錳脅迫對黃花草種子萌發及幼苗生理生化特征的影響
2019-12-20 02:41:30肖澤華李欣航潘高吳耀文楊燦鑫匡雪韶劉文勝
草業學報 2019年12期
關鍵詞:植物
肖澤華,李欣航,潘高,吳耀文,楊燦鑫,匡雪韶,劉文勝*
(1.中南林業科技大學生命科學與技術學院,湖南 長沙 410004;2.湖南省環境資源植物開發與利用工程技術研究中心,湖南 長沙 410004)
錳作為植物生長所必需的微量元素之一,參與植物光合作用、酶促反應和氧化還原等活動,在植物生命活動中具有不可替代的作用[1]。過量的錳則抑制植物生長,甚至通過食物鏈危害人類健康[2]。我國因錳礦開采等原因造成了大面積的錳污染區,嚴重影響人民生產生活。因而,治理錳礦區污染刻不容緩。植物修復作為一種成本低、適應范圍廣、環境擾動小且治理效果持久的重金屬修復技術,是環境領域的研究熱點[3]。因此,植物修復技術是錳污染治理的重要選擇。
植物修復的前提是篩選出對重金屬具有耐性,且經濟利用價值較高的植物。種子萌發至幼苗生長是植物對環境脅迫最敏感的時期,是篩選修復植物的較好時期。脅迫環境下植物的生長往往會受到抑制,主要表現為生物量、葉綠素含量下降及積累大量丙二醛(malondialdehyde,MDA),而抗氧化酶活性及滲透調節物質等的變化是植物對脅迫的直接反應,體現了植物對脅迫的耐性,是其適應脅迫環境的重要依據[4]。抗氧化酶系統主要由超氧化物歧化酶(superoxide dismutase,SOD)、過氧化物酶(peroxidase,POD)和過氧化氫酶(catalase,CAT)組成,SOD是一種可清除植物體內具有強烈毒性超氧陰離子自由基的酶,反應產物H2O2可被CAT進一步分解或被POD利用。滲透調節物質中的脯氨酸、可溶性蛋白和可溶性糖在植物適應重金屬脅迫中發揮著重要作用,能夠維持細胞的含水率和滲透勢,保護生物大分子,降低因滲透失水所帶來的損傷。因此,研究種子萌發和幼苗生理的變化特征對污染土壤的修復具有實際意義。
黃花草(Cleomeviscosa)為山柑科(Capparaceae)白花菜屬(Cleome)的一種一年生草本植物,在全世界熱帶與亞熱帶均廣泛分布,主要分布在我國南方各省。該植物生物量大、株型美觀、生長速度快、抗逆性強、種子量大,可作為植被恢復的先鋒物種[5]。通過在湖南湘潭錳尾礦實地調研,發現該植物是錳礦廢棄地優勢植物之一,在該錳礦廢棄地能正常開花結實,是一種值得開發用于錳尾礦植物修復的材料。目前,對黃花草的研究較少,主要集中于種子萌發特性、植被恢復和藥用價值等方面[5-6],關于錳脅迫下種子萌發、幼苗生長及生理特性的研究尚未見報道。
