不同養殖模式對黑鯛生長、血清生化指標及抗氧化性能的影響
2020-02-22 05:41:08王裕玉徐跑張志偉張志勇聶志娟吳建平曾海峰徐鋼春
江蘇農業科學 2020年23期
王裕玉 徐跑 張志偉 張志勇 聶志娟 吳建平 曾海峰 徐鋼春

摘要:比較池塘流水槽養殖和傳統池塘養殖模式下黑鯛幼魚(初始平均體質量為5.29 g)生長、血清生化指標及抗氧化能力。60 d的養殖結果表明,流水槽養殖組黑鯛增質量率、特定生長率均顯著高于池塘養殖組(P<0.05);血清中谷草轉氨酶活性、谷丙轉氨酶活性、總蛋白含量、總膽固醇含量、皮質醇含量、溶菌酶含量和超氧化物歧化酶在處理組之間無顯著性差異;流水槽養殖組血清中堿性磷酸酶活性顯著高于池塘養殖組,而血糖和甘油三酯含量正好相反;流水槽養殖組肝臟谷胱甘肽過氧化物酶活性、總超氧化物歧化酶活性、總抗氧化能力均顯著高于池塘養殖組,而過氧化氫酶活性和丙二醛含量無顯著性差異。總之,在本試驗條件下,池塘流水槽養殖模式能提高黑鯛(體質量為5~37 g)的生長和機體免疫力,降低肌肉脂肪含量。
關鍵詞:池塘流水槽循環水養殖;黑鯛;養殖模式;血清生化指標;抗氧化
中圖分類號: S965.231 ?文獻標志碼: A ?文章編號:1002-1302(2020)23-0155-05
池塘工程化循環水生態養殖,別稱為池塘流水槽循環水養殖,是我國漁業轉型升級、創新發展的一種養殖新模式,具有節水節地、高產量、低漁藥使用量、污物資源化利用、肉質緊實美味、養殖全程可控等優點,已被列入《農業綠色發展技術導則(2018—2030年)》,成為農業農村部主推的健康養殖技術之一。目前有關池塘工程化循環水養殖模式的研究主要集中在系統建設優化、淡水養殖品種篩選等方面[1],而關于海水養殖魚類生長適應情況及配套技術的研究尚未見報道。研究表明,養殖模式能影響魚類的生長性能、肌肉品質、免疫性能、養殖水質以及經濟效益等[2-5]。黑鯛(Acanthopagrus schlegelii),別稱烏頰魚、黑立、黑加吉等,隸屬鱸形目(Perciformes),鯛科(Sparidae)棘鯛屬(Acanthopagrus),為暖溫性底層魚類,主要分布在太平洋西部沿海,如日本、朝鮮半島南部和東南亞國家附近海域等,在我國主要分布于渤海、黃海、東海沿岸以及臺灣海峽南部水域等,是我國重要的海水養殖的經濟魚類之一,因其具有生長快、抗病力強、適鹽性和適溫性廣、肉味鮮美、營養價值高等優點,深受廣大養殖戶和消費者的喜愛。本試驗比較池塘工程化循環水養殖和傳統池塘養殖模式下黑鯛生長、血清生化及抗氧化指標的影響,以期為黑鯛工程化循環水養殖技術的推廣與示范提供參考依據。
1 材料與方法
1.1 試驗條件
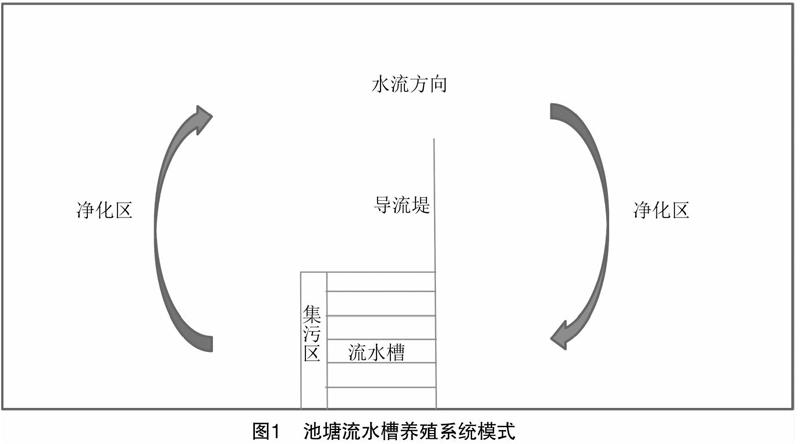
養殖試驗在江蘇省南通市通州灣漁業發展有限公司的6條流水槽(磚混結構)內進行。流水槽由氣提推水增氧區、養殖區(20 m×5 m×2 m)和集排污區(30 m×5 m)組成(圖1)。流水槽總面積為 600 m2,占池塘總面積的1.12%,剩余池塘面積為水質凈化區,凈化區內放養牡蠣和文蛤等貝類用于水質凈化和增加養殖效益。傳統池塘面積為 1 334 m2,水深為2.5 m。海水水質清新、無污染,符合《漁業水質標準》。
1.2 試驗魚養殖管理
