miRNA在植物響應高溫脅迫中的研究進展
2020-02-22 04:48:32李澤卿劉彩賢邢文文亞峰
生物技術通報 2020年2期
李澤卿 劉彩賢 邢文 文亞峰
(中南林業科技大學風景園林學院,長沙 410004)
高溫是制約植物生長發育的主要環境因素之一。近年來,隨著全球年平均最低溫度和最高溫度的持續上升,全球氣候變暖和“溫室效應”現象日益明顯。氣候變暖不僅會影響植物的生長發育和植物的分布,還會嚴重降低很多經濟作物的產量和品質,直接威脅種植業的發展[1]。因此,研究高溫影響植物生長發育的機制,提高植物抵抗高溫的能力對農林業生產具有重要意義。
為了抵抗高溫脅迫的危害,植物在長期進化過程中形成了一系列應激反應機制,如熱激蛋白等分子伴侶的含量增加,能夠幫助維持細胞內蛋白的構象,減少細胞的傷害[2];或產生HSF、DREB等轉錄因子進行轉錄調控[3]。此外,也有研究表明,為了適應溫度的變化,植物進化出了復雜的表觀遺傳機制以響應環境溫度的脅迫[1]。microRNA(miRNA)調控基因的表達是植物表觀遺傳調控機制中的一種重要調控方式。microRNA(miRNA)在真核生物基因組中普遍存在,其是一類由20-24個核苷酸組成的小的非編碼RNA,通常通過序列互補降解或抑制其靶標基因轉錄后的翻譯過程,從而在轉錄水平和轉錄后水平上調控基因的表達[4]。大量研究表明,植物的miRNA在調控植物生長和發育,響應逆境脅迫方面均起重要作用[5-9]。近年來,隨著測序技術的快速發展,研究者們研究發現高溫脅迫能夠影響植物miRNA的形成,大量響應高溫脅迫的miRNA在植物中被鑒定出來,部分miRNA在高溫脅迫中的調控功能也得到證實[10-13]。
本文綜述了近年來在植物研究中發現的與高溫脅迫相關的miRNA,總結了miRNA在響應高溫脅迫過程中的主要的調控機制,及其與其他調控因子和信號通路之間可能存在內部聯系。為深入研究高溫脅迫相關miRNA的作用機制和調控通路,利用miRNA提高植物的抗熱性提供科學依據。
1 植物響應高溫脅迫的分子機制