本研究根據不同錳濃度脅迫下黃花草種子的萌發特性、幼苗生長和生理特征,探明該植物在錳脅迫下的生理耐受機制,研究結果可作為認識植物生長適應錳污染的基礎數據,同時為利用黃花草作為錳污染的修復植物提供理論依據。
1 材料與方法
1.1 試驗材料
供試黃花草種子于2016年11月下旬種子成熟時期采于湖南省湘潭錳礦區。該礦是我國開發最早的錳礦之一,礦區廢棄地污染嚴重。現場調查顯示,該地黃花草長勢良好,以草叢形式在多處出現。選擇該地區長勢相同的100株黃花草,采集其種子并置于信封帶回實驗室室溫保存備用。
1.2 試驗方法
1.2.1種子萌發試驗 挑選顆粒飽滿、大小及色澤一致的種子進行萌發試驗。將種子在0.5%的KMnO4溶液中浸泡30 min,然后用去離子水徹底清洗干凈。根據趙懷寶等[5]和預試驗的研究結果,本研究認為500 mg·L-1赤霉素(gibberellin,GA)溶液浸種能提高黃花草種子的萌發率。為檢驗赤霉素的效果,一批種子在500 mg·L-1赤霉素溶液中浸泡24 h;作為對照,另一批種子則在去離子水中浸泡24 h。然后將種子置于直徑為120 mm的培養皿中,其內裝200 mL經過清洗滅菌及過2 mm篩的沙子。參照潘高等[7]的方法,利用MnCl2·4H2O分別設置0,1000,5000,10000,15000,20000 μmol·L-1(分別記為CK、T1、T2、T3、T4、T5,下同) 6個錳濃度(組)。每組處理5個重復,每個培養皿30粒種子。將培養皿放置在人工氣候培養箱,設置溫度為(30±0.5)℃,12 h光照/12 h黑暗,相對濕度為70%。每天添加去離子水保持培養基質濕潤。播種后每天定時觀察并記錄種子萌發情況,以胚根伸出種皮且長度達到種子直徑1/2作為種子萌發的標志。連續統計萌發率直至連續3 d無新種子萌發即認為萌發結束,在第7天統計發芽勢,第16天統計萌發率。萌發結束后測定黃花草幼苗的根長、芽長、鮮重、干重,并計算發芽指數、活力指數、根冠比。
1.2.2幼苗生長試驗設計 幼苗生長試驗采用盆栽沙培法,盆缽內裝200 mL過2 mm篩的潔凈沙子,置于與萌發試驗相同條件的人工氣候培養箱中。挑選與萌發試驗同一批的黃花草種子進行播種,每盆10粒種子。每天添加去離子水保持種子萌發環境所需水分,幼苗長出2至4片真葉時用Hoagland營養液進行培養。設置與萌發試驗相同的錳濃度處理,每個處理重復10次,待長出6至8片真葉且長勢相同時利用MnCl2·4H2O進行錳脅迫[8]。分別在脅迫后的第7、15和30天時選取黃花草幼苗中、上部位的葉片進行生理生化試驗。
1.3 測定項目和方法
1.3.1種子萌發指標測定 黃花草種子萌發相關指標的測定參考黃玉梅等[8]的方法。
發芽率=16 d內正常發芽的種子數/供試種子數×100%
發芽勢=7 d內正常發芽種子數/供試種子數×100%
發芽指數(GI)=∑(Gt/Dt)
