羊MHC基因與疾病相關性研究進展
2020-01-08 02:52:26宋曉越劉錦旺朱海鯨李隴平
中國獸醫雜志 2019年8期
關鍵詞:研究
宋曉越,屈 雷,史 雷,劉錦旺,朱海鯨,李隴平
(榆林學院 陜西省陜北絨山羊工程技術研究中心,陜西 榆林 719000)
主要組織相容性復合體(Major histocompatibility complex,MHC)基因是廣泛存在于脊椎動物染色體上的一段緊密連鎖且高度多態的多基因家族。MHC基因可編碼種類極為豐富的產物,主要用于呈遞抗原從而引起機體的免疫應答。眾多研究表明,MHC基因與抗病性及繁殖成功率密切相關[1]。
羊隸屬于偶蹄目羊亞科,除肉用和毛用外,羊奶、羊血、羊骨、羊肝和羊膽等還具較高藥用價值,可治療多種疾病。近年來,我國許多農戶依靠養羊而大發“羊財”。但在養殖過程中存在著眾多的疫病危害,例如結核病和布魯菌病,都屬于人獸共患病,嚴重危害人畜的健康,對養羊戶和社會造成巨大的經濟損失。目前,對疫病的防治主要采用消毒、疫苗防控和藥物治療等方式,雖起到一些作用,但每年各地仍有一定規模的疫情暴發,威脅人畜健康,對養殖業也造成較大沖擊。如何從根本上減少疫病感染,提高羊的抗病性能已成為一項急待解決的重要課題。目前,國內外研究已發現MHC基因與多種動物的抗病性緊密相關[2],從而使MHC基因成為了家畜抗病育種的新希望。因此,本文總結了近年國內外對羊MHC基因與抗病相關性的研究進展,旨在為羊等家畜的疫病預防提供新的思路,并展望基于MHC基因提高羊等家畜免疫力和抗病性能的前景。
1 MHC基因結構與特點
依據基因的功能,MHC基因可分為I類、II類和III類3個亞家族。每一亞家族中又包含著若干個基因座位,研究表明,這些基因座位中很多都具有高度的多態性。I類與II類MHC分子能夠分別呈遞內源性和外源性的抗原,參與調節機體的免疫反應[3];III類分子主要是一些補體成分。
綿羊MHC稱為綿羊白細胞抗原(Ovine leucocyte antigen,OLA),位于20號染色體上;山羊的MHC簡稱為GOLA。研究表明,羊的MHC基因具有豐富的多態性,特別是在DQ和DR區。而MHC基因的多態性水平恰恰反映了物種對疾病的抵御能力和種群的生存能力。這是由于不同MHC分子的抗原結合位點(Antigen binding site,ABS)構成不同,因此能夠呈遞的抗原種類也有差異,從而可抵御種類繁多的病原體[4]。
2 羊MHC基因與疾病的相關性
目前,已有大量研究表明,羊MHC基因確實與數十種疾病的抗性或易感性相關。其中大部分是由細菌、病毒等微生物引發的傳染病和寄生蟲病(見表1)。
2.1 由細菌、病毒等微生物引發的傳染病 在綿羊中,Hulme等[5]發現,MHC基因的多態性和蛇形毛圓線蟲病相關。Litchfield等[6]研究表明,MHC基因與綿羊對腐蹄病的抵御有關系。隨后有報道發現,OLA-I、DRB1、DY和C4基因與綿羊胃腸道線蟲病相關;OLA-DRB1的多態水平與綿羊對慢性進行性肺炎和肺腺癌(OPA)的抵御能力有關[7-8]。但也有報道表明,MHC基因與疾病無顯著的相關性[9]。我國眾多研究者也在積極開展綿羊MHC基因與疾病抗性的研究,并獲得了豐碩的成果。劉秀等[10]發現,雖然藏綿羊MHC等位基因的種類與國外品種存在差異,但仍可推斷出DQA2*F和DQA2*L等位基因具有較強的抗腐蹄病潛力。