春大豆苗期低溫脅迫下葉綠素?zé)晒忭憫?yīng)
2019-12-30 01:45:36田鑫鐘程李性苑
安徽農(nóng)業(yè)科學(xué) 2019年24期
關(guān)鍵詞:大豆
田鑫 鐘程 李性苑
摘要? [目的]探討春大豆與低溫脅迫有關(guān)的光合參數(shù)以及種質(zhì)與低溫之間的相關(guān)性。[方法]以5個(gè)春大豆品種的幼苗為試驗(yàn)材料,用葉綠素?zé)晒鉁y(cè)定儀測(cè)定低溫脅迫后的葉綠素含量。[結(jié)果]5個(gè)春大豆品種的Fv(樣品可變熒光)/Fm(樣品最大熒光產(chǎn)量)和qP(光化學(xué)淬滅系數(shù))均顯著降低;與室溫對(duì)照相比,F(xiàn)0(初始熒光)和qN(非光學(xué)淬滅系數(shù))在低溫脅迫下顯著增加。與其他品種相比,骨粒青的Fo和qN均顯著增大而Fv/Fm顯著降低,qP下降但仍處于高水平。43號(hào)的Fo 增加最少。95-1的Fv/Fm最高,但qP最低。南農(nóng)513的qN增加最少,屬于低溫敏感型材料。光氧化(qN)是引起南農(nóng)513損傷的主要原因,而qP和qN反映出骨粒青能在低溫下繼續(xù)光合作用進(jìn)行生長(zhǎng),以減少光氧化帶來(lái)的損傷。[結(jié)論]qP和qN是用來(lái)評(píng)估大豆的低溫抗性指標(biāo)的較可靠的光合參數(shù)。
關(guān)鍵詞? 大豆;低溫脅迫;葉綠素?zé)晒?光抑制;光氧化
中圖分類號(hào)? S565.1??? 文獻(xiàn)標(biāo)識(shí)碼? A
文章編號(hào)? 0517-6611(2019)24-0028-04
doi:10.3969/j.issn.0517-6611.2019.24.010
Chlorophyll Fluorescence Response of Spring Soybean to Chilling Stress at Seedling Stage
TIAN Xin, ZHONG Cheng, LI Xing-yuan
(College of Comprehensive Health, Kaili University, Kaili, Guizhou 556011)
Abstract? [Objective]To explore correlation among the photosynthetic parameters under the chilling stress, as well as the correlation between germplasm and temperature. [Method]With seedlings of five spring soybean varieties as the test varieties, we detected the chlorophyll contents after temperature stress by chlorophyll fluorescence meter. [Result]Fv/Fm and qP of five spring soybeans all significantly reduced. Compared with the CK under room temperature, F0 and qN enhanced significantly,? and a significant increase in original fluorescence (Fo) and non-photochemical quenching (qN) Following chilling stress compared to room temperature controls. Compared to other soybeans,the Fo and qN increased the greatest and the yield and Fv/Fm decreased the greatest in the tolerant cultivar Guliqing. Interestingly, the qP declined, but remained at a high level. Fo increased the least in cultivar #43, but it still had a high yield. The Fv/Fm ratio was the highest in cultivar 95-1, but had the lowest qP. qN increased the least in chilling-sensitive cultivar Nannong 513. Taken together, photo-oxidation (qN) may be a major cause of damage in Nannong 513, resulting in its chilling sensitivity. In contrast, chilling tolerant cultivar Guliqing may maintain photosynthesis to maintain growth and reduce photo-oxidation damage, as reflected by parameters qP and qN. Thus, qP and qN may be the most reliable parameters to evaluate chilling resistance in soybeans.