活力指數(VI)=GI×S
式中:Gt為在t日的發芽種子數;Dt為發芽天數;S為幼苗高度。
1.3.2幼苗生物量指標測定 萌發第16天時,每個濃度處理隨機選取10株幼苗,用精度為0.1 mm的游標卡尺測定幼苗的根長、芽長;用濾紙吸去幼苗表面的水分后稱量鮮重,再于95 ℃下殺青10 min,70 ℃下烘至恒重。
根冠比=根干重/地上部干重
1.3.3生理生化指標測定 采用丙酮浸提法測定葉綠素含量[9];采用蒽酮比色法測定可溶性糖含量[9];采用考馬斯亮藍法測定可溶性蛋白含量[9];采用酸性茚三酮顯色法測定游離脯氨酸含量[9];采用硫代巴比妥酸法測定丙二醛含量[9];采用氮藍四唑法測定超氧化物歧化酶活性[9];采用愈創木酚法測定過氧化物酶活性[9];采用紫外吸收法測定過氧化氫酶活性[9]。各指標測定重復3次,每次稱取新鮮黃花草葉片0.3 g。
1.4 數據處理
種子萌發及幼苗生理各指標的數據結果均以“平均值±標準差”表示。采用SPSS 19.0統計軟件對數據進行單因素方差分析(One-way ANOVA),并利用最小顯著差數法(LSD法)對每個指標不同處理間的差異顯著性進行多重比較。最后使用Sigma Plot 12.5作圖。
2 結果與分析
2.1 錳脅迫對黃花草種子萌發及幼苗生長的影響
在去離子水浸種的處理下,黃花草種子的發芽勢、發芽指數和活力指數隨著錳濃度的增加(表1),總體呈下降趨勢。T1處理與對照相比黃花草種子萌發各項指標差距不大,但T2~T5處理與對照和T1相比均形成顯著性差異。T2~T5處理使黃花草種子的發芽率分別降低了38.78%,36.72%,28.56%,30.60%。可見當錳濃度達到T2處理時,對黃花草種子的萌發產生較強的抑制作用。在500 mg·L-1赤霉素浸種的處理下,黃花草種子的發芽勢、發芽率、發芽指數和活力指數均隨著錳濃度的增加,呈現總體下降的趨勢,且比去離子水浸種處理的下降幅度大。CK(GA)、T1(GA)和T2(GA)處理的各項萌發指標均顯著高于CK、T1和T2處理,可見赤霉素浸種對黃花草種子在較低濃度錳處理下的萌發具有促進作用。T5(GA)處理下的黃花草種子發芽率與T5無顯著差異,但發芽勢、發芽指數和活力指數均顯著低于T5處理,分別降低了41.94%,29.52%,85.79%。可見當錳濃度達到T5處理時,赤霉素浸種處理對黃花草種子的萌發產生了較強的抑制作用。
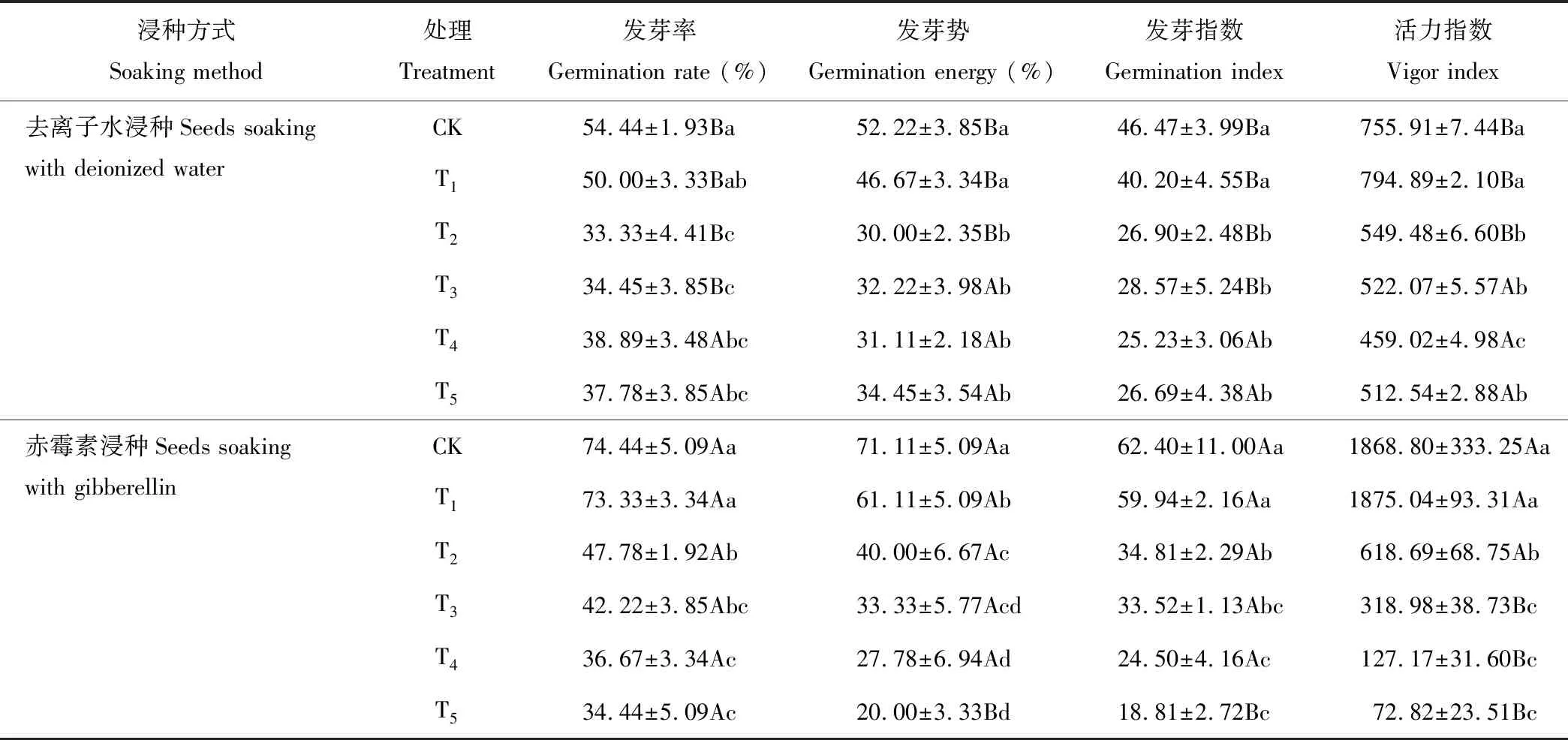
表1 錳脅迫對黃花草種子萌發的影響
注:同列不同大寫字母分別表示不同浸種處理下相同錳處理之間差異顯著,不同小寫字母分別表示相同浸種處理下不同錳處理下之間差異顯著(P<0.05),下同。