高溫是限制植物生長發育的重要脅迫因子。分子生物學和生物技術的飛速發展為我們從分子水平認知植物抵御高溫脅迫的機制提供了重要信息和研究手段。研究表明,植物在高溫脅迫下發生一系列的應激反應,以減少高溫脅迫對于機體的傷害。在分子水平上,植物響應高溫脅迫的主要調控途徑有活性氧途徑(Reactive oxygen species,ROS)和激素調控途徑[14]、熱激轉錄因子-熱激蛋白(Heat stress transcription factor-heat shock protein、Ca離子-鈣調蛋白(Ca2+- CaM)途徑。熱信號傳遞在這3個途徑中都涵蓋3個步驟:先熱信號的感知、緊接著信號的傳遞和放大、再在分子水平上轉錄調控和功能基因的表達。通過MAPK途徑活性氧信號調控下游的轉錄因子表達,下游的MBF1c、ZAT家族和WRKY轉錄因子等轉錄因子表達能被MAPK的磷酸化激活[15]。這些轉錄因子再調控編碼具有抗氧化功能的抗氧化酶類[Ascorbate peroxidase(APX)、Catalase(CAT)等]和非酶類(還原型抗壞血酸、谷胱甘肽、類黃酮等)基因的表達,從而提高植物的抗性[16]。而參與其中的高溫脅迫響應基因可大致分為3類:第1類為轉錄調控基因及參與信號級聯導系統,如轉錄因子(WRKY、HSF、ZAT、DREB等),以及蛋白激酶、泛素化酶和磷酸化酶等;第2類為直接對蛋白及生物膜起作用的功能蛋白,如在植物體內分布最廣的是熱激蛋白HSPs(Heat shock proteins) 和 LEA(Late embryo-genesis abundant proteins)等(圖1)。在高溫脅迫下,植物體內大部分mRNA的轉錄及蛋白質的合成會受到抑制,而熱激蛋白常常作為 “分子伴侶”,參與植物受損蛋白的修復和維持植物細胞的存活[17-18];第3類為水和離子吸收和轉運相關的蛋白[14,19]。
近年來,大量研究表明,高溫脅迫除了導致植物編碼基因的表達發生改變之外,一些非編碼RNA的表達也發生了顯著改變,其中,miRNA作為重要的非編碼RNA調控手段,參與了編碼分子伴侶和轉錄因子等靶標基因的表達調控,影響了植物的抗熱性[20-21]。
