氧化應激在魚藤素致SH-SY5Y細胞神經毒性中的作用
2020-01-09 07:41:42方瓊彤呂滿霞陳彥潔柳傳毅吳新榮
中國藥理學通報 2020年1期
方瓊彤,呂滿霞,陳彥潔,柳傳毅,吳新榮
(1. 南方醫科大學藥學院,廣東 廣州 510515;2. 中國人民解放軍南部戰區總醫院藥學部,廣東 廣州 510010)
魚藤素是一種天然黃酮類化合物,在腫瘤發生、轉移、凋亡和血管生成的綜合信號通路中,在腫瘤細胞凋亡和惡性轉移過程中發揮著關鍵的作用。魚藤素對肺癌、乳腺癌、肝癌、淋巴癌及骨髓瘤等多種腫瘤具有化學防治作用[1-4],揭示了魚藤素對多種腫瘤有潛在的預防和治療作用。但是有研究表明在高劑量的魚藤素濃度下可能會產生毒副作用,能夠減弱陽性神經元酪氨酸羥化酶的活性,從而引發帕金森樣綜合癥[5],這阻礙魚藤素成為潛在的抗腫瘤藥物。
氧化應激對正常細胞的生長起調控作用,在腫瘤細胞代謝中也能產生重大的影響[6]。氧化應激是由自由基在體內產生的一種負面作用,并被認為是導致衰老和疾病的重要因素[7-8]。由于活性氧的反應性,高濃度的活性氧(reactive oxygen species,ROS)可導致細胞死亡或氧化應激,即破壞促氧化和抗氧化水平之間的平衡,并且ROS能夠氧化各種細胞成分,導致細胞氧化破壞[9]。神經元細胞膜中的不飽和脂肪酸含量高,耗氧量高,抗氧化防御能力弱,因此,氧化應激極易對神經系統造成損傷,造成神經細胞受損和神經元的丟失,從而演變為神經退變性疾病[10-11]。
MAPK/ERK信號通路調控多種細胞活動,包括增殖、分化、存活和死亡。MAPK/ERK信號通路的失調與許多人類疾病的發展密切相關,其中包括阿爾采末病(Alzheimer’s disease,AD)、帕金森病(Parkinson's disease,PD)和各種腫瘤[12]。有相關研究表明MAPK/ERK信號通路對于神經細胞功能的調節起到十分重要的作用[13]。
本研究通過測定氧化應激相關指標和MAPK/ERK信號通路相關蛋白表達,探究魚藤素致神經毒性的潛在相關機制。
1 材料
1.1 細胞株人神經瘤細胞株SH-SY5Y,由人民解放軍南部戰區總醫院醫學實驗科提供
1.2 試劑魚藤素(美國sigma公司,批號:D0817,HPLC 級)、二甲基亞砜(美國sigma公司,批號:RNBD6895);活性氧ROS檢測試劑盒(批號:S0033),LDH乳酸脫氫酶測試試劑盒(批號:A020-1),總SOD活性檢測試劑盒(批號:S0101),脂質氧化(MDA)檢測試劑盒(批號:S0131),谷胱甘肽過氧化物酶檢測試劑盒(批號:S0056),Western blot及IP細胞裂解液(批號:P0013),100 mmol·L-1PMSF(批號:ST506),BCA 蛋白定量試劑盒(批號:P0012),SDS-PAGE蛋白上樣緩沖液(5×)(弗德生物公司,批號:FD002),SDS-PAGE凝膠快速配制試劑盒(批號:P0012AC),均購自碧云天公司;蛋白Maker(美國Thermo公司,批號:26616),山羊抗兔IgG H&L (HRP) (天津三箭,批號:LK2001);p-ERK1/2 Rabbit mAb(批號:4370),ERK1/2 Rabbit mAb(批號:4695)均購自Cell signaling technology;Anti-MEK1 + MEK2抗體(批號:ab178876),Anti-MEK1 + MEK2 (phospho S218 + S222)(批號:ab194754),Anti-CREB抗體(批號:ab32515),均購自abcam;EGF Antibody(批號:DF2225),GAPDH Antibody(批號:AF7021),均購自affinity。
1.3 儀器3111 水套式CO2細胞培養箱(美國Thermo公司),G41054-F000型臺式離心機(KUBOTA公司),IX7型倒置熒光顯微鏡(日本OLYMPUS公司),HWS28型恒溫水浴鍋(上海精宏實驗設備有限公司),Multiscan-GO型全波長酶標儀,RRESCO 17型低溫高速離心機(美國Thermo公司),BD LSRFortessa流式細胞儀 (美國 BD 公司),ACY-1002-U型超純水機(美國 Aquapro 公司),PowerPac 電泳儀、Sub-Cell 電泳槽、Tans-Blot Turbo Transfer System轉膜儀(美國伯樂 Bio-Rad),MiniChemi Ⅲ化學發光成像儀(北京賽智創業科技有限公司)。
2 方法
2.1 魚藤素的配制100 mmol·L-1魚藤素母液配制:將50 mg魚藤素完全溶解于1.27 mL的DMSO溶液中,并于-20 ℃低溫避光保存,實驗組中不同濃度的魚藤素用新鮮培養基稀釋,現配現用。
2.2 細胞培養從液氮罐中取出神經母細胞瘤(SH-SY5Y)細胞,迅速37 ℃水浴復蘇,并在超凈工作臺將細胞轉移至離心管中,加入適量含10%胎牛血清的高糖DMEM培養基,以1 000 r·min-1轉速離心5 min ,去掉上層舊液后加入新鮮培養基,于37 ℃、5% CO2恒溫培養箱培養,取對數生長期細胞進行后續實驗。
2.3 CCK-8法檢測細胞增殖毒性取對數生長細胞,以5×107·L-1的密度接種于96孔板中,每孔接種細胞液100 μL,每組5個復孔,放入37 ℃、5% CO2恒溫培養箱培養,使細胞貼壁生長。次日去除舊培養基,以濃度為1.56 μmol·L-1,3.13 μmol·L-1,6.25 μmol·L-1,12.5 μmol·L-1,25 μmol·L-1,50 μmol·L-1,100 μmol·L-1的魚藤素作用于SH-SY5Y細胞 24 h、48 h、72 h 后,將培養基與CCK-8試劑按9 ∶1比例配制的混合液以每孔100 μL加入,在培養箱中孵育2 h,在450 nm波長處,酶標儀測定吸光度OD值。細胞存活率/%=[(OD實驗組-OD空白組)/(OD對照組-OD空白組)] ×100%。實驗平行重復3次,后續計算得到其細胞存活率。
2.4 細胞比色法測定LDH、MDA、SOD、GPx活力根據細胞中乳酸脫氫酶(lactate dehydrogenase,LDH)催化乳酸生成丙酮酸二硝基苯肼化合物,產物與顯色劑反應生成的化合物在堿性溶液中呈棕紅色,通過比色法可測得細胞上清液中LDH的含量。根據細胞中的LDH漏出量,可判斷細胞的受損程度。取對數生長期細胞,以1×108·L-1的密度,按每孔2 mL進行接種,均勻分布。在恒溫培養箱中培養24 h,使細胞貼壁。根據CCK-8法測定的結果設置低、中、高濃度實驗組如下:A組:對照組;B組:12.5 μmol·L-1魚藤素;C組:25 μmol·L-1魚藤素;D組:50 μmol·L-1魚藤素。藥物處理細胞后,取細胞培養上清液,按照LDH試劑盒操作說明進行檢測和計算。