Note: Different capital letters within the same column show significant differences among the same manganese treatments under different soaking treatments, different lowercase letters within the same column show significant differences among different manganese treatments under the same soaking treatments (P<0.05). The same below.
幼苗生長是植物對脅迫響應的綜合體現[10]。隨著錳濃度的增加,黃花草幼苗芽長、根長、生物量和根冠比均呈現先升高后下降的變化趨勢(表2),且均在T2處理達到最大值,在T5處理達到最低值。說明低濃度錳對黃花草幼苗生長具有促進作用,高濃度則有抑制作用。
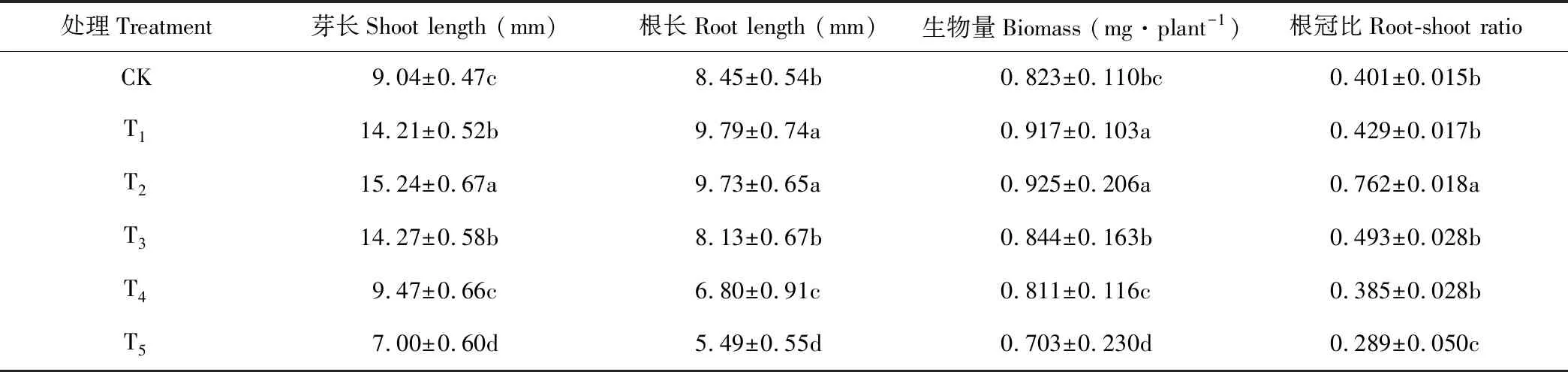
表2 錳脅迫對黃花草幼苗生長的影響
2.2 錳脅迫對黃花草幼苗葉綠素含量的影響
葉片葉綠素含量是反映植物光合能力的重要指標。在錳脅迫7 d時,黃花草幼苗葉綠素a、葉綠素b、葉綠素a+b含量及葉綠素a/b值隨錳濃度的增加均呈現先上升后下降的趨勢(表3),其中,T2處理顯著提高了葉綠素含量,T3處理葉綠素a/b的值最高,呈現顯著性差異。在處理15 d時,隨著錳濃度的增加黃花草幼苗葉片葉綠素a、葉綠素b及葉綠素a+b含量值均呈先下降后升高的變化趨勢,且在T3處理達到最低值,T5處理的葉綠素a、葉綠素a+b、葉綠素a/b值均比對照組要高,且差異達到顯著水平,表明T3、T4和T5處理對黃花草葉綠素的合成產生了較大影響。在錳脅迫30 d時,T1處理下葉綠素a、葉綠素b、葉綠素a+b含量及葉綠素a/b值達到峰值,分別比對照增加了43.6%,2.9%,35.1%,38.4%,T5處理葉綠素a、葉綠素b、葉綠素a+b含量降到最低,葉綠素a含量降幅較大。整體上,T1、T2處理可增加黃花草葉綠素含量,T1增加效果最顯著;T5處理會降低葉綠素含量,可能是因為高濃度錳抑制了黃花草葉綠素的合成。
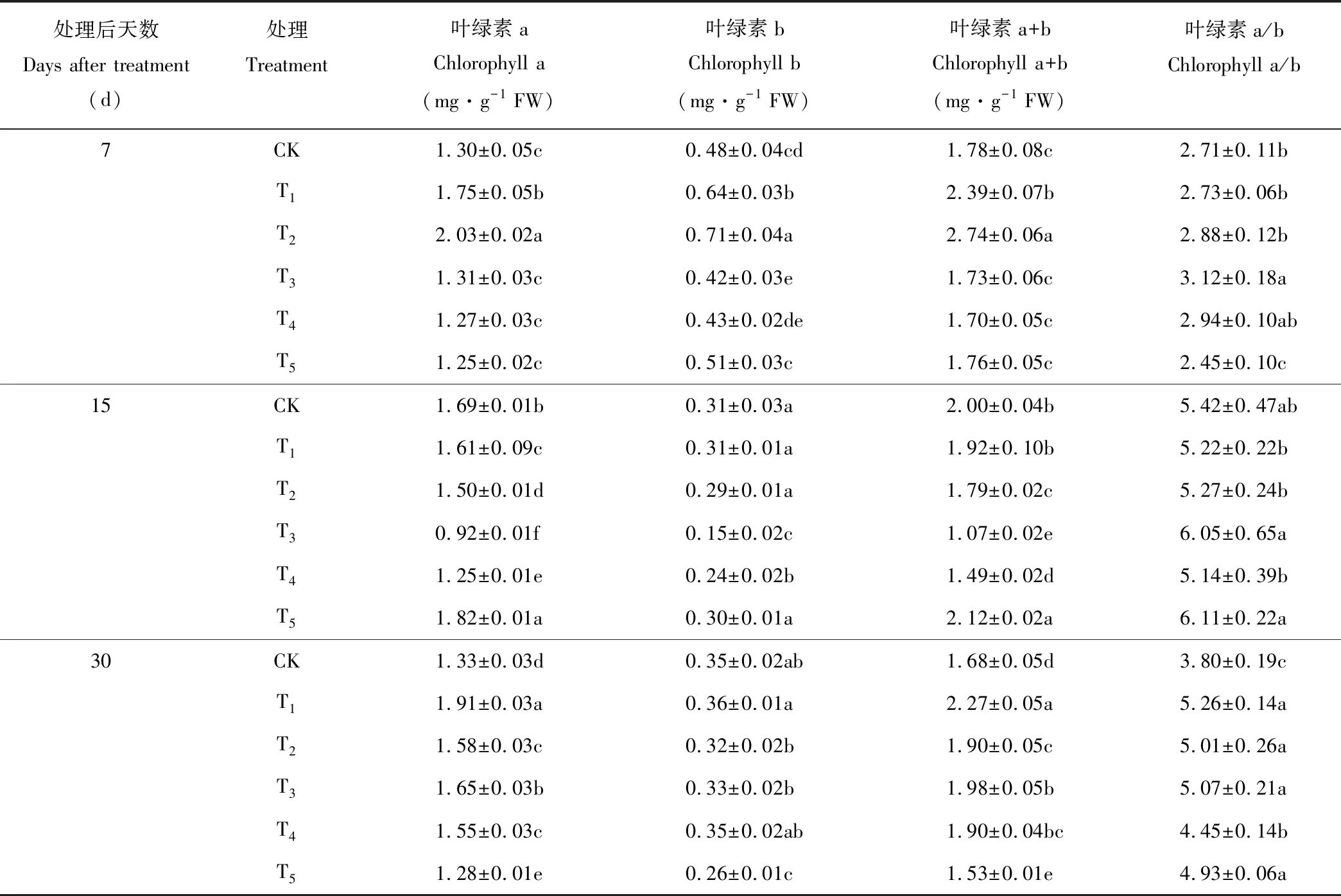
表3 錳脅迫對黃花草幼苗葉綠素含量的影響
2.3 錳脅迫對黃花草幼苗抗氧化酶活性的影響
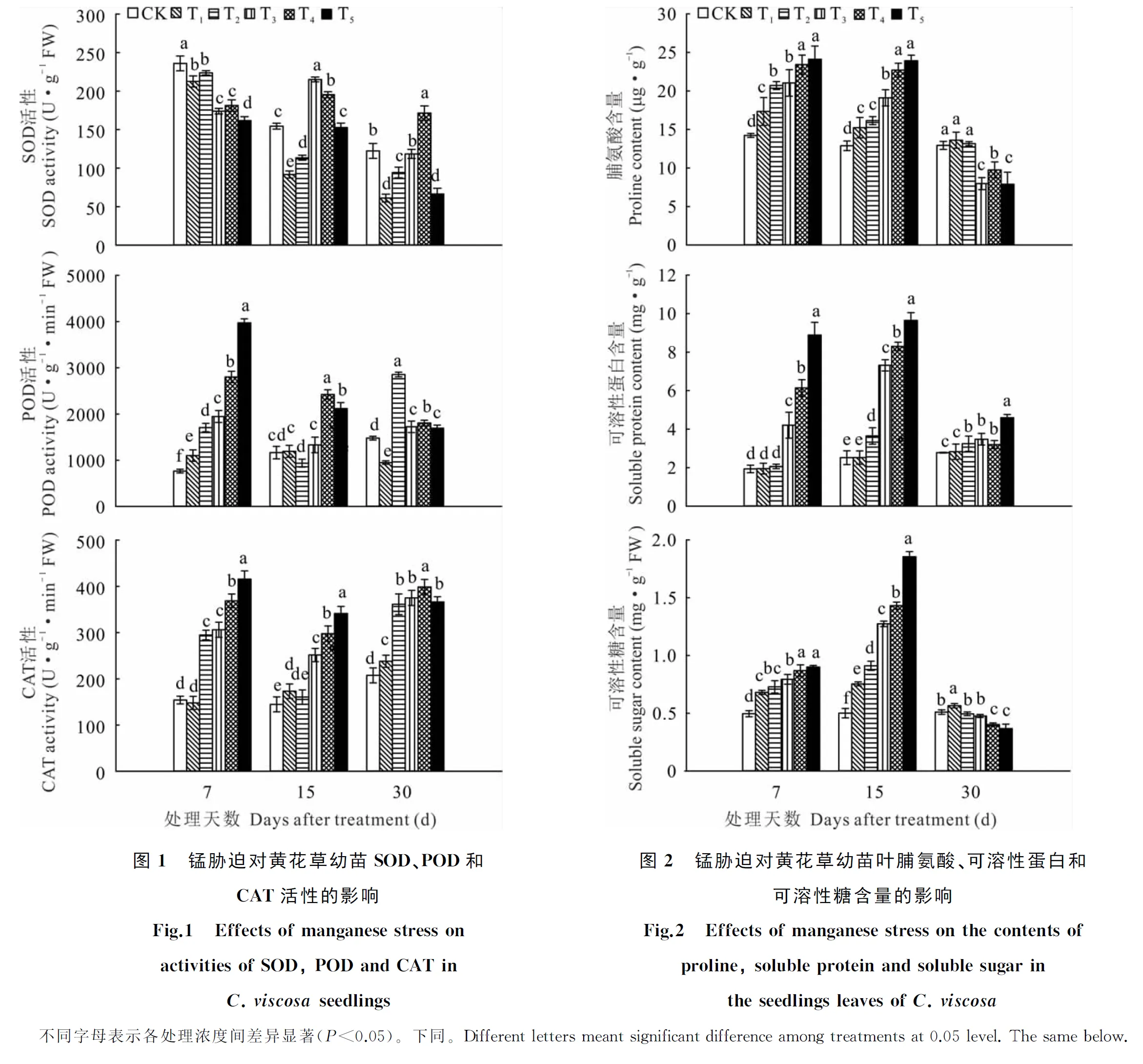