黑鯛苗種為江蘇省海洋水產研究所海水增養殖
技術及種苗中心自主培育的苗種。試驗開始前,將規格一致、體質健康的魚苗(初始平均體質量為5.29 g)隨機分組,流水槽養殖密度為15 000尾/槽,池塘放養密度為3 000尾/0.067 hm2,每個處理組3個平行。試驗期間,2種養殖模式均投喂浮性顆粒飼料[飼料粗蛋白含量≥40%,購自天邦食品股份(寧波)有限公司]。投喂頻率均為每天3次(08:00、13:00、17:00),日投喂量均為體質量的2%~4%,具體投喂量根據水溫、天氣變化、生理狀況等適時調整。投喂飼料時確保飼料不漂出流水槽,每次投飼時長為30 min左右,以上浮搶食魚數量明顯減少時停止投喂。試驗期間,水體鹽度為25,水溫為24.0~33.8 ℃,溶解氧含量為5.2 mg/L,pH值為8.3,流水槽水流速度為2~4 cm/s。養殖試驗于2018年6月3日開始,于2018年8月1日結束,持續60 d。
1.3 樣品采集
采樣前試驗魚饑餓24 h。每槽隨機取20尾魚,用200 mg/L間氨基苯甲酸乙酯甲磺酸鹽(MS-222)將其麻醉,測量體質量和體長等指標。尾靜脈取血,在溫度為4 ℃條件下靜置 4 h,4 000 r/min離心15 min,吸取血清于-80 ℃條件下保存待測。采血完成后,迅速解剖取肝臟、肌肉,液氮中速凍,轉入-80 ℃保存,用于抗氧化指標和肌肉成分分析。
增質量率(WG)、特定生長率(SGR)、肥滿度(CF)、肝體指數(HSI)和臟體指數(VSI)計算公式如下:
1.4 樣品分析
血清生化指標采用全自動生化分析儀(邁瑞 BS-400,深圳)進行測定,包括總蛋白(total protein,簡稱TP)含量、谷草轉氨酶(aspartate aminotransferase,簡稱AST)活性、谷丙轉氨酶(alanine aminotransferase,簡稱ALT)活性、堿性磷酸酶(alkaline phosphatase,簡稱ALP)活性、血糖(glucose,簡稱GLU)含量、總膽固醇(total cholesterol,簡稱TC)含量、甘油三酯(triglyceride,簡稱TG)含量等;采取酶聯免疫檢測法(enzyme-linked immunosorbent assay,簡稱ELISA)測定血清超氧化物歧化酶(superoxide dismutase,簡稱SOD)活性、皮質醇和溶菌酶含量。
將肝臟樣品解凍,準確稱量肝臟質量,并加入9倍體積預冷的0.86%生理鹽水,勻漿,勻漿液經 3 000 r/min 離心(4 ℃) 10 min,取上清液,在 -80 ℃ 條件下保存用于抗氧化酶活性測定。抗氧化酶活性采用酶標儀(Synergy H1,Bio-Tek,美國)進行測定,包括過氧化氫酶(catalase from micrococcus lysodeiktic,簡稱CAT)活性、谷胱甘肽過氧化物酶(glutathione peroxidase,簡稱GSH-PX)活性、總抗氧化能力(total antioxidant capacity,簡稱 T-AOC)、總超氧化物歧化酶(T-SOD)活性及丙二醛(malondialdehyde,簡稱MDA)含量,均采用南京建成生物工程研究所生產的試劑盒進行測定。肝臟蛋白含量采用考馬斯亮藍法進行測定。