2.5 流式細胞術法檢測細胞ROS水平取對數生長期細胞,以1×108·L-1的密度,按每孔2 mL進行接種,均勻分布。在恒溫培養箱中培養24 h,使細胞貼壁。實驗分組同“2.4”,藥物作用24 h后收集細胞,1 000 r·min-1轉速下離心5 min,去除上清液。用無血清的培養基按1 000 ∶1的比例將10 mmol·L-1的DCFH-DA母液,每個樣品用1 mL稀釋液懸浮細胞,在37 ℃避光敷育30 min。期間每隔5 min振蕩1次,離心后,PBS洗滌并重懸細胞。用流式細胞儀檢測各組細胞ROS水平。
2.6 Western blot 檢測細胞中EGF、ERK、CREB蛋白的表達取對數生長期的SH-SY5Y細胞,每孔均勻接種2 mL密度為1×108·L-1的細胞,于培養箱培養24 h貼壁后,實驗分組同“2.4”,藥物處理24 h后,除去舊培養基,加入2 mL PBS緩沖液洗滌2次,加入100 μL配置好的細胞裂解液(每1 mL的細胞裂解液中含10 μL PMSF,現配現用),在冰上裂解30 min,期間間隔5 min吹打細胞,細胞刮收集蛋白裂解液,并置于提前預冷至4 ℃的離心機中,12 000 r·min-1轉速下離心20 min。離心完畢,將上清液吸出,通過BCA法測定蛋白濃度,按體積4 ∶1加入5×loading buffer,于沸水中煮沸5 min,使蛋白充分變性,保存于-20 ℃冰箱中。采用12% SDS-PAGE膠跑電泳,用PVDF膜進行轉膜,用5%的脫脂奶粉配置的TBST封閉液室溫封閉1 h,TBST緩沖液洗膜3次,每次10 min,然后加稀釋1 000倍的一抗4 ℃孵育過夜,一抗孵育完畢,回收孵育盒中的一抗,加入1×TBST緩沖液洗膜3次,每次10 min。加入相應稀釋5 000倍的二抗孵育1 h,用1×TBST緩沖液洗膜3次,每次10 min,最后ECL顯像液顯影,得出蛋白條帶灰度后,使用Image Jx軟件分析蛋白條帶的灰度值,以GAPDH為內參,以目的條帶光密度值和GAPDH光密度值的比值即為目的蛋白的相對表達水平。
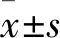
3 結果
3.1 魚藤素對SH-SY5Y細胞增殖的影響濃度為1.56、3.13、6.25、12.5、25、50、100 μmol·L-1的魚藤素作用SH-SY5Y細胞 24 h、48 h、72 h 后,測得其細胞存活率如下圖如Fig 1 所示。隨著藥物作用濃度的增加和作用時間的延長,SH-SY5Y細胞的存活率均下降(P<0.05)。魚藤素作用于SH-SY5Y細胞24 h、48 h、72 h的IC50值分別為28.25±1.67、19.36±1.44、11.50±0.89 μmol·L-1。結果表明,魚藤素對SH-SY5Y細胞的增殖有抑制作用并呈濃度和時間依賴性。