2 miRNA的形成過程、作用機制及生物學功能
miRNA需要經過轉錄、加工和復合體裝配3個過程才能最終形成。植物中非編碼的MIR基因,能夠在細胞核中由轉錄聚合酶Ⅱ轉錄出初級miRNA轉錄本(Primary miRNA transcripts),它是一段能形成不完美莖環結構的序列。初級miRNA轉錄本經過3個步驟加工成為具有轉錄功能的成熟miRNA:(1)首先在鋅指蛋白SE、核糖核酸酶Ⅲ(RNaseⅢ)DCL1,以及輔酶因子HYL1的共同作用下被加工成 miRNA 的 前 體(Precursor of miRNAs);(2) 在SE、DCL1及HYL1共同作用下被加工成miRNA和miRNA* 的二聚體結構(miRNA/miRNA*duplex)[22];(3)其中被甲基化的一條miRNA*的二聚體會被導出細胞核進入細胞質,然后與Agonaute蛋白(AGO)結合裝配成RISC,這時的sRNA具有了轉錄后沉默功能,是成熟的miRNA。而一般核酸酶會降解另一條被稱為miRNA*的二聚體。有研究者發現,其中一條在二聚體的5'端并配對不緊密的miRNA與AGO蛋白結合并行使功能[23]。
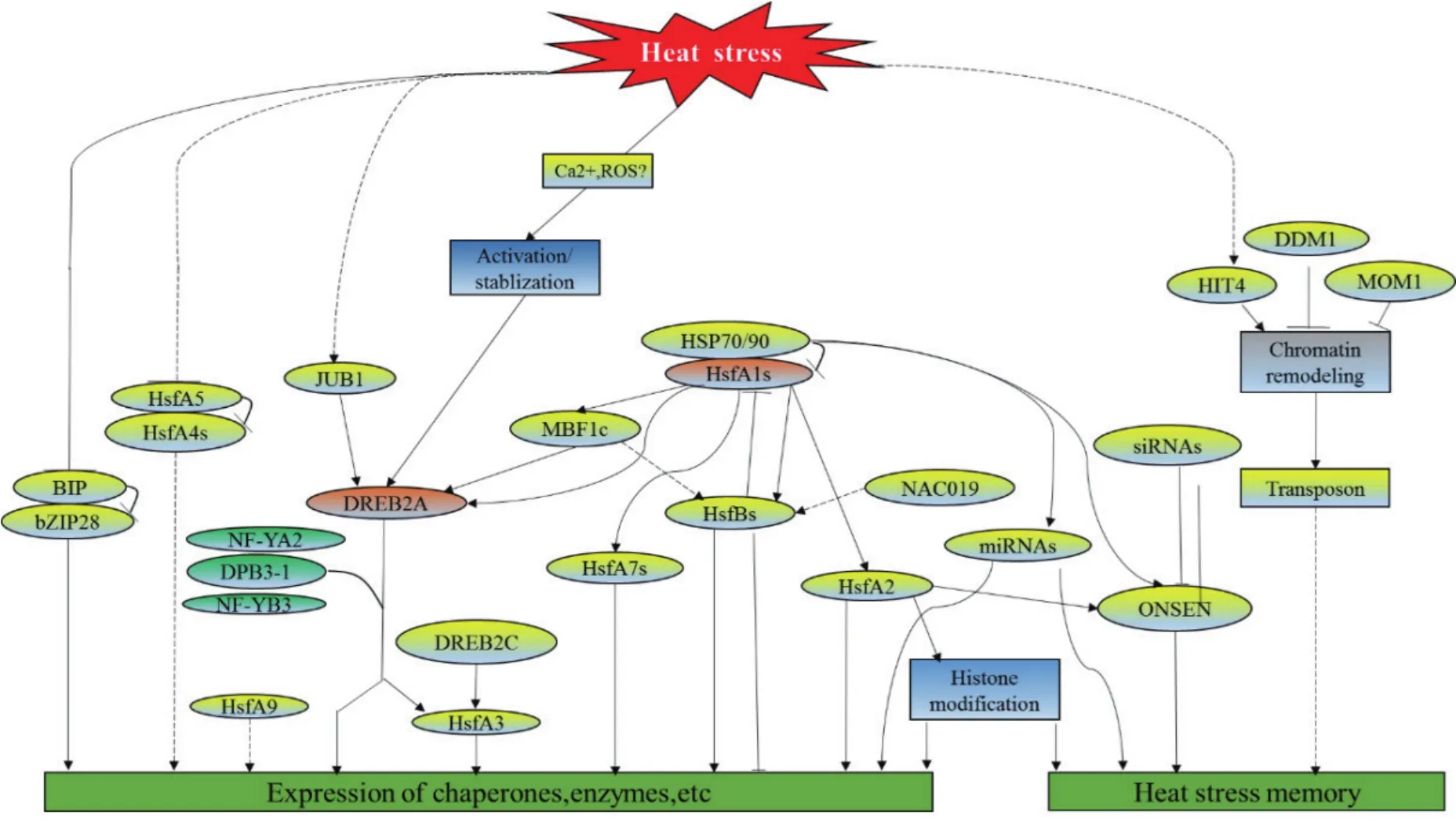
圖1 高溫脅迫下轉錄因子調控網絡(改自Ohama 等[3])
miRNA通過堿基互補配對原則識別靶基因[5]。根據其與靶基因的互補程度不同,miRNA通過對靶mRNA切割和翻譯抑制2種轉錄調控機制,指導RISC下調基因表達:在靶基因和miRNA匹配性廣泛或者匹配度較高的時候,mRNA會被RISC剪切;而當靶基因和miRNA的匹配度較低且低到不能夠讓RISC剪切靶基因的時候,靶基因會被RISC通過翻譯抑制而沉默[23]。因為植物miRNA和靶基因的匹配屬于廣泛性匹配,所以miRNA主要是通過RISC選擇剪切mRNA來抑制靶基因的表達[24]。不過還是有一小部分植物的靶基因和miRNA的匹配度較低,則miRNA調控RISC通過翻譯抑制來沉默靶基因[25-26]。