Key words? Soybean;Chilling stress;Chlorophyll fluorescence;Photo-inhibition;Photo-oxidation
基金項(xiàng)目? 貴州省科學(xué)技術(shù)基金“貴州省大豆突變體的人工誘導(dǎo)及高產(chǎn)、優(yōu)質(zhì)、多抗品種的選育”(黔科合J字〔2014〕2151)。
作者簡(jiǎn)介? 田鑫(1983—),男,湖南鳳凰人,副教授,博士,從事作物遺傳育種研究。
收稿日期? 2019-06-17
大豆是世界最重要的經(jīng)濟(jì)作物之一,是主要的油料植物。大豆是低溫敏感型作物[1],低溫影響大豆的各個(gè)生育階段,從而降低大豆的產(chǎn)量和品質(zhì)[2-3]。低溫脅迫影響光合機(jī)構(gòu)的性能和活性[4],最嚴(yán)重的危害是在高光效下抑制氣孔開(kāi)度和降低CO2的固定[5],干擾光氧化再生,對(duì)低溫的響應(yīng)可抑制光合作用合成。此外,低溫脅迫可降低電子鏈的氧化還原水平[6],光合作用減弱可降低葉溫、增加光合系統(tǒng)II敏感性,降低PSII電子傳遞和光能轉(zhuǎn)化效率[7],產(chǎn)生光抑制現(xiàn)象[8],導(dǎo)致光氧化降解類囊體[9]。因此,研究低溫脅迫對(duì)光合作用的影響是了解傷害機(jī)理的重要方面[8]。
低溫是我國(guó)南方早春期間常見(jiàn)的逆境脅迫因子,極大影響了春大豆的出芽、成苗及幼苗的生長(zhǎng)發(fā)育,延長(zhǎng)了春大豆的生長(zhǎng)期,對(duì)農(nóng)業(yè)生產(chǎn)造成不利影響,因此有必要對(duì)春大豆材料進(jìn)行低溫耐性機(jī)制研究。目前,對(duì)春大豆的耐低溫研究主要集中在東北三省大豆材料生育期的低溫脅迫,但鮮見(jiàn)我國(guó)南方春大豆材料早春耐低溫的研究報(bào)道。而提高春大豆耐低溫性能力是選育春大豆優(yōu)良種質(zhì)的目標(biāo)之一,通過(guò)快速準(zhǔn)確地評(píng)估春大豆耐低溫能力是低溫育種的重要途徑。葉綠素?zé)晒鈨x可快速檢測(cè)植株在脅迫下的光合作用[10],通過(guò)熒光參數(shù)能夠反映PSⅡ?qū)饽艿奈铡鬟f和耗散狀況等生理過(guò)程。目前,該方法已廣泛用于評(píng)估西瓜[11]、棉花[12]、玉米[13]等作物對(duì)低溫響應(yīng)的動(dòng)態(tài)過(guò)程,但葉綠素?zé)晒鈨x檢測(cè)大豆的低溫耐受力仍鮮見(jiàn)報(bào)道。鑒于此,筆者在低溫脅迫下采用葉綠素?zé)晒鈨x檢測(cè)春大豆光合參數(shù)進(jìn)行抗低溫材料篩選,為早春培育壯苗提供技術(shù)支持,為引種和抗寒種質(zhì)改良和品種選育提供理論依據(jù)。
1? 材料與方法
1.1? 試驗(yàn)材料
對(duì)41份大豆種質(zhì)進(jìn)行低溫篩選,共獲得發(fā)芽率高且苗期長(zhǎng)勢(shì)較好的5個(gè)春大豆品種[14],包括南農(nóng)513、骨粒青、青酥2號(hào)、43號(hào)、95-1。其中,南農(nóng)513為低溫敏感型,骨粒青是最耐低溫型,青酥2號(hào)、43號(hào)和95-1屬于中間型材料。