類似的結果在Chios羊的DQA2中也得到了證實[11]。陳月娥等[12]發現,中國美利奴羊DRB1基因SNP 109C > T位點與布魯菌病有關,Hap8和Hap9單倍型在病例組中的頻率均顯著高于正常組(P<0.05),推測這2種單倍型和布魯菌病的易感性有關。隨后DQB2基因也被發現與布魯菌病易感性呈顯著相關[13]。還有報道表明,巴什拜羊和盤羊的DRB3與支原體肺炎有關[14];甘肅高山細毛羊、小尾寒羊的DQB1和DRB1基因與乳房炎有關[15-16]。
在山羊中,Ruff和Lazary首次證明了導致關節炎的CAE病毒(Caprine arthritis-encephalitis virus)受MHC系統的影響[17-18]。之后相繼發現DRB位點與羊副結核病的抗性及易感性相關[19];DYA位點還與由立克次氏體引起的心水病的易感性有關[20];高山野山羊的MHC雜合性與對角膜結膜炎的抗性顯著相關[21]等。
2.2 寄生蟲病 研究發現,多個品種的綿羊DRB基因都與對線蟲的抗性和易感性有關;哈薩克綿羊的DRB1基因、多浪羊和中國美利奴羊DQB1、DRB1基因的若干個位點都分別與棘球蚴病(包蟲病)的抗性和易感性相關[22-26]。
在山羊中,發現GOLA-DYA基因與線蟲病的易感性有關[20];Corley等[27]發現肉用山羊的DRB1*1101等位基因表達量在被線蟲感染的山羊組中顯著高于抗性山羊組(P<0.05),說明是線蟲病的易感性等位基因。
3 MHC基因的抗病機制
研究表明,I類和II類MHC基因都具有很高的多態性,特別是在抗原結合位點。MHC基因正是借助于表達出的高度變異的抗原結合區來抵御種類繁多的病原體。MHC基因的抗病機制主要表現在抗原結合區和特定的抗原肽能否結合以及結合能力的強弱。而MHC基因的多態性則直接影響著抗原結合區的蛋白質組成與構象,從而形成與特定的抗原肽在結合能力上的差異。可以說,MHC基因的高度多態性是物種抵御病原體的一個良好表征,而且對于適應性進化和免疫的研究都具有意義。邢鳳等[28]發現,波爾山羊、萊蕪黑山羊和魯波山羊DQA2及DQB1基因都具有較高的多態性,并且揭示了DQA2和DQB1基因都與血液免疫指標存在著一定的相關性。惠文巧等[29]對不同MHC基因型的哈薩克綿羊人工感染細粒棘球病,發現抗性組表現出以抗體IgE和IgM以及Th1細胞因子為優勢的分泌,這提示著具有抗性MHC基因型的綿羊可能受其高水平細胞因子和抗體的作用而表現出強的抗病能力。蔣松等[30]利用哈薩克綿羊MHC基因的DRB1-MvaⅠbc-SacⅡab-Hin1Ⅰab單倍型對細粒棘球蚴病的抗性來區分不同基因型的哈薩克綿羊,分別將抗病羊和非抗病羊人工感染細粒棘球蚴病一定時間后取小腸組織做Solexa深度測序,發現miRNA的差異表達主要涉及表皮細胞細菌入侵、代謝途徑和腫瘤通路等,說明上述途徑中有可能具有和宿主抗病性相關的調控機制,而特定的MHC基因型與免疫相關基因的表達有聯系。類似的方法還推測在哈薩克綿羊中高表達的PPA1基因可能和MHC抗性基因型有關,綿羊可通過PPA1在機體能量供給中的重要作用來抵抗細粒棘球蚴病[31]。