近年來,隨著高通量測序技術的發展,越來越多的miRNA從植物中被鑒定。在最新的第22版本miRBase里,一共有326個編碼miRNA的位點存在于擬南芥基因組,有588個編碼miRNA的位點存在于水稻基因組,有697個編碼miRNA的位點存在于楊樹基因組(http://www.mirbase.org/)。隨著對miRNA研究得不斷深入,研究者們發現miRNA在植物的生長和發育中均起著調控作用[5-6],其作為調控因子,廣泛地參與到植物生長發育的不同階段(表1)。此外,miRNA也參與到了植物響應生物及非生物逆境的調控網絡中。近年來,越來越多參與響應各種生物及非生物逆境的miRNA在模式植物被鑒定出來(表1)。已有大量研究表明,miRNA在植物響應高溫脅迫中也充當重要角色[21,27-44]。
3 miRNA在植物響應高溫脅迫中的研究
3.1 miRNA在植物響應高溫脅迫中的表達
近年來,隨著測序技術的快速發展與成熟,通過small RNA深度測序,從不同植物中鑒定出大量響應高溫脅迫的保守或者非保守 miRNA[12-13,45]。研究表明,在高溫脅迫下,植物miRNA的表達呈現出或上調或下調的表達方式,在部分植物中上調與下調的miRNA的數量差異并不顯著:在水稻中鑒定出47個響應高溫脅迫的miRNA,其中26個miRNA表達下調,21個miRNA表達上調[11];在番茄中,處于適當升溫的環境下,分別有39/38個已知的miRNA出現了顯著性的上調/下調,而在劇烈的升溫下,則分別有62/57個已知的miRNA出現了顯著性的上調/下調[58]。而部分植物中則大部分表現為下調表達:在蘿卜中,26個已知的和19個新的miRNA在高溫脅迫下出現了顯著性差異表達,且大部分表現為下調表達,只有少數上調表達[59]。在中國白楊中,15個高溫響應的miRNA家族被鑒定出來,且大部分miRNA的表達量下調[27]。