1.2? 試驗(yàn)方法
1.2.1? 生長(zhǎng)條件。在人工氣候室內(nèi)進(jìn)行常規(guī)種植,土∶沙∶蛭石=4∶2∶1,植株光照采用熒光燈和白熾燈2種,光照時(shí)間為光照∶黑暗=12 h∶12 h(25 ℃/18 ℃,相對(duì)濕度70%),CO2濃度為(398.65 ±20.47)μmol/mol(用Li-6400、LI-COR、USA檢測(cè)獲得),吸收的光通量密度為300 μmol/(m2·s),每2 d澆1次水,共生長(zhǎng)30 d,
1.2.2
低溫處理。當(dāng)幼苗三出復(fù)葉展開(kāi)時(shí),選擇長(zhǎng)勢(shì)一致、生長(zhǎng)狀態(tài)良好、無(wú)病蟲(chóng)害的植株進(jìn)行低溫處理,將每個(gè)材料分成2組,其中一組在低溫培養(yǎng)箱中進(jìn)行低溫處理(5 ℃,相對(duì)濕度70%,光照∶黑暗=12 h∶12 h,連續(xù)處理4 d),CO2濃度為(405.05 ±25.86)μmol/mol,其他條件按常規(guī)進(jìn)行;另一組室溫生長(zhǎng)。
1.2.3? 葉綠素?zé)晒鉁y(cè)定。在第1片三出復(fù)葉完全展開(kāi)時(shí),分別測(cè)定0,1,2,3和4 d低溫處理(5 ℃)的葉綠素?zé)晒鈪?shù),以相應(yīng)的時(shí)間梯度室溫(25 ℃)為對(duì)照。利用紅外氣體分析儀監(jiān)測(cè)平均CO2濃度控制在405 μmol/mol以降低試驗(yàn)誤差,參數(shù)測(cè)定重復(fù)6次,葉綠素?zé)晒鉁y(cè)定按照脈沖幅度調(diào)制熒光計(jì)說(shuō)明書(shū)使用(PAM 2100,H.Waltz,Effeltrich,Germany)[15]。測(cè)定前,葉片先暗適應(yīng)30 min以排除光照誤差。參數(shù)測(cè)定如下:葉綠素?zé)晒猓‵y)、初始熒光(Fo)、最大熒光(Fm);暗適應(yīng)后葉片光適應(yīng)的量子產(chǎn)量閾值、光反應(yīng)階段熒光最大值(Fm)、光反應(yīng)階段光適應(yīng)的最大值(Fm)和光穩(wěn)定值(Ft)。利用這些參數(shù)計(jì)算光化學(xué)反應(yīng)的利用效率:Fv/Fm=(Fm-Fo)/Fm,光反應(yīng)階段光化學(xué)反應(yīng)的光量子產(chǎn)率=(Fm-Ft)/Fm,葉綠素?zé)晒獾墓饣瘜W(xué)淬滅qP=(Fm-Ft)/(Fm-Fo),葉綠素?zé)晒獾姆枪饣瘜W(xué)淬滅qN=(Fm– Fm)/(Fm–Fo)。
1.3? 統(tǒng)計(jì)分析
所有數(shù)據(jù)采用單因素方差分析,差異顯著性用鄧肯測(cè)驗(yàn)。無(wú)特殊注明差異顯著性分析均為P< 0.01。
2? 結(jié)果與分析
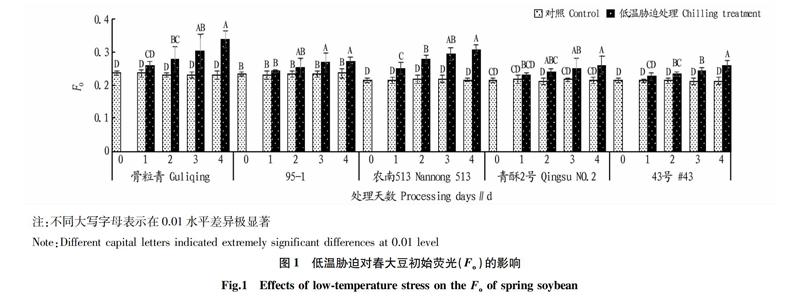