Fig 1 Effects of different concentrations of deguelin on inhibitory rate of SH-SY5Y cells treated for 24, 48 and 72 h n=3)
3.2 魚藤素對SH-SY5Y細胞的LDH漏出量的影響細胞上清液中LDH水平反映了細胞的受損程度。0、12.5、25、50 μmol·L-1魚藤素作用于SH-SY5Y細胞24 h后,測得SH-SY5Y細胞的LDH漏出量,結果如Fig 2所示。結果表明,高于25 μmol·L-1的魚藤素作用SH-SY5Y細胞后,LDH漏出量明顯增加,并呈現濃度依賴性。

Fig 2 The LDH leakage (% of control) of SH-SY5Y cells treated with different concentrations of deguelin for 24 h n=3)
3.3 魚藤素影響SH-SY5Y細胞的ROS水平0、12.5、25、50 μmol·L-1魚藤素作用于SH-SY5Y細胞24 h后,測得SH-SY5Y細胞的ROS水平,結果如Fig 3和Tab 1所示。與對照組相比,12.5、25、50μmol·L-1魚藤素處理后的細胞內的ROS水平明顯增加,對照組中的ROS水平為1102.4±105.6,而隨著魚藤素給藥濃度的增加,ROS水平增加,12.5 μmol·L-1、25 μmol·L-1、50 μmol·L-1魚藤素作用后測得ROS分別為1891.6±92.71、2828.3±76.49、4605.0±141.6,結果表示ROS水平的變化呈濃度依賴性,同時說明魚藤素對SH-SY5Y細胞造成的損傷可能與ROS產生有關。

Fig 3 Determination of ROS of SH-SY5Y cells after deguelin treatment for 24 h n=3)

