English abstract)
[26]姜啟興. 鳙魚肉熱加工特性及其機理研究[D]. 無錫:江南大學,2015.
Jiang Qixing. Study on Thermal Processing Properties and Mechanism of Bighead Carp() Muscle[D]. Wuxi: Jiangnan University, 2015.
[27]Chen H H, Xu S Y, Wang Z. Interaction between flaxseed gum and meat protein[J]. Journal of Food Engineering, 2007, 80(4): 1051-1059.
[28]Kang Z L, Wang P, Xu X L, et al. Effect of beating processing, as a means of reducing salt content in frankfurters: A physico-chemical and Raman spectroscopic study[J]. Meat Science, 2014, 98(2): 171-177.
[29]Shimada M, Takai E, Ejima D, et al. Heat-induced formation of myosin oligomer-soluble filament complex in high-salt solution[J]. International Journal of Biological Macromolecules, 2015, 73: 17-22.
[30]Chen X, Xu X, Zhou G. Potential of high pressure homogenization to solubilize chicken breast myofibrillar proteins in water[J]. Innovative Food Science & Emerging Technologies, 2016, 33: 170-179.
[31]王艷敏,于加美,石彤,等. 原子力顯微鏡研究L-半胱氨酸對鰱肌球蛋白溶液熱聚集行為的影響[J]. 食品科學,2018,39(14):1-8.
Wang Yanmin, Yu Jiamei, Shi Tong, et al. Influence of l-cysteine on the heat-induced aggregation of bighead carp myosin studied by atom force microscope[J]. Food Science, 2018, 39(14): 1-8. (in Chinese with English abstract)
[32]Guo Z, Li Z, Wang J, et al. Gelation properties and thermal gelling mechanism of golden threadfin bream myosin containing CaCl2 induced by high pressure processing[J]. Food Hydrocolloids, 2019, 95: 43-52.
Effects of rinsing and mashing on physical and chemical properties of myosin in sea bass
Lin Wanling1,2, Liu Fangfang1,3, Li Laihao1※, Wu Yanyan1, Yang Shaoling1, Huang Hui1, Yang Xianqing1, Lin Zhi4
(1.,,510300; 2.,521041,; 3.,,201306; 4..,.,529800,)
The physicochemical properties, such as total sulfhydryl (T-SH) group, active sulfhydryl (R-SH) group, surface hydrophobicity and turbidity, were determined for the myosin that extracted separately from the raw materials, rinsed- and simmer surimi in sea bass, while the secondary structure and surface morphology of the myosin were analyzed using the combined techniques including the infrared spectroscopy and atomic force microscopy (AFM). The experimental results showed that the content of total sulfhydryl (T-SH) groups in the myosin of the rinsed surimi decreased by 19.5% from 8.51 to 6.85 mol/105g, whereas the active sulfhydryl group increased by 63.9% from 1.39 to 3.86 mol/105g, compared with the raw materials. The content of total sulfhydryl (T-SH) and active sulfhydryl (R-SH) groups in the myosin of the simmer surimi decreased to 5.3 and 1.28 mol/105g compared with the raw materials, indicating decrease by 22.6% and 66.8%, respectively. The ratio of the active sulfhydryl (R-SH) to the total sulfhydryl (T-SH) group in the myosin of the rinsed surimi is the largest than others, inferring that the myosin of the rinsed surimi can be the most deformed in this case. The turbidity and surface hydrophobicity, the indictor for the aggregation and degree of denaturation in proteins, in the myosin of the surimi increased after the treatment of rinsing and simmering. Specifically, the turbidity of the myosin in the surimi increased by 12.5% after rinsing and 22.2% after simmering, compared with raw materials , whereas the surface hydrophobicity increased by 47.9% after rinsing and 5% after simmering. The data reveal that the treatment of the rinsing and simmering can significantly affect the denaturation and aggregation of proteins in the varying level. The rinsing has a greater influence on the surface hydrophobicity, while the mixing effect has a greater influence on the turbidity. The simmering can cause the slight aggregation of the myosin to form a sort of microgel. There was a much more obvious effect of the rinsing on the secondary structure of the myosin. In Fourier transform of the infrared spectroscopy the-helix relative content of the myosin that extracted can decreased by 33.16%, while the random curl relative content increased by 79.42%,-fold and-turn increased by 1.11% and 10.38%, respectively, after the rinsing of raw materials in the sea bream. After rinsing, the change rate of the secondary structure in the surimi myosin was smaller, and the-helix and random curl content decreased by 1.16% and 7.95%,-fold and-turn increased by 4.16% and 1.92%, respectively. Generally, the decrease in the-helix content can be conducive to the aggregation of proteins, and the increase in-sheet and-turn can be beneficial to the formation of the good gel structure. The characterized results from atomic force microscopy (FAM) showed that the myelin myosin exhibited a sort of protein bundle under the natural conditions, and the distribution of the protein bundle in the suspension was uniform without obvious granularity. Both treatment of the rinsing and simmering can change the surface morphology of the myosin in the sea bass, indicating reduce significantly the aggregation clusters of the myosin, while increase the height of the aggregates in the myosin suspension from 14 nm to 25 nm. After mixing, the height of the myosin aggregates did not change significantly. These findings demonstrate that the treatment of the rinsing and simmering have a great influence on the physicochemical properties of the myosin in rinsed and simmer surimi of the sea bass, as well as a certain effect on the degeneration and aggregation of the myosin suspension. This study can be expected to provide potential reference for the future development on the formation mechanism of surimi gel in food industry.
protein; fish; monitoring; myosin; degeneration and aggregation; secondary structure; surface morphology
林婉玲,劉芳芳,李來好,吳燕燕,楊少玲,黃 卉,楊賢慶,林 織. 漂洗和斬拌對海鱸魚肌球蛋白理化特性的影響[J]. 農業工程學報,2020,36(1):310-317.doi:10.11975/j.issn.1002-6819.2020.01.037 http://www.tcsae.org
Lin Wanling, Liu Fangfang, Li Laihao, Wu Yanyan, Yang Shaoling, HuangHui, Yang Xianqing, Lin Zhi. Effects of rinsing and mashing on physical and chemical properties of myosin in sea bass[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2020, 36(1): 310-317. (in Chinese with English abstract) doi:10.11975/j.issn.1002-6819.2020.01.037 http://www.tcsae.org
2019-08-20
2019-12-25
“十二五”國家重點研發計劃(2016YFD0400201-6);中央級公益性科研院所基本科研業務費專項資金項目(2019TS16);現代農業產業技術體系專項(CARS-47);“揚帆計劃”引進創新創業團隊專項(2015YT02H109)
林婉玲,博士,副研究員,研究方向為水產品加工與質量安全研究。Email:lwlscsf@163.com
李來好,博士,研究員,研究方向為水產品加工與質量安全。Email:laihaoli@163.com
10.11975/j.issn.1002-6819.2020.01.037
TS254.4
A
1002-6819(2020)-01-0310-08
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