2.1? 低溫脅迫對(duì)春大豆初始熒光(Fo)的影響
由圖1可知,在整個(gè)低溫處理暗適應(yīng)階段,對(duì)照處理的5個(gè)春大豆品種種子的Fo沒(méi)有變化,而低溫處理5個(gè)春大豆品種均產(chǎn)生不同程度變化,說(shuō)明植物體光反應(yīng)中心造成潛在傷害或可逆失活。其中,骨粒青的Fo增加最多,較對(duì)照提高了42%;95-1的Fo增加最少,僅有15%,說(shuō)明低溫脅迫對(duì)不同大豆品種Fo的影響程度不同。骨粒青受低溫脅迫后光反應(yīng)中心嚴(yán)重失活或損害,而95-1受到的傷害最輕。
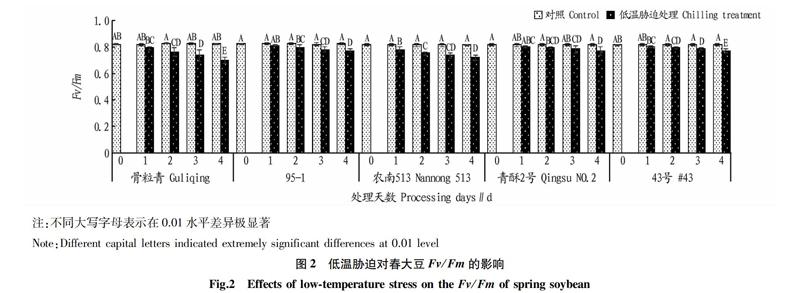
2.2? 低溫脅迫對(duì)春大豆Fv/Fm的影響
由圖2可知,對(duì)照植株在暗適應(yīng)階段Fv/Fm為0.80~0.84。低溫脅迫后,各春大豆品種的Fv/Fm均降低,其中骨粒青下降最多,導(dǎo)致Fo顯著增加、Fm降低;43號(hào)的Fv/Fm降低最小。室溫條件下,春大豆品種對(duì)Fv/Fm沒(méi)有顯著影響,經(jīng)過(guò)低溫脅迫的春大豆品種Fv/Fm存在極顯著差異。不同春大豆品種中,骨粒青的光反應(yīng)中心受到的傷害最多,而43號(hào)最少。
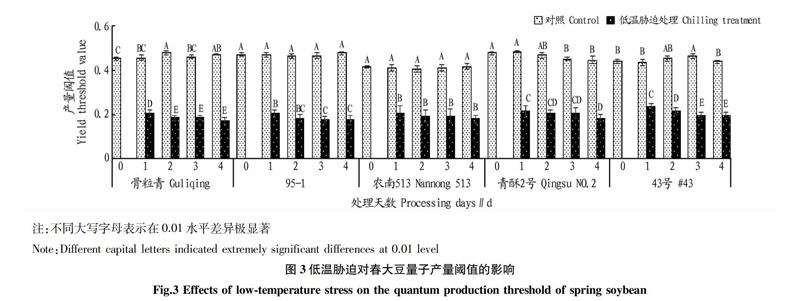
2.3? 低溫脅迫對(duì)春大豆量子產(chǎn)量閾值、qP和qN的影響
量子產(chǎn)量閾值是光系統(tǒng)II中光化學(xué)能轉(zhuǎn)化成光量子的效率。由圖3可知,低溫脅迫下5個(gè)春大豆品種的量子產(chǎn)量閾值均急劇降低,說(shuō)明暗適應(yīng)階段春大豆光合效率很低。其中,骨粒青量子產(chǎn)量閾值下降最多,而43號(hào)下降最少。室溫處理下,不同春大豆品種間量子產(chǎn)量閾值存在極顯著差異,說(shuō)明低溫處理量子產(chǎn)量閾值受種質(zhì)和溫度的交互影響。
2.4? 低溫脅迫對(duì)春大豆qP的影響
葉綠素?zé)晒獾墓饣瘜W(xué)淬滅(qP)是光系統(tǒng)II中天線色素吸收能量的比例。由圖4可知,低溫脅迫下5個(gè)春大豆品種的qP均顯著降低。其中,低溫處理下春大豆品種骨粒青的qP最高,而95-1最低。室溫處理下,南農(nóng)513暗適應(yīng)階段的qP較低。
2.5? 低溫脅迫對(duì)春大豆qN的影響
由圖5可知,低溫脅迫下,骨粒青的葉綠素?zé)晒獾姆枪饣瘜W(xué)淬滅(qN)極顯著高于其他春大豆品種,而南農(nóng)513的qN最低。由此可見(jiàn),骨粒青通過(guò)輻射耗散釋放多余能量,從而避免光氧化傷害;南農(nóng)513保持相對(duì)較高的光合作用,受到嚴(yán)重的光氧化傷害,導(dǎo)致不同春大豆品種間耐低溫能力有差異。
3? 結(jié)論與討論
3.1? 低溫脅迫下的Fo、Fv/Fm和閾值