Tab 1 ROS intensity of SH-SY5Y cells after deguelin treatment for 24 h
3.4 魚藤素對SH-SY5Y細胞的SOD和Gpx活力和MDA含量的影響0、12.5、25、50 μmol·L-1魚藤素作用于SH-SY5Y細胞24 h后,測定細胞中SOD和Gpx的活力以及MDA含量,結果如Fig 4 所示,與對照組相比,隨著魚藤素作用濃度的增加,SH-SY5Y細胞內SOD和GPx活力出現逐步降低的趨勢;MDA含量明顯升高。魚藤素對SH-SY5Y細胞的損傷伴隨著氧化應激相關指標的變化,說明氧化應激與其神經毒性密切相關。
3.5 魚藤素對SH-SY5Y細胞中MAPK/ERK信號通路的影響0、12.5、25、50 μmol·L-1濃度的魚藤素作用于SH-SY5Y細胞24 h后測定細胞內MEK、EGF、RAS、CREB蛋白表達水平,結果如Fig 5 所示。與對照組相比,各個濃度的魚藤素組能夠下調P-MEK、MEK和CREB蛋白蛋白的表達(P<0.05)。25.0 μmol·L-1及其以上濃度的魚藤素能夠下調EGF和RAS蛋白的表達(P<0.01)。

Fig 4 SOD(A) , Gpx(B) and MDA(C) activities of SH-SY5Y cells treated with different concentrations of

Fig 5 Expression levels of p-MEK, MEK(A) and EGF, RAS, CREB(B) in SH-SY5Y cells treated with different concentrations of deguelin for 24 h n=3)
4 討論
許多研究表明,魚藤素具有潛在的抗腫瘤活性,但是給予大鼠治療劑量的魚藤素后,大鼠出現了帕金森樣綜合癥[5]。中樞神經系統退行性疾病的發生與氧化應激密切相關。氧化應激被認為是衰老和各種神經障礙的關鍵因子,酶催化的各種氧化還原反應在氧化磷酸化過程中發生[10]。當機體發生氧化應激時,其體內脂質氧化水平會提高,同時清除自由基能力會下降,促進了神經細胞的凋亡[14]。SOD和Gpx能夠清除活細胞內過氧化物,ROS、MDA是與脂質氧化水平密切相關的標志物,測定細胞中抗氧化酶的活力和細胞脂質氧化水平,可以探究細胞的損傷情況。本研究采用的神經母細胞瘤SH-SY5Y細胞系具有合成多巴胺和去甲腎上腺素能力,被廣泛用于神經系統疾病發病機制的研究[15]。實驗結果發現,魚藤素干預后,細胞內氧化應激水平提升,并隨著魚藤素濃度的增加愈發明顯,說明魚藤素使SH-SY5Y細胞發生氧化應激,致使細胞抵御外界損傷能力減弱。
采用蛋白免疫印跡實驗探究MAPK/ERK通路中的EGF、RAS、CREB、p-MEK和MEK蛋白表達水平,發現隨著魚藤素濃度的增加,對蛋白表達均有不同程度的抑制。結果說明,MAPK/ERK通路可能在魚藤素致SH-SY5Y細胞毒性中可能起關鍵作用。后續需要通過基因沉默技術與靶向抑制劑進行深入的探究其是否為魚藤素產生神經毒性的靶向信號通路。
綜上所述,魚藤素致SH-SY5Y細胞神經毒性的機制可能是降低細胞內抗氧化酶SOD和Gpx的活力,增加丙二醛MDA和活性氧ROS的生成,導致細胞不飽和脂肪酸過氧化。其中可能通過下調MAPK/ERK通路的相關蛋白表達使細胞活性受損,這為進一步研究魚藤素的結構改造與降低魚藤素毒性提供依據。
(致謝:本實驗在南部戰區總醫院醫學實驗科完成,特別感謝醫學實驗科各實驗室老師的指導和幫助)
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
西南軍醫(2016年6期)2016-01-23 02:21:19
學苑創造·A版(2015年11期)2016-01-14 09:03:27
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
西南軍醫(2015年2期)2015-01-22 09:09:37
食品科學(2013年15期)2013-03-11 18:25:48