研究發現,MHC基因與氣味相關,在親緣關系近的個體中MHC基因型差異較小,個體間的氣味也較相近。而動物一般喜歡與氣味差異大的個體交配以避免近交可能帶來的疾病風險,這種行為維持了MHC的多態性,也促進了MHC雜合個體的增加。眾所周知,雜合個體具有的雜合子優勢,其在抵御疾病的能力上更強;且MHC基因是共顯性基因,雜合個體能夠呈遞更多種類的抗原,從而具有更廣泛的抗病性。Schaschl等[32]認為,在雄性中隨年齡增加的DRB雜合子比例表明MHC雜合的雄性壽命比純合子高,這說明MHC基因通過雜合子優勢介導的免疫反應對雄性的重要作用。
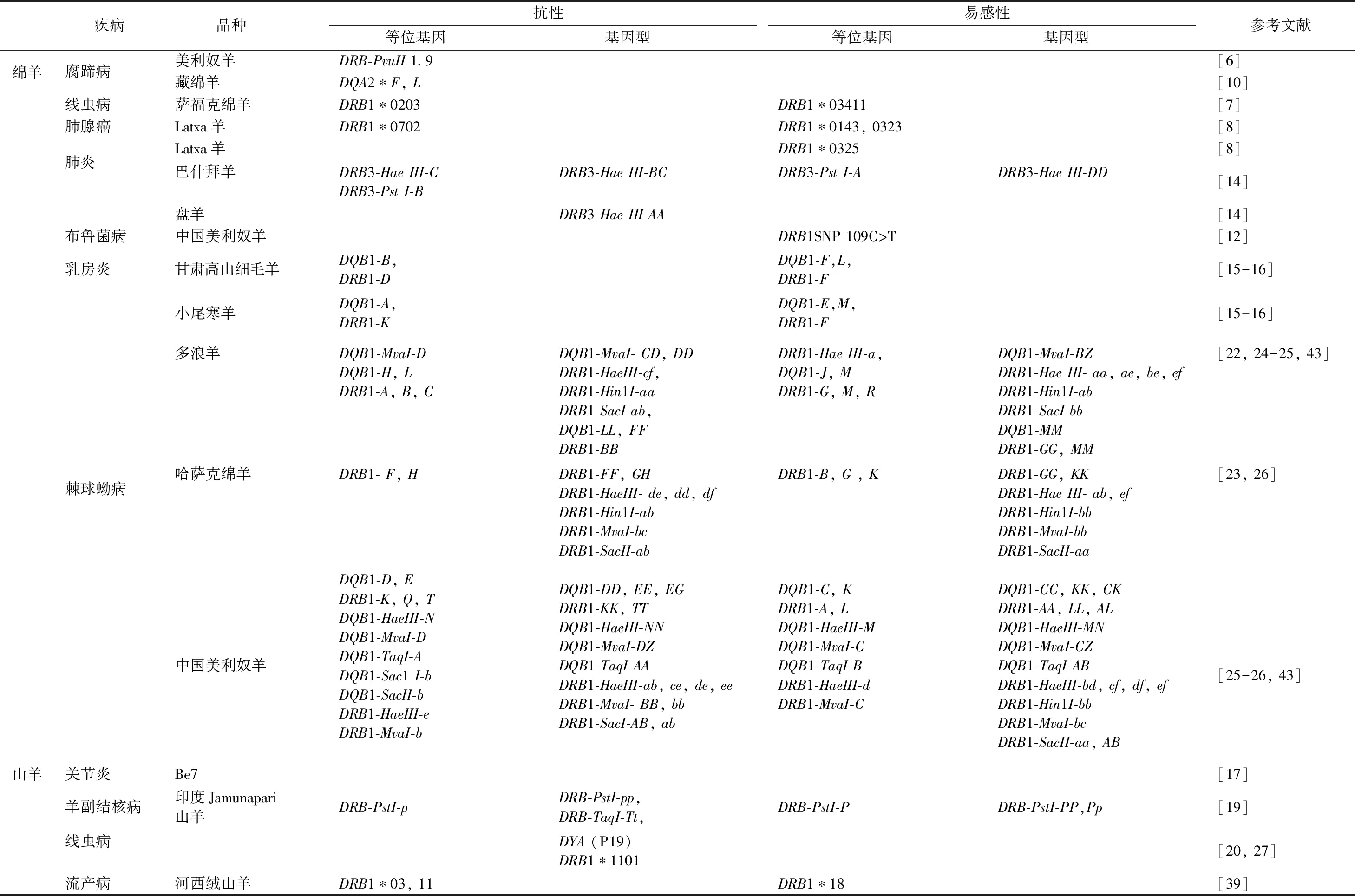
表1 羊MHC基因與疾病的相關性研究匯總
在進化中,宿主與病原體之間能形成“共進化”模式:原本可以抵御病原體的MHC基因型會被不斷進化的病原體適應,以致不再具有較強的抗病性,這時在宿主中如果產生一個新的等位基因,則病原體短期內還不能適應,所以這個新等位基因就可以憑借較強的抗病性在宿主中保留。如此循環就產生了大量的MHC等位基因,可產生出大量非同義替換的SNP從而影響MHC分子的結構和功能[33-34],進而引發對疾病反應的差異。這種新的等位基因可被看作是一種“好基因”,該基因不僅具有抵御病原體的潛力,還可以通過表形吸引異性,從而將“好基因”在種群內傳播,提高群體的免疫力。Eide等[35]對山羊和其他動物的研究發現具有特定的MHC I類等位基因的動物比沒有這一等位基因的動物免疫力更高,說明特定的MHC基因可能通過“好基因”的方式影響著動物的抗病能力。
4 羊MHC基因的應用與前景
近年來,國內外對MHC基因的大量研究對探索某些疾病的免疫性、基因預測和早期預防都有著重要的意義,也為動物的抗病育種提供了理論支撐。
目前,有研究者已將部分發現應用于疾病檢測上。例如綿羊的腐蹄病一般采用藥物防治但不易控制病情還可能有藥物殘留的風險,而國外已將DQA2基因作為抗腐蹄病的標記,并應用于抗病選育的商業化檢測中,林肯大學通過檢測MHC基因的2個基因座位來評估腐蹄病的感染風險,從而可提供具腐蹄病抗性的種羊[36]。我國研究者發明了藏綿羊腐蹄病抗性檢測試劑盒,通過檢測DQA2基因外顯子2的多態性來判斷羊只對腐蹄病的抗性,從而確定是否留種[37]。
在繁殖方面的研究發現,河西絨山羊DQB2基因外顯子3的CF基因型可能與流產抗性有關[38]。羅玉柱等[39]也發現具有DRB1*18等位基因的河西絨山羊更易發生流產,而DRB1*03和DRB1*11等位基因則與流產病的抗性相關;并以此為基礎發明了山羊流產病檢測試劑盒,可快速判斷羊只抗流產能力的強弱從而指導選育。
但是,迄今為止,對羊MHC基因的研究仍不夠全面和深入,大多數研究都集中在II類基因的DQ和DR區,且研究方法也較為單一。因此,如何充分利用我國豐富的綿羊及山羊品種資源,深入挖掘MHC基因和抗病性的聯系,在抗病育種領域和農牧區經濟的提升中都具有十分重大的意義。此外,Yakubu等[40]發現尼日利亞山羊的DRB基因與耐熱性有關。還有研究表明,MHC基因和動物存活率[41]、繁殖、嗅覺、配偶選擇[42]及親緣識別等相關。提示著MHC基因在種群生存發展和抗病育種研究外還有更多有意義的探索方向。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