前人研究表明,光合作用的Fv/Fm可作為判斷低溫脅迫和低溫抗性的依據(jù)[16-19],F(xiàn)v/Fm還適用于低溫抗性植株或低溫抗性研究。此外,越來(lái)越多的證據(jù)表明Fv/Fm適用于低溫不敏感型作物,并且不能把參數(shù)作為篩選抗低溫植物的標(biāo)準(zhǔn)[18]。光反應(yīng)中心嚴(yán)重受損時(shí),F(xiàn)v/Fm不一定會(huì)下降,通常Fv/Fm降低是由于光反應(yīng)中心受低溫冷害在短期內(nèi)暫時(shí)失活以避免光氧化造成的傷害,短期(0~7 d)內(nèi)溫度降到0 ℃以上時(shí),植物因缺乏能量而生長(zhǎng)暫時(shí)受到抑制,然而冰點(diǎn)溫度嚴(yán)重影響細(xì)胞組織,植物體內(nèi)結(jié)冰導(dǎo)致組織緊縮和細(xì)胞濃度增大,細(xì)胞體內(nèi)有毒離子積累和細(xì)胞膜損傷造成細(xì)胞死亡[19],冷害比凍害的危害要輕一些,由于沒(méi)有結(jié)冰的影響,盡管細(xì)胞膜的流動(dòng)性減少,短期冷害是可以恢復(fù)的。但長(zhǎng)期低溫條件下質(zhì)壁分離不能維持正常離子濃度,細(xì)胞代謝紊亂導(dǎo)致細(xì)胞死亡[20-21]。
因此,大豆受低溫冷害時(shí)會(huì)影響光反應(yīng)中心的光利用效率,F(xiàn)o、Fv/Fm和閾值參數(shù)能較好反應(yīng)光反應(yīng)中心的活性,但與低溫冷害無(wú)相關(guān)性。該試驗(yàn)結(jié)果表明,骨粒青的低溫抗性表現(xiàn)在隨著低溫時(shí)間的延長(zhǎng),光反應(yīng)中心活性逐漸降低,兩者間的上升和下降表明光合能力并不是決定低溫抗性主要因素,F(xiàn)o、Fv/Fm和閾值參數(shù)并不能作為評(píng)估大豆低溫抗性的可靠依據(jù)。
3.2? 低溫脅迫下的qP和qN
低溫敏感植物的光合作用和光飽和點(diǎn)在低溫脅迫時(shí)急劇下降,白天光照過(guò)強(qiáng)會(huì)導(dǎo)致光抑制,引起光系統(tǒng)II中的捕光色素蛋白復(fù)合物構(gòu)象改變和降解[22],光抑制是通過(guò)吸收的能量過(guò)度耗散或光系統(tǒng)II D1反應(yīng)中心蛋白損壞造成光系統(tǒng)II可逆降解[23]。此外,光抑制與能量過(guò)度耗散和光合結(jié)構(gòu)受損過(guò)程有關(guān),是植物體自身的保護(hù)機(jī)制[18,24]。qN表示能量以熱能形式耗散,反映光強(qiáng)過(guò)強(qiáng)產(chǎn)生光抑制,如果光抑制較低,光合系統(tǒng)損害較小;qP是衡量開(kāi)放的光系統(tǒng)II反應(yīng)中心的光抑制程度。綜上所述,qP和qN可以判斷植物自我保護(hù)能力。
此外,光抑制、低溫、光強(qiáng)增強(qiáng)可能對(duì)光合膜吸收能力造成損害,引起類囊體光氧化降解[25],黑暗低溫傷害的程度不同,光和氧氣依賴于光氧化表明光合作用的光反應(yīng)可能產(chǎn)生一種或多種有毒的活性氧,這些活性氧對(duì)生物體造成明顯傷害。