黃花草幼苗葉片SOD、POD和CAT活性在處理至第7天時具有明顯的變化趨勢(圖1),但隨處理時間延長,3種抗氧化酶活性的變化趨勢則無明顯規律。黃花草葉片SOD活性在脅迫第7天時,隨處理濃度的增加呈下降趨勢;15 d時,T3、T4處理下SOD活性達到最高值,T5處理與對照差異不顯著;30 d時,只有T4處理下的SOD活性高于對照,T5比對照低45.7%。黃花草葉片POD的活性在處理第7天時隨濃度的增加呈上升趨勢,處理間差異顯著;15 d時,T2~T5處理下的POD活性均分別低于第7天時各處理活性;在30 d時,T2處理達到該處理最大值,T4、T5處理繼續下降。隨處理時間的延長,T1處理的CAT活性逐漸升高;T2~T5處理下黃花草葉片CAT活性呈先下降后升高的變化趨勢,T2~T4處理的最高值則出現在第30天,T5處理的最高值出現在第7天。結果表明,黃花草幼苗3種抗氧化酶活性在錳脅迫初期較高,可緩解高濃度錳脅迫造成的傷害,但隨著脅迫時間的延長SOD和POD的活性出現不同程度的下降,而CAT的活性有所提高,說明高濃度錳脅迫后期黃花草的抗氧化能力衰退,CAT起清除活性氧的關鍵作用。
2.4 錳脅迫對黃花草幼苗葉片脯氨酸、可溶性蛋白、可溶性糖含量的影響
隨處理濃度的增加,黃花草葉片中脯氨酸、可溶性蛋白和可溶性糖含量在第7、15天呈上升的趨勢(圖2)。處理第30天時,黃花草葉片中脯氨酸、可溶性糖含量隨錳濃度的增加呈下降趨勢,且相較第15天,可溶性糖含量的總體下降幅度大于脯氨酸,表明可溶性糖對錳脅迫更加敏感。可溶性蛋白含量在處理第30天依然呈隨濃度增加而上升的趨勢,但T3~T5處理明顯降低了其含量。綜上所述,黃花草幼苗在高濃度錳脅迫的前、中期可通過提高滲透調節物質來緩解毒害,但隨脅迫時間的增加,這一機制就會受到破壞,主要表現在T3~T5處理。
2.5 錳脅迫對黃花草幼苗MDA含量的影響
植物器官衰老或在逆境下遭受脅迫時往往發生膜脂過氧化作用,MDA是膜脂過氧化的最終分解產物,其含量的高低可反映植物細胞遭受逆境脅迫的程度。隨著錳濃度的增加,黃花草幼苗的MDA含量與對照相比呈先下降后升高的變化趨勢;隨著脅迫時間增加,高錳濃度處理下MDA含量均顯著升高(圖3)。T1處理下黃花草葉片MDA含量均低于對照組,但差異不顯著;T2~T5處理MDA含量隨錳濃度的增加和時間的延長上升幅度增大。在7、15和30 d時,T5處理下葉片MDA含量分別是對照組的3.1、3.5、3.6倍。綜上表明,T1處理并不會脅迫損傷黃花草幼苗,T2處理會隨時間延長逐漸表現出較小的脅迫效果,T3、T4、T5處理對黃花草幼苗表現出的脅迫效應會隨時間延長而加大。
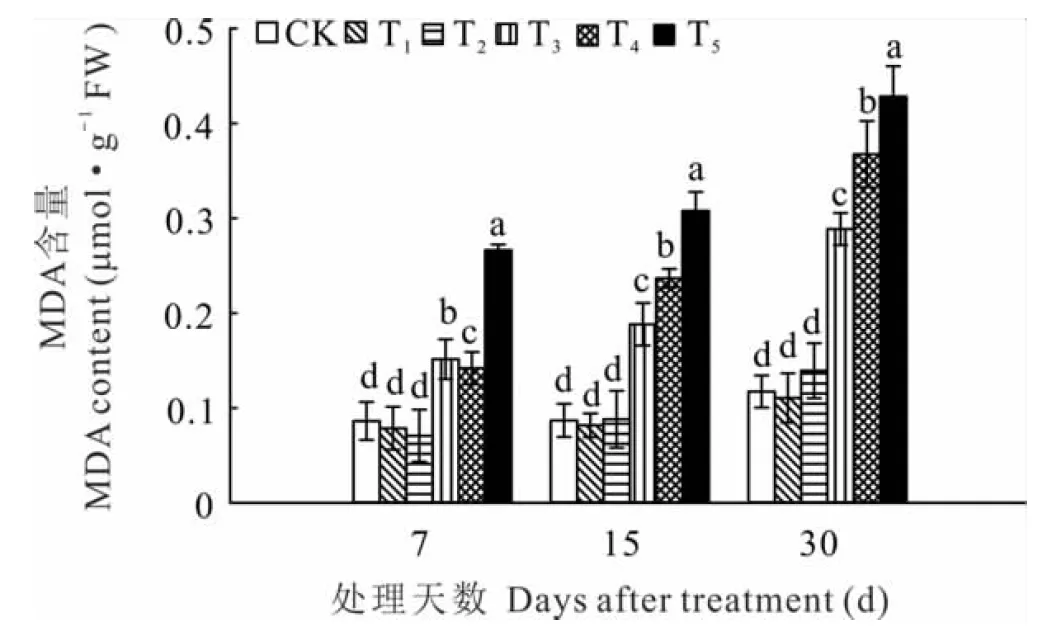
圖3 錳脅迫對黃花草幼苗MDA含量的影響
3 討論
3.1 錳脅迫對黃花草種子萌發及幼苗生長的影響