在不同植物中響應高溫脅迫的miRNA不盡相同,目前,已發現在大部分植物中均響應高溫脅 迫 的 miRNA 包 括 miR472、miR408、miR398、miR395、miR171、miR169、miR168、miR167、miR166、miR160、miR159 和 miR156 等[28-29]。 相同的miRNA在不同植物響應高溫脅迫下的的表達模式也不完全相同(表2)。miR159在木薯和水稻中在受高溫脅迫后下調表達[11,30];而在擬南芥、小麥和大白菜中則表現為上調表達[33,35,41]。miR160 則在木薯和小麥中下調表達,而在水稻、擬南芥和棉花中上調表達[11,21,28,48,60]。在高溫脅迫下,miR398則在水稻、小麥和擬南芥中上調表達,而在大白菜中下調表達[33,35,41]。
還有研究發現,同一家族的miRNA在不同植物中對高溫脅迫的響應模式也不同。在小麥、擬南芥和大白菜中,高溫脅迫下,miR156家族的miR156a、miR156h和miR156g上調表達;而在樺木、木薯和水稻中,miR156a和miR156d則受高溫抑制表達[11,30,32-33,35,61]。
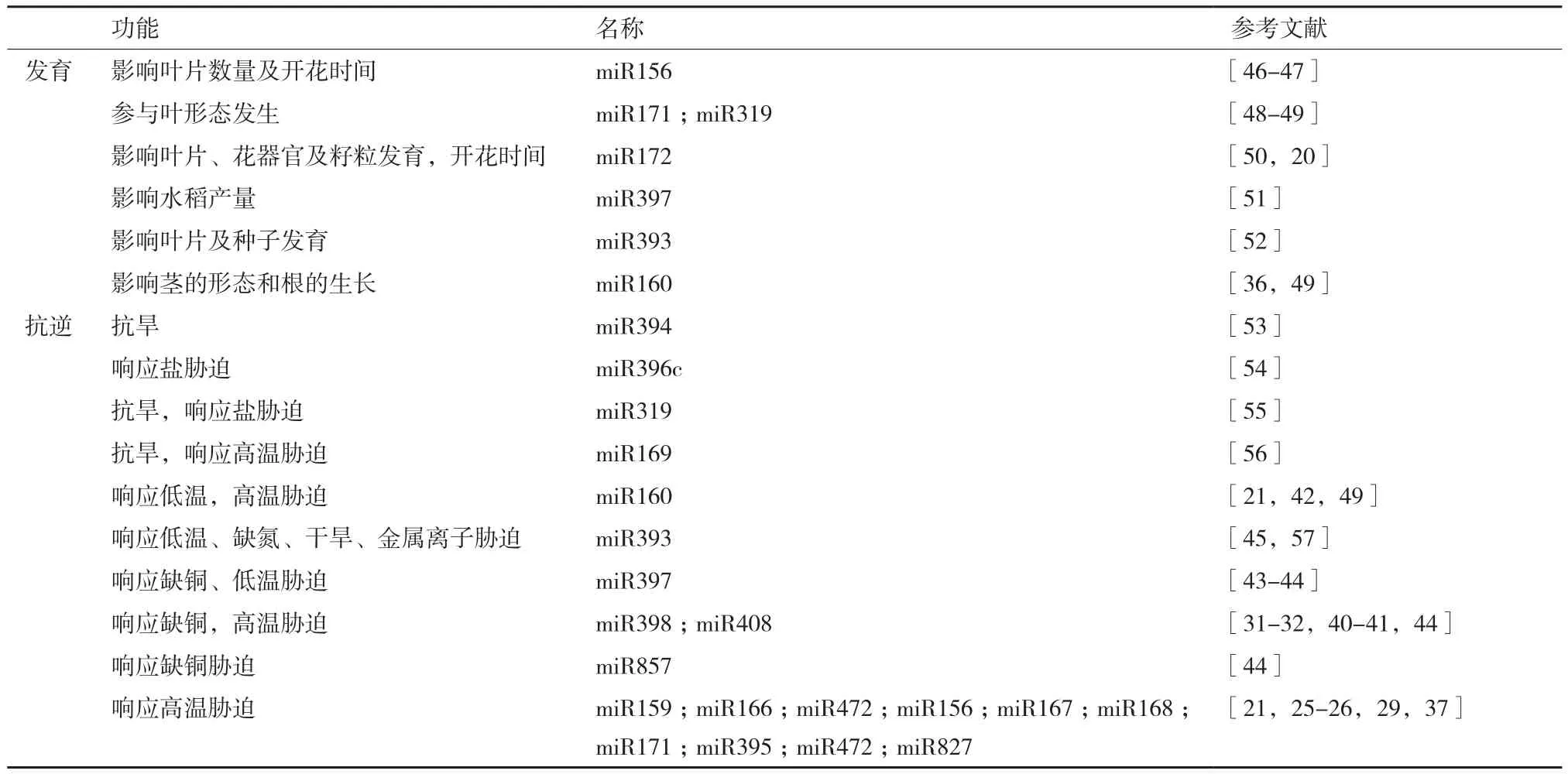
表1 miRNA的生物學功能
3.2 miRNA在植物高溫脅迫響應中的調控機制
miRNA主要是通過抑制靶基因的表達參與到植物的各種代謝和信號途徑當中。隨著研究得不斷深入,明確了部分miRNA的靶向基因,并對其在高溫脅迫下的作用機制有了一定的了解。同一miRNA可能存在多個靶向基因,且不同植物受到高溫脅迫時,作用的靶向基因不完全一致。轉錄因子是miRNA作用的重要靶向基因之一。目前發現的植物在高溫脅迫下受到miRNA調控較多的有ARF、MYB、WRKY和SPL等家族的轉錄因子(表2)。miR160的靶向基因為ARF類轉錄因子:miR160在擬南芥中抑制其靶基因ARF17,ARF16和ARF10的表達,并通過調控HSP基因的表達從而提高擬南芥的抗熱性[21];而在玉米中,miR160參與生長素信號途徑,通過抑制其靶基因ARF17和ARF10的表達,從而導致高溫脅迫下玉米雄性不育[62]。