植物通過(guò)進(jìn)化葉綠體的淬滅機(jī)制來(lái)減少有毒物質(zhì)對(duì)機(jī)體的破壞,有30%~40%的細(xì)胞抗壞血酸、10%~50%的谷胱甘肽以及所有的α維生素E和β胡蘿卜素都位于葉綠體,雖然它們只占葉肉細(xì)胞體積的3%~4%[26],但這些復(fù)合物能夠清除植物活性氧。報(bào)道顯示,低溫敏感型植物容易產(chǎn)生活性氧[26],此外低溫敏感型植物在低溫、抗氧化劑環(huán)境下不能處理積累的活性氧,并且在早期會(huì)因過(guò)氧化氫的產(chǎn)生而導(dǎo)致機(jī)體受損,光氧化是植物發(fā)生氧化傷害的一種機(jī)制。
該研究結(jié)果顯示,與其他4個(gè)大豆品種相比,在低溫冷害期骨粒青的qN增加幅度較大,且qP始終處于較高水平,說(shuō)明骨粒青的光合效率較低,但光系統(tǒng)II反應(yīng)中心仍然能使光合作用產(chǎn)生能量維持植物的正常生長(zhǎng)。此外,能量過(guò)度耗散能有效抑制QA的過(guò)度降低[27],有效減少光氧化帶來(lái)的傷害,骨粒青的qP和qN最高,南農(nóng)513的 qN較其他材料極顯著降低,說(shuō)明南農(nóng)513能吸收更多能量,產(chǎn)生大量活性氧,導(dǎo)致存活率降低,此外還發(fā)現(xiàn)過(guò)度的能量耗散和抗氧化是判斷大豆低溫抗性的最重要因素。
3.3? 葉綠素?zé)晒庠u(píng)估大豆的低溫抗性
早春時(shí)植物易受低溫、光抑制、光氧化以及其他因素的交替影響,如溫度、降雨、光照、濕度、風(fēng)、病害等。人為低溫脅迫試驗(yàn)允許操控植物低溫的耐受性,有研究用Fv/Fm來(lái)評(píng)估植物的低溫抗性[16-17,22,24],這種方法適用于生長(zhǎng)在低溫條件下的低溫抗性材料,通過(guò)其光合效率的不同機(jī)制來(lái)提供能量,從而維持長(zhǎng)期低溫冷害。因此,低溫抗性植物繼續(xù)進(jìn)行光合作用以維持抗性機(jī)制,盡管低溫敏感型植物可以耐受短期低溫,但不能長(zhǎng)期在低溫條件下存活,低溫敏感型植物面臨的最大問(wèn)題是氧化還原系統(tǒng)紊亂和活性氧傷害。因此,低溫敏感型植物短期低溫下存活的重要因素是具有有效的抗氧化系統(tǒng)。
植物的抗氧化系統(tǒng)很復(fù)雜,然而葉綠素?zé)晒夥从吵隹寡趸到y(tǒng)在某種程度上可以代替現(xiàn)有的低溫測(cè)試,植物存活分析較敏感和客觀,而低溫測(cè)試存在較大的誤差和延遲[10]。與離子釋放測(cè)試試驗(yàn)相比,葉綠素?zé)晒夥椒ň哂胁黄茐男院秃?jiǎn)單易測(cè)的特點(diǎn),因此葉綠素?zé)晒膺m用于育種中評(píng)估大規(guī)模材料的低溫抗性,但葉綠素?zé)晒鉁y(cè)試的材料具有種間差異,如耐低溫作物燕麥的Fv/Fm和低溫敏感型作物大豆的qP和qN均可測(cè)。
參考文獻(xiàn)
[1]ROHACˇEK K.Chlorophyll fluorescence parameters:The definitions,photosynthetic meaning,and mutual relationships[J].Photosynthetica,2002,40(1):13-29.