1.5 統計分析
采用SAS 9.12對數據進行統計學分析。先作單因素方差分析(one-ANOVA),若處理組之間差異顯著,再作Duncans多重比較,P<0.05表示處理間在0.05水平上差異顯著。試驗結果采用“平均值±標準差”表示。
2 結果與分析
2.1 不同養殖模式對黑鯛生長性能的影響
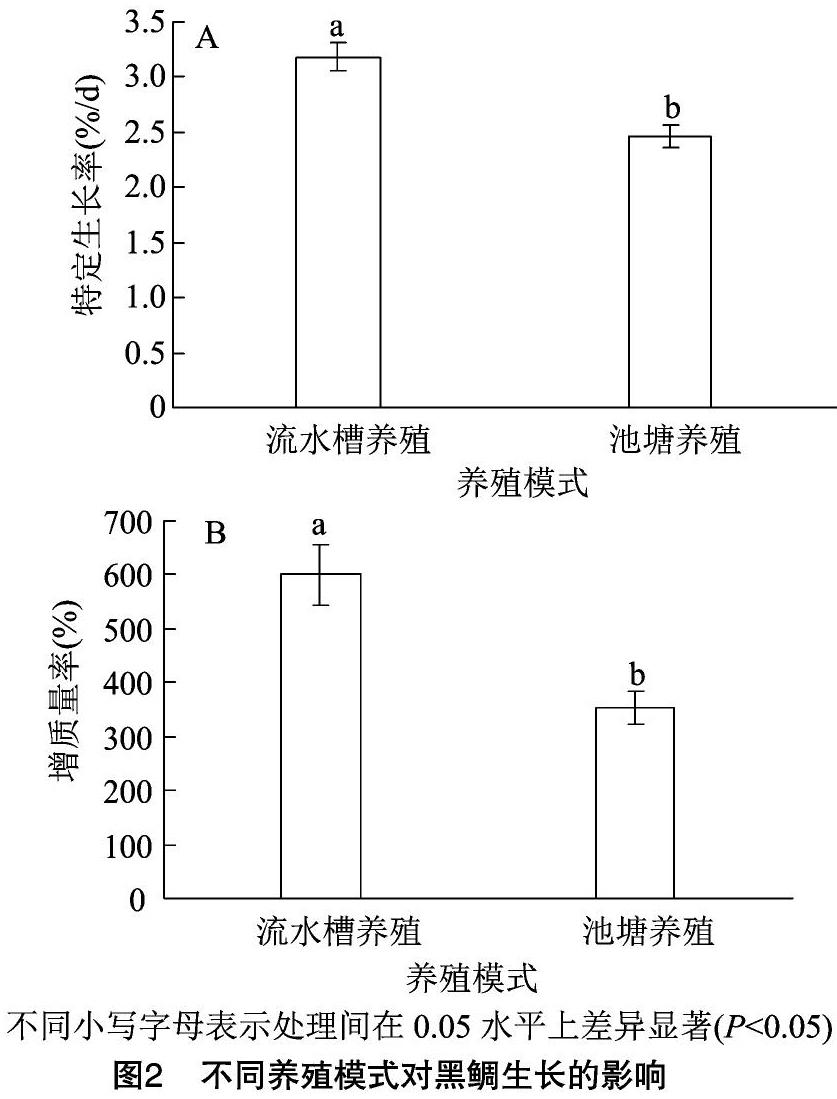
60 d的養殖結果表明,流水槽養殖組黑鯛終增質量率和特定生長率均顯著高于池塘養殖組(圖2)。
黑鯛體長和體質量的關系如圖3所示,將試驗數據擬合,得到流水槽養殖和池塘養殖黑鯛體質量(y)與體長(x)均呈冪函數關系,擬合方程分別為y流=0.069 3x2.718 2(r2=0.958 3);y池=0.049 3x2.745 6(r2=0.965 5)。冪指數均接近3,表明黑鯛為近似等速生長類型。
2.2 血清生化指標
由表1可知,不同養殖模式對血清總蛋白含量、ALY活性、AST活性、總膽固醇含量、超氧化物歧化酶活性、皮質醇含量和溶菌酶含量均無顯著影響。流水槽養殖組黑鯛血清堿性磷酸酶活性顯著高于池塘養殖組,而血糖和甘油三酯含量顯著低于池塘養殖組。
2.3 抗氧化酶活性
由表2可知,流水槽養殖組肝臟谷胱甘肽過氧化物酶活性、總超氧化物歧化酶活性、總抗氧化力均顯著高于池塘養殖組,而過氧化氫酶活性和丙二醛含量在組間無顯著性差異。
2.4 體組成與形體指數
由表3可知,不同養殖模式對肌肉水分、蛋白質和灰分無顯著性影響;而流水槽養殖組黑鯛肌肉脂肪含量顯著低于池塘養殖組。由表4可知,流水槽養殖組黑鯛肥滿度、肝體指數和臟體指數均顯著高于池塘養殖組。
3 討論
3.1 養殖模式對黑鯛生長的影響
黑鯛具有集群搶食的特性,攝食能力較強。在整個養殖試驗周期內,試驗魚很快適應了養殖環境和膨化飼料,未發生病害和明顯死亡,流水槽和池塘養殖黑鯛成活率分別為94.17%、92.24%。流水槽養殖模式將傳統池塘開放式散養模式創新為新型的圈養模式,通過構建濾水性魚類、貝類和水生植物為一體的水質凈化技術,有利于營養物質的循環利用,既可以解決養殖自身污染的根本問題,又可以做到變廢為寶,增加經濟效益。在本研究中,流水槽養殖組黑鯛增質量率和特定生長率均顯著高于池塘養殖組,這可能與持續適宜的水流有利于魚類的生長有關,流水槽中水流速度約為3 cm/s,在推薦范圍內。Ibarz等對金頭鯛(Sparus aurata)的研究結果表明,運動強度為1.5 cm/s組飼料攝入量無增加,而體質量和特定生長率顯著上升,這說明通過游泳鍛煉可以提高營養利用率,有助于改善魚類的生長和減少氮的流失[6]。類似的相關研究中,Hackbarth等對頭石脂鯉(Brycon cephalus)[7]、Brown等對黃尾鰤(Seriola lalandi)[8]和Huang等對西伯利亞鱘(Acipenser baeri)[9]的研究結果均表明,適度的游泳運動訓練(14.0~35.7 cm/s)對生長有明顯的促進作用。