種子萌發與幼苗生長是植物對外界環境變化最敏感的階段,也是植物生活史中最脆弱的時期,能夠反映植物的適應能力[11]。植物能否在錳污染土壤上生存取決于其種子及幼苗在錳脅迫下是否可以萌發、生長,幼苗生長狀況直接影響植株以后的生長發育。本研究中,黃花草種子的發芽勢、發芽率、發芽指數和活力指數均隨錳濃度的增加而下降,說明錳脅迫對黃花草種子的萌發有抑制作用。這與文珂等[10]對錳脅迫下野大豆(Glycinesoja)以及徐雅梅等[12]對錳、鉛脅迫下垂穗披堿草(Elymusnutans)的研究結果一致。這表明錳脅迫對植物種子萌發有抑制作用是一種常見現象,原因可能是錳脅迫環境下種子內部儲存物質的分解及轉化受到抑制,進而影響種子萌發。
黃花草種子存在休眠現象,本研究前期試驗顯示采集種子后放置6個月,未見種子萌發。本研究是將該植物種子于室溫下放置一年后開展萌發試驗,但發芽率仍較低(50%左右)。采用500 mg·L-1赤霉素浸種處理后黃花草種子的發芽勢、發芽率、發芽指數和活力指數在0~5000 μmol·L-1錳處理時均顯著高于去離子水浸種處理,最高可達74.44%,說明赤霉素浸種對黃花草種子的萌發具有促進作用。當錳濃度達到15000~20000 μmol·L-1時,發芽率則無明顯差異,說明赤霉素浸種對高濃度錳處理下種子的萌發無促進作用。這與陳志飛等[13]對干旱脅迫下赤霉素對高羊茅(Festucaelata)種子萌發以及牛宋芳等[14]對鹽脅迫下赤霉素對紅砂(Reaumuriasoongorica)種子萌發的研究結果一致。這說明脅迫條件下赤霉素處理效果具有交互作用。其原因可能是赤霉素在高脅迫條件下加速了種子的新陳代謝活動,卻產生了較多的廢物,導致種子發芽情況未能顯著提高。這需要有更多的研究來驗證這一結果。
本研究顯示,在1000~5000 μmol·L-1錳處理下黃花草幼苗芽長、根長、干重和根冠比均高于對照且差異顯著;當錳濃度達到20000 μmol·L-1時,幼苗芽長、根長、干重和根冠比則均顯著低于對照,表明高濃度錳處理會對黃花草幼苗的生長產生顯著抑制作用,這種低促高抑的現象與王佳等[15]對錳脅迫下美洲商陸(Phytolaccaamericana)幼苗生長的研究結果一致。本研究顯示隨著錳濃度的增加,黃花草幼苗根受到的抑制作用比芽大,這與多數研究結果一致[7-16]。原因可能是因為根最先接觸錳,且是植物吸收和轉運重金屬的主要器官,導致受到的抑制程度大于芽。
3.2 錳脅迫對黃花草幼苗葉綠素含量的影響