[2]BALESTRASSE K B,TOMARO M L,BATLLE A,et al.The role of 5-aminolevulinic acid in the response to cold stress in soybean plants[J].Phytochemistry,2010,71(17/18):2038-2045.
[3]張大偉,杜翔宇,劉春燕,等.低溫脅迫對(duì)大豆萌發(fā)期生理指標(biāo)的影響[J].大豆科學(xué),2010,29(2):228-232.
[4]許耀照,曾秀存,張芬琴,等.低溫對(duì)白菜型冬油菜幼苗葉綠素?zé)晒鈫?dòng)過(guò)程與品種抗寒性的影響[J].河西學(xué)院學(xué)報(bào),2016,32(5):60-68.
[5]呂星光,周夢(mèng)迪,李敏.低溫脅迫對(duì)甜瓜嫁接苗及自根苗光合及葉綠素?zé)晒馓匦缘挠绊慬J].植物生理學(xué)報(bào),2016,52(3):334-342.
[6]余麗玲,陳凌艷,何天友,等.低溫脅迫對(duì)西洋杜鵑葉片葉綠素?zé)晒鈪?shù)的影響[J].西南林業(yè)大學(xué)學(xué)報(bào),2013,33(6):1-6.
[7]王春萍,雷開(kāi)榮,李正國(guó),等.低溫脅迫對(duì)水稻幼苗不同葉齡葉片葉綠素?zé)晒馓匦缘挠绊慬J].植物資源與環(huán)境學(xué)報(bào),2012,21(3):38-43.
[8]陳梅,唐運(yùn)來(lái).低溫脅迫對(duì)玉米幼苗葉片葉綠素?zé)晒鈪?shù)的影響[J].內(nèi)蒙古農(nóng)業(yè)大學(xué)學(xué)報(bào)(自然科學(xué)版),2012,33(3):20-24.
[9]尹賾鵬,劉雪梅,商志偉,等.不同干旱脅迫下歐李光合及葉綠素?zé)晒鈪?shù)的響應(yīng)[J].植物生理學(xué)報(bào),2011,47(5):452-458.
[10]侯偉,孫愛(ài)花,楊福孫,等.低溫脅迫對(duì)西瓜幼苗光合作用與葉綠素?zé)晒馓匦缘挠绊慬J].廣東農(nóng)業(yè)科學(xué),2014,41(13):35-39.
[11]武輝,戴海芳,張巨松,等.棉花幼苗葉片光合特性對(duì)低溫脅迫及恢復(fù)處理的響應(yīng)[J].植物生態(tài)學(xué)報(bào),2014,38(10):1124-1134.
[12]YIN Z T,MENG F F,SONG H N,et al.Mapping quantitative trait loci associated with chlorophyll a fluorescence parameters in soybean(Glycine max(L.) Merr.)[J].Planta, 2010, 231(4):875-885.
[13]陳梅,唐運(yùn)來(lái).低溫脅迫對(duì)玉米幼苗葉片葉綠素?zé)晒鈪?shù)的影響[J].內(nèi)蒙古農(nóng)業(yè)大學(xué)學(xué)報(bào)(自然科學(xué)版),2012,33(3):20-24.
[14]田鑫,何小玲,顧衛(wèi)紅,等.5份春大豆苗期耐低溫性的鑒定及評(píng)價(jià)[J].大豆科學(xué),2013,32(6):755-762.
[15]RIZZA F,PAGANI D,STANCA A M,et al.Use of chlorophyll fluorescence to evaluate the cold acclimation and freezing tolerance of winter and spring oats[J].Plant breeding,2010,120(5):389-396.
[16]楊萬(wàn)基,蔣欣梅,高歡,等.28-高蕓苔素內(nèi)酯對(duì)低溫弱光脅迫辣椒幼苗光合和熒光特性的影響[J].南方農(nóng)業(yè)學(xué)報(bào),2018,49(4):741-747.
[17]張玲,王華,汪天,等.不同桉樹(shù)品種在低溫脅迫下的生理反應(yīng)[J].安徽農(nóng)業(yè)大學(xué)學(xué)報(bào),2014,41(3):456-461.