與之相反,虞順年等探討了水流速度(0、6.75、13.50、27.00 cm/s)對黑鯛生長的影響,結果表明,游泳訓練組的增質量率和特定生長率均顯著下降[10];Alcaraz等研究發現,與對照組相比較,水流速度為4.1、7.8、12.9 cm/s組的劍尾魚(Xiphophorus montezumae)生長率分別降低33.8%、39.3%、21.8%[11];持續游泳運動訓練會抑制金帶籃子魚(Siganus rivulatus)(13.3 cm/s)[12]和真鯛(Pagrosomus major)(3~16.8 cm/s)[13]的生長,這可解釋為魚類消耗大量的能量用于游泳運動,而用于生長的能量分配減少,從而抑制生長率,更為嚴重的會對魚體造成損傷,最終導致魚死亡。不同研究者得出不同的結論可能與魚的種類、生活習性、生長階段、養殖周期和環境條件等的差異有關。
3.2 養殖模式對黑鯛血清生化指標的影響
魚類血液學指標能夠反映機體營養、生理代謝及健康狀況等,當魚體受到外界因子的影響而發生生理或病理變化時,必定會在血液指標中反映出來,因此其通常被廣泛地用于評價魚類的營養、健康以及對環境的適應狀況,并為防治方案提供有價值的信息[14]。AST和ALT是動物體內2種重要的轉氨酶,在非必需氨基酸合成和蛋白質分解代謝中起著重要的作用,當肝臟受損時會釋放到血液中,血液中AST和ALT含量升高表明肝臟可能受到了損傷[15]。虞順年等研究表明,游泳訓練強度為 2.0 bl/s (bl為體長) 時,黑鯛血清中AST和ALT活性均顯著降低,表明在該強度下運動訓練能保護肝臟,增強肝臟的機能[10]。本研究中,流水槽養殖和池塘養殖對黑鯛血清中AST和ALT活性無顯著影響,這說明水流速度為2~4 cm/s以及池塘養殖對肝臟功能和蛋白質分解代謝無影響。血清總蛋白和ALP對魚體代謝和免疫機能有重要的作用,可反映機體蛋白質代謝水平和抵抗力等[16]。本研究中,流水槽養殖的魚血清中ALP含量顯著高于池塘養殖組,這表明逆流運動能增強機體代謝和免疫機能,這與虞順年等對黑鯛的研究結果[10]相似。研究者對黑鯛[10]和大鼠[17-18]的研究結果表明,適度的有氧運動能夠調節血清中GLU含量,降低TG、TC、低密度脂蛋白(LDL)含量。本研究結果顯示,流水槽養殖組黑鯛血清中血糖和甘油三酯含量顯著降低,表明流水槽養殖中適度的逆流運動具有降血糖和血脂的作用,這可解釋為在適度的運動刺激下魚類的代謝機能提高,促進了魚體對血脂的轉運和血糖的利用。值得注意的是,并不是水流刺激越高越好。虞順年等研究發現,當游泳訓練強度為27 cm/s時,超過魚體自身的負荷,一方面會加劇對能量的消耗,另一方面會對黑鯛機體造成損傷,引起血糖和脂質代謝功能紊亂,最終影響魚類生長與健康[10]。當養殖水環境對魚體產生脅迫效應時,會激活下丘腦-垂體-腎間軸(HPI)而產生皮質醇[19]。皮質醇含量過高時,會造成動物生長減緩、體內脂肪合成速率降低,肌肉消退,最終導致機體消瘦[20]。SOD和溶菌酶作為非特異性免疫因子,在魚類的免疫應答和抗病力方面起著重要作用[21],魚類可以通過增加溶菌酶或補體水平、提高免疫水平以應對應激[22-23]。本研究中,血清中皮質醇含量、SOD活性和溶菌酶含量在流水槽養殖和池塘養殖組之間無顯著性差異,表明這2種養殖模式未對黑鯛應激和免疫應答產生負面影響。須要注意的是,血清生化指標這些參數的波動可能與養殖季節、溫度和營養攝入等有關。
3.3 養殖模式對黑鯛抗氧化酶的影響