葉綠素含量是衡量植物光合作用強弱的重要生理參數,間接體現植物對脅迫的適應能力[17]。在本研究中,低濃度(1000~5000 μmol·L-1)錳處理下黃花草幼苗葉片葉綠素a、葉綠素b、葉綠素a+b和葉綠素a/b均比對照組有所增加,而隨著錳濃度的增加,這些指標則逐漸下降并在較高濃度低于對照組。這與侯明等[18]對釩、鉻單一和復合脅迫下小麥(Triticumaestivum)幼苗葉片葉綠素含量及張雯莉等[19]對枸杞(Lyciumbarbarum)葉片在混合鹽脅迫下葉綠素含量的研究結果一致。這說明低濃度錳處理會促進葉綠素的合成,更有利于光合作用;而在錳處理濃度超出植物耐受范圍時,葉綠體受到損傷,葉綠素分解速度加快,光合能力減弱。其可能原因是錳既是組成葉綠體的必要元素,又直接參與光合放氧過程,所以低濃度錳處理下葉綠素含量增加,而高濃度錳處理下植物體內錳含量過高,受到毒害,抑制了葉綠體正常結構的合成及相關酶活性,葉綠素含量降低[1]。
此外,本研究還顯示,在脅迫第7天時,葉綠素a+b最大含量和葉綠素a/b最大值分別出現在5000 μmol·L-1處理和10000 μmol·L-1處理,脅迫至第30天時,則均出現在1000 μmol·L-1處理。這與燕江偉等[20]對干旱與鎘復合脅迫下互葉醉魚草(Buddlejaalternifolia)幼苗葉綠素含量的研究結果一致。一般認為植物葉綠素a/b的值越大,類囊體堆疊的程度越大,光能利用率越高[21]。隨著錳脅迫時間的延長,1000 μmol·L-1錳處理下的黃花草幼苗能保持葉綠素a+b含量和葉綠素a/b值高于對照,這說明該濃度處理能夠促進黃花草幼苗葉綠素的合成,維持自身正常的生長發育。
3.3 錳脅迫對黃花草幼苗抗氧化酶活性的影響
一般情況下,植物體內活性氧的產生和消除處于一種動態平衡的狀態。而當植物處于逆境中時,植物細胞會產生更多的活性氧自由基,造成膜脂過氧化,降低酶活及細胞滲透調節能力,對植物產生毒害。植物的抗氧化酶系統主要由SOD、POD和CAT協同構成,起著清除活性氧、降低脅迫所造成毒害的重要作用。本研究中,黃花草幼苗葉片中SOD活性在脅迫第7天隨錳脅迫濃度的增加而下降,但在整個試驗期內屬于較高水平;隨脅迫時間的延長SOD活性有所下降,20000 μmol·L-1處理下降最顯著。這與段才緒等[22]對鹽脅迫下決明(Cassiaobtusifolia)的研究結果一致。說明高濃度長時間的錳脅迫會降低SOD活性。在脅迫第7天,黃花草幼苗葉片POD和CAT呈現相同的變化趨勢,活性均隨錳濃度增加而快速增加,但隨脅迫時間延長,高濃度處理下的POD活性會下降,而CAT的活性變化不明顯。辛建攀等[23]研究表明隨著鎘濃度的增加和脅迫時間的延長,梭魚草(Pontederiacordata)葉片SOD和POD活性下降明顯,CAT活性無明顯變化。這與本研究結果一致。說明隨著錳濃度的增加和脅迫時間的延長,植物體內活性氧的產生已大于清除能力,加劇了植物細胞功能膜及酶系統的損傷。綜上,黃花草通過SOD、POD和CAT協同作用緩解氧化損傷,可忍受較高濃度錳脅迫。