[18]李慶會(huì),徐輝,周琳,等.低溫脅迫對(duì)2個(gè)茶樹(shù)品種葉片葉綠素?zé)晒馓匦缘挠绊慬J].植物資源與環(huán)境學(xué)報(bào),2015,24(2):26-31.
[19]王兆,劉曉曦,鄭國(guó)華.低溫脅迫對(duì)彩葉草光合作用及葉綠素?zé)晒獾挠绊慬J].浙江農(nóng)業(yè)學(xué)報(bào),2015,27(1):49-56.
[20]于永暢,張林,王厚新,等.ABA和PP333對(duì)國(guó)蘭低溫脅迫及恢復(fù)中光合作用和葉綠素?zé)晒鈪?shù)的影響[J].農(nóng)學(xué)學(xué)報(bào),2014,4(4):30-37.
[21]王春萍,黃啟中,雷開(kāi)榮,等.低溫弱光下辣椒幼苗葉綠素?zé)晒馓匦约捌渑c品種耐性的關(guān)系[J].園藝學(xué)報(bào),2015,42(9):1798-1806.
[22]SHARMA P,SHARMA N,DESWAL R.The molecular biology of the low-temperature response in plants[J].Bio Essays,2005,27(10):1048-1059.
[23]肖春燕,邢瀟晨,劉會(huì)芳,等.低溫下NO對(duì)黃瓜光合熒光及抗氧化特性的影響[J].核農(nóng)學(xué)報(bào),2014,28(6):1083-1091.
[24]盧廣超,許建新,薛立,等.低溫脅迫對(duì)4種幼苗的葉綠素?zé)晒馓匦缘挠绊慬J].中南林業(yè)科技大學(xué)學(xué)報(bào),2014,34(2):44-49.
[25]杜曉華,齊陽(yáng)陽(yáng),李建飛,等.低溫脅迫對(duì)大花三色堇和角堇光合作用與葉綠素?zé)晒獾挠绊慬J].西北農(nóng)業(yè)學(xué)報(bào),2017,26(10):1499-1506.
[26]黃璇.夜間低溫和短時(shí)高溫對(duì)不同烤煙品種生長(zhǎng)影響的研究[D].長(zhǎng)沙:湖南農(nóng)業(yè)大學(xué),2014.
[27]陶宏征,趙昶靈,李唯奇.植物對(duì)低溫的光合響應(yīng)[J].中國(guó)生物化學(xué)與分子生物學(xué)報(bào),2012,28(6):501-508.
猜你喜歡
農(nóng)業(yè)科技通訊(2023年1期)2023-02-12 07:09:18
今日農(nóng)業(yè)(2022年16期)2022-11-09 23:18:44
中國(guó)化肥信息(2022年7期)2022-08-31 01:29:28
中國(guó)化肥信息(2022年5期)2022-08-30 01:58:26
今日農(nóng)業(yè)(2021年20期)2021-11-26 01:23:56
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農(nóng)產(chǎn)品市場(chǎng)周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫(huà)報(bào)·智力大王(2015年10期)2016-01-27 01:01:35
讀寫(xiě)算(中)(2015年10期)2015-11-07 07:24:12

- 安徽農(nóng)業(yè)科學(xué)的其它文章
- 西藏高校植物組織培養(yǎng)技術(shù)教學(xué)改革的研究
- 農(nóng)業(yè)大學(xué)資源環(huán)境類本科生科教融合培養(yǎng)模式的實(shí)踐
- 林木組織培養(yǎng)技術(shù)農(nóng)事訓(xùn)練課程探索與實(shí)踐
- 加強(qiáng)校企合作培養(yǎng)食品科學(xué)與工程專業(yè)研究生科研能力的策略探索
- 模塊化課程在綜合氣象業(yè)務(wù)崗位培訓(xùn)中的應(yīng)用
- 加強(qiáng)對(duì)外合作交流助力高水平農(nóng)業(yè)科研院所建設(shè)