氧化應激在集約化水產養殖中是常見的現象,當魚體受到擁擠等應激時抗氧化系統失調,導致代謝紊亂,最終影響其生長、抗氧化力和抗病力等[24]。本試驗比較適宜養殖密度下2種模式黑鯛抗氧化酶活性的差異。結果表明,流水槽養殖組肝臟GSH-PX活性、T-SOD活性、T-AOC顯著高于池塘養殖組,這表明游泳運動訓練能增強肝臟代謝水平和免疫機能[25-26]。類似的,運動訓練對黑鯛[10]和中華倒刺鲃[27]肝臟T-AOC均有提升趨勢,但組間差異并不顯著,說明肝臟總抗氧化能力處于相對穩定的狀態。在本研究中,CAT活性在組間無顯著性差異。不同的是,虞順年等研究發現,隨著運動強度的增加,肝臟CAT活性呈上升趨勢,其中流速為 4 bl/s 試驗組顯著高于對照組,這可能是在該強度下訓練能促進黑鯛肝臟相關酶的合成或分泌,從而有利于肝臟代謝機能和抗氧化機能的提升[10]。肝臟MDA含量在組間無顯著性差異,這說明2~4 cm/s的水流速度和靜水養殖并未引發黑鯛肝臟脂質過氧化。這與虞順年等的結果[10]相似。
4 結論
綜上所述,在本試驗條件下,池塘工程化循環水養殖模式有利于提高黑鯛(體質量為5~37 g)的生長、代謝和部分免疫指標。隨著黑鯛的生長,養殖后期高養殖密度可能會引起應激反應,這有待于進一步深入研究。
參考文獻:
[1]Wang Y,Xu G C,Nie Z J,et al. Growth performance of bluntnose black bream,channel catfish,yellow catfish,and largemouth bass reared in the in-pond raceway recirculating culture system[J]. North American Journal of Aquaculture,2019,81(2):153-159.
[2]邵俊杰,張世勇,朱昱璇,等. 不同養殖模式對斑點叉尾生長和肌肉品質特性的影響[J]. 水產學報,2017,41(8):1256-1263.
[3]張 曦,李 天,李小兵,等. 不同養殖模式對尼羅羅非魚生長、免疫性能及水質的影響[J]. 水生態學雜志,2017,38(4):65-70.
[4]廖 凱,張英麗,楊正勇,等. 中國花鱸不同養殖模式經濟效益比較研究[J]. 上海海洋大學學報,2018,27(4):508-514.
[5]高露姣,黃艷青,夏連軍,等. 不同養殖模式下紅鰭東方鲀的品質比較[J]. 水產學報,2011,35(11):1668-1676.
[6]Ibarz A,Felip O,Fernández-Borràs J,et al. Sustained swimming improves muscle growth and cellularity in gilthead sea bream[J]. Journal of Comparative Physiology B,2011,181(2):209-217.
[7]Hackbarth A,Moraes G.Biochemical responses of matrinx Brycon cephalus (Günther,1869) after sustained swimming[J]. Aquaculture Research,2010,37(11):1070-1078.