miR156在擬南芥中作用的靶向轉錄因子為SPL家族轉錄因子,miR156通過抑制SPL的表達增強了擬南芥對高溫的記憶從而調節抗熱性[33]。而在小麥中,高溫脅迫下miR156的靶向基因為TaGAMYB1和TaGAMYB2[63]。此外,在耐熱性較好的向日葵中還發現miR396通過調控WRKY家族轉錄因子HaWRKY6的表達,從而幫助向日葵不受到高溫脅迫的傷害[60]。除了轉錄因子以外,在小麥中還發現了miR156、miR159、miR160、miR167、miR319、miR395和miR398等眾多miRNA家族參與了HSP17、HSP70和superoxide dismutase等高溫脅迫相關基因的表達調控[35]。
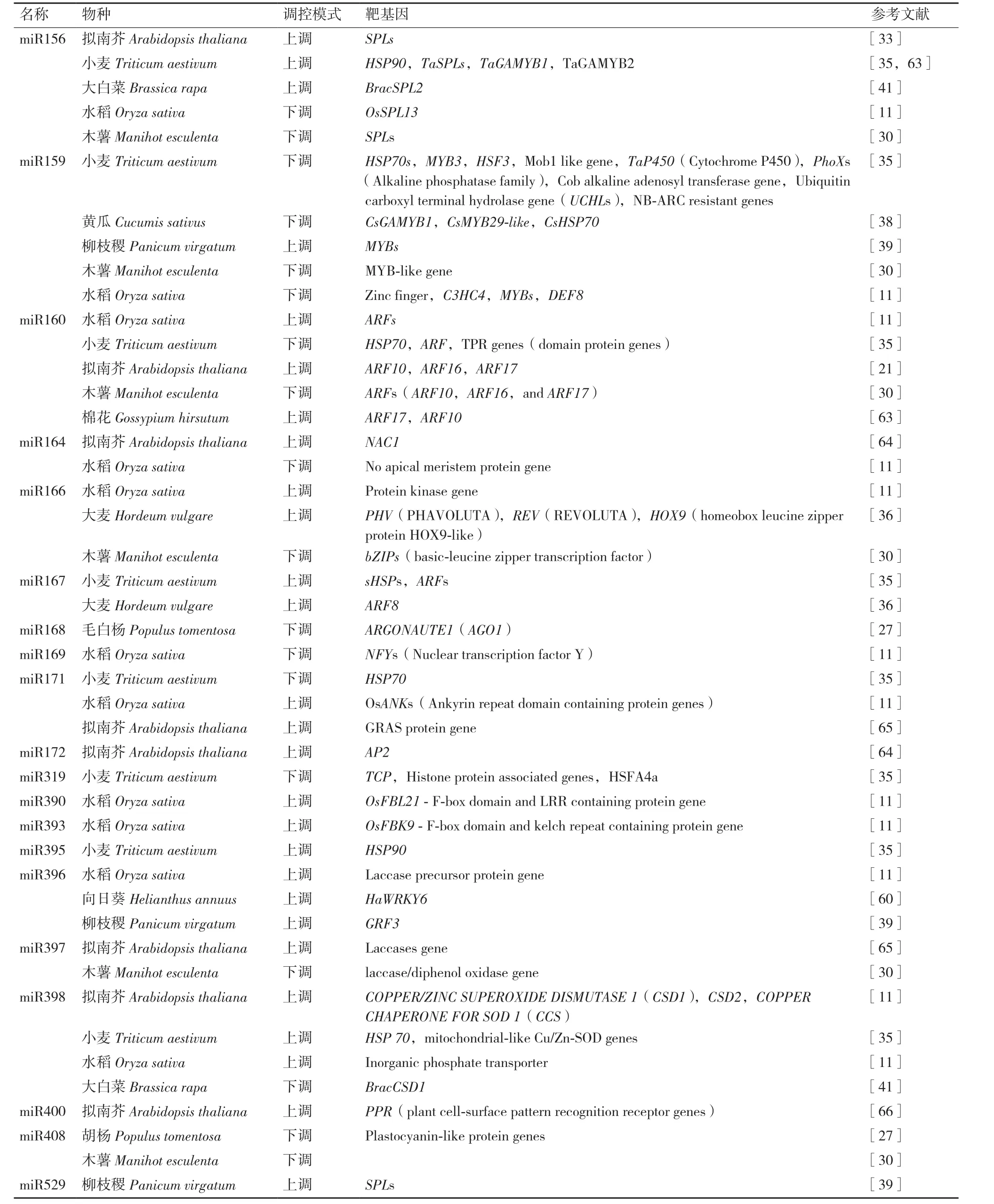
表2 植物中響應高溫脅迫的miRNA
大量研究還表明,miRNA通過調控激素響應途徑參與到植物響應高溫脅迫。miR160、miR167、miR390和miR393在高溫脅迫下均參與到了生長素代謝途徑中[35-37]。miR159在高溫脅迫下能夠調控GA3代謝途徑[35,38]。而 miR319 在高溫脅迫下則參與調控茉莉酸代謝途徑[35,39]。
在這些響應高溫脅迫的miRNA中,miR398和miR156在植物響應高溫脅迫中的調控途徑研究較為深入。在擬南芥中miR398的靶標為控制ROS的產生的COPPER/ZINC SUPEROXIDE DISMUTASE 1(CSD1)、CSD2和 COPPER CHAPERONE FOR SOD 1(CCS)基因[40]。miR398超量表達抑制了這些活性氧清除酶的表達,導致ROS的積累,改變細胞內氧化還原狀態,從而誘導HSPs及HSFs的表達,最終提高了擬南芥對高溫的抗性。還發現HSFA7b和HSFA1b能結合在miR398的啟動子區域,從而參與miR398的熱誘導過程[40]。因此在擬南芥中,miR398與其靶標CSD1、CCS以及CSD2,HSFs共同組成了一條耐熱回路(圖3)。