3.4 錳脅迫對黃花草幼苗葉片脯氨酸、可溶性蛋白、可溶性糖含量的影響
植物在遭受各種逆境脅迫時,會迅速產生并積累脯氨酸、可溶性蛋白、可溶性糖等滲透調節物質,以保持細胞正常含水率及膨壓,對抗外界的不利環境[24]。脯氨酸是植物體內最常見且有效的小分子滲透調節物質;可溶性糖在參與滲透調節的同時可為植物生長和對抗不利環境因素提供能量;可溶性蛋白不僅參與植物體內大部分的代謝活動,還具有較強的親水性,可緩解因滲透脅迫所造成的細胞失水[7-8]。滲透調節物質含量在一定程度上反映了植物的抗逆性,含量越多表示抗性越強。但當脅迫超過植物的耐受范圍時,則會破壞這一機制,滲透調節物質的合成受到阻礙,含量減少[25]。本研究中,黃花草幼苗葉片中脯氨酸、可溶性蛋白和可溶性糖含量在脅迫第7、15天均快速積累增加,至第30天時急劇下降,濃度高的下降更為明顯。這與張虎等[26]對鈷脅迫下紫花苜蓿(Medicagosativa)的研究結果一致。說明黃花草幼苗通過提高脯氨酸、可溶性蛋白和可溶性糖含量緩解較高濃度錳脅迫帶來的毒害,但隨著脅迫時間的延長,滲透調節物質含量減少,平衡機制受到破壞,抗逆性降低。
3.5 錳脅迫對黃花草幼苗MDA含量的影響
逆境條件下,植物體內會產生大量活性氧,使細胞膜脂發生過氧化,細胞膜通透性增大。MDA作為細胞膜脂過氧化的產物,其含量高低代表植物遭受毒害的強度和對逆境條件下的反應能力。在本研究中,隨著錳濃度的增加,黃花草幼苗葉片MDA呈先降低后升高的變化趨勢,但隨著處理時間的延長,僅1000 μmol·L-1處理下的MDA含量低于對照組,說明該濃度處理不會對黃花草幼苗產生脅迫作用,與本研究中關于葉綠素含量的結果一致。高濃度錳處理下黃花草幼苗葉片MDA含量會隨濃度增加和脅迫時間的延長明顯增加,這與汪結明等[27]對4種藤本植物在錳污染土壤中的生理響應研究結果一致。說明高濃度錳脅迫導致黃花草幼苗細胞膜脂過氧化,植株所受毒害程度加大。
4 結論
錳脅迫會降低黃花草種子發芽勢、發芽率、發芽指數和活力指數。赤霉素浸種處理可增加黃花草種子發芽勢、發芽率、發芽指數和活力指數。隨著錳濃度的增加,黃花草幼苗芽長、根長、生物量和根冠比呈現“低促高抑”的現象。黃花草可通過提高SOD、POD和CAT活性及積累脯氨酸、可溶性糖和可溶性蛋白含量來緩解錳脅迫所造成的毒害。同時,本研究發現1000 μmol·L-1錳處理能夠促進黃花草的生長,表現為葉綠素含量的增加。但當錳濃度高于15000 μmol·L-1時,黃花草葉片MDA含量急劇增加,且抗氧化酶活性和滲透調節物質含量降低,說明黃花草生長受到抑制,且隨脅迫時間延長抑制作用更加明顯。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