[8]Brown E J,Bruce M,Pether S,et al. Do swimming fish always grow fast? Investigating the magnitude and physiological basis of exercise-induced growth in juvenile New Zealand yellowtail kingfish,Seriola lalandi[J]. Fish Physiology and Biochemistry,2011,37(2):327-336.
[9]Huang N Y,Cheng Q Q,Gao L J,et al. Effect of water current and temperature on growth of juvenile Acipenser baeri[J]. Journal of Fisheries of China,2007,31(1):31-38.
[10]虞順年,魏小嵐,韋芳三,等. 不同運動強度對黑鯛生長、血清和肝臟抗氧化指標的影響[J]. 水生生物學報,2018,42(2):255-263.
[11]Alcaraz G,Urrutia V.Growth in response to sustained swimming in young montezumae swordtails,Xiphophorus montezumae[J]. Marine and Freshwater Behaviour and Physiology,2008,41(1):65-72.
[12]Ghanawi J,Chadi M,Saoud I P.Effect of continuous water movement on growth and body composition of juvenile rabbitfish,Siganus rivulatus[J]. Journal of the World Aquaculture Society,2010,41(5):834-839.
[13]Forster I P,Ogata H.Growth and whole-body lipid content of juvenile red sea bream reared under different conditions of exercise training and dietary lipid[J]. Fisheries Science,1996,62(3):404-409.
[14]周 玉,郭文場,楊振國,等. 魚類血液學指標研究的進展[J]. 上海水產大學學報,2001,10(2):163-165.
[15]王曉艷,王際英,馬晶晶,等. VE和L-肌肽對大菱鲆幼魚生長、抗氧化、非特異性免疫及血清生化指標的影響[J]. 水生生物學報,2017,41(1):86-94.
[16]趙書燕,林黑著,黃 忠,等. 不同蛋白質水平下添加小肽對石斑魚生長、消化酶、血清生化和抗氧化能力的影響[J]. 南方水產科學,2016,12(3):15-23.
[17]Tan N,Morimoto K,Sugiura T,et al. Effects of running training on the blood glucose and lactate in rats during rest and swimming[J]. Physiology and Behavior,1992,51(5):927-931.
[18]章羅庚.有氧運動對大鼠血糖、血脂和血液凝固功能的影響[J]. 北京體育大學學報,2009,32(7):66-68.
[19]Vijayan M M,Leatherland J F.High stocking density affects cortisol secretion and tissue distribution in brook charr,Salvelinus fontinalis[J]. The Journal of Endocrinology,1990,124(2):311-318.
[20]張 墨,李吉方,溫海深,等. 放養密度對大雜交鱘生長性能的影響及生理應答機制[J]. 海洋科學,2016,40(8):35-41.
[21]Dong J,Zhao Y Y,Yu Y H,et al. Effect of stocking density on growth performance,digestive enzyme activities,and nonspecific immune parameters of Palaemonetes sinensis[J]. Fish & Shellfish Immunology,2018,73:37-41.
[22]Ortuo J,Esteban M A,Meseguer J.Effects of short-term crowding stress on the gilthead seabream (Sparus aurata L.) innate immune response[J]. Fish and Shellfish Immunology,2001,11(2):187-197.
[23]Ming J H,Xie J,Xu P,et al. Effects of emodin and vitamin C on growth performance,biochemical parameters and two HSP70s mRNA expression of Wuchang bream (Megalobrama amblycephala Yih) under high temperature stress[J]. Fish and Shellfish Immunology,2012,32(5):651-661.
[24]Sevcikova M,Modra H,Slaninova A,et al. Metals as a cause of oxidative stress in fish:a review[J]. Veterinární Medicína,2011,56(11):537-546.
[25]Tung B T,Rodriguez-Bies E,Thanh H N,et al. Organ and tissue-dependent effect of resveratrol and exercise on antioxidant defenses of old mice[J]. Aging Clinical and Experimental Research,2015,27(6):775-783.
[26]Radak Z,Chung H Y,Goto S.Systemic adaptation to oxidative challenge induced by regular exercise[J]. Free Radical Biology & Medicine,2008,44(2):153-159.
[27]于麗娟. 運動訓練對中華倒刺鲃幼魚生長、抗氧化及免疫機能的影響[D]. 重慶:西南大學,2014.程云輝,潘藝偉,許能祥,等. 添加乳酸菌及不同含水量對多花黑麥草青貯品質的影響[J]. 江蘇農業科學,2020,48(23):160-166.