除了擬南芥以外,在大白菜和毛白楊中同樣也發現了類似的耐熱回路[28,41]。與miR398不同,在擬南芥中,miR156的靶向基因為SQUAMOSAPROMOTER BINDING-LIKE(SPL)transcription factors。該家族轉錄因子的下調表達能夠促進熱擊響應基因HsfA2和HSPs的穩定持續表達,從而提高植物的抗熱性(圖3)。研究還發現miR156在高溫脅迫下特異表達。因此,miR156和SPLs組成的調控模塊被認為是植物形成高溫脅迫記憶的關鍵元件,對植物的獲得性抗熱有促進作用[33]。
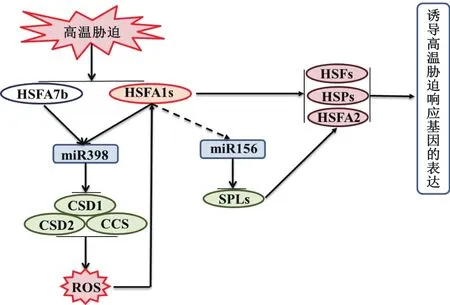
圖3 擬南芥中miR398和miR156與其靶基因參與的耐熱性調控途徑
4 結語
植物逆境脅迫廣泛存在于植物的生長發育過程中,而隨著溫室效應的愈加嚴重,高溫脅迫對植物的生長和發育影響愈加深遠。近年來,對植物中響應高溫脅迫的miRNAs研究已經取得一定進展:隨著測序技術的不斷發展,諸多miRNA及其靶基因被發現和鑒定;已有研究明確了miRNA作為重要的非編碼RNA通過轉錄后調控廣泛參與植物高溫脅迫響應,其調控的靶基因大多編碼轉錄因子或酶類。然而,目前,miRNA參與植物的高溫脅迫的報道主要集中于模式植物中,對在其他觀賞植物和經濟植物中的研究報道較少。且大多miRNAs及其靶基因的功能僅停留在生物信息學預測與分析上,并沒有得到試驗證明。miRNA及其靶基因是如何參與到植物響應高溫脅迫的分子調控網絡途徑還有待進一步研究。但不可否認的是,以miRNA差異表達以及對靶基因的調控角度探究植物在高溫脅迫下的調控機制為植物抗高溫脅迫提供了新的研究思路,對挖掘植物抗高溫機制以及培育抗高溫新材料具有十分重要的意義。在后續的研究中,miRNA及其靶基因在高溫脅迫中的功能驗證及其參與植物響應高溫脅迫分子調控網絡的作用機制將成為植物抗高溫育種的研究重點。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
