小麥幾丁質酶基因家族的全基因組鑒定及禾谷鐮刀菌脅迫下的表達分析
2019-11-21 02:52:28劉建豐袁祖麗
河南農業科學 2019年11期
徐 武,劉建豐,張 戈,徐 文,袁祖麗
(1.河南農業大學 生命科學學院,河南 鄭州 450002; 2.河南省農業科學院 農業經濟與信息研究所,河南 鄭州 450002; 3.河南省農業科學院 作物設計中心,河南 鄭州 450002)
幾丁質酶(EC 3.2.1.14)是廣泛存在于植物中的一種糖苷水解酶,能水解N-乙酰氨基葡萄糖聚合物[1]。幾丁質酶是一類與疾病相關的蛋白質,可以破壞病原菌的細胞壁和昆蟲的外骨骼,在植物防御生物和非生物脅迫過程中發揮著重要作用[2]。根據氨基酸序列相似性,可將植物幾丁質酶分為糖苷水解酶亞家族18(GH-18)和糖苷水解酶亞家族19(GH-19)[3]。根據CAZy數據庫(http://www.cazy.org/Glycoside-Hydrolases.html),植物幾丁質酶被分為5個不同的類(Ⅰ—Ⅴ)[4]。其中,Ⅰ類、Ⅱ類和Ⅳ類屬于GH-19亞家族,Ⅲ類和Ⅴ類屬于GH-18亞家族[3]。Ⅰ類幾丁質酶成員具有N-末端甲殼素結合區和GH-19催化結構域;Ⅱ類幾丁質酶成員只有1個催化結構域,與Ⅰ類幾丁質酶成員具有較高的序列和結構相似性,但缺乏甲殼素結合區;Ⅳ類幾丁質酶成員與Ⅰ類幾丁質酶成員具有較高的同源性,但由于甲殼素結合區和催化結構域的缺失,其同源性較小[5];Ⅲ類和Ⅴ類幾丁質酶成員均具有GH-18催化結構域和保守序列DXDXE,但與其他類成員無氨基酸序列同源性[6]。
在高等植物生長發育過程中,幾丁質酶基因在植物抗病過程中起到重要作用[7-16]。1998年,TABEI等[7]通過農桿菌介導的方法將水稻的幾丁質酶基因RCC2導入黃瓜,與非轉基因植株相比,轉基因黃瓜對灰霉病的抗性顯著提高。2000年,YAMAMOTO等[8]將1個水稻Ⅰ類幾丁質酶基因導入葡萄,顯著提高了轉基因葡萄對白粉病的抗性。近年來,已將幾丁質酶基因轉入水稻[9-10]、棉花[11]、大豆[12-13]、花生[14]、煙草[15-16]等高等植物中,也明顯提高了轉基因植株對各類病菌的抗性,這些研究說明幾丁質酶基因在植物抗病應答過程中具有重要的作用。
小麥作為世界上重要的糧食作物之一,其生長發育過程中受到許多病菌的危害,尤其是禾谷鐮刀菌侵染引起的赤霉病。近年來,赤霉病成為造成小麥嚴重減產的病害之一,不僅降低小麥產量,而且還產生赤霉毒素影響品質,甚至危害人類健康。自小麥參考基因組序列公布以來,小麥基因功能的研究報道越來越多,但是小麥中幾丁質酶家族成員全基因組鑒定的研究尚未見報道。研究幾丁質酶基因對提高小麥赤霉病抗性具有重要意義。為此,對小麥幾丁質酶基因家族成員進行鑒定,并對其基因結構、保守基序(Motif)、染色體分布、系統進化及禾谷鐮刀菌脅迫下的表達情況進行分析,為進一步研究幾丁質酶基因在小麥生物脅迫應答方面的作用及小麥抗病分子育種中的應用奠定基礎。
1 材料和方法
1.1 數據來源
小麥基因組參考序列及其注釋文件均下載自Ensembl Plants數據庫(ftp://ftp.ensemblgenomes.org/pub/plants/release-41)。
1.2 小麥幾丁質酶基因家族成員的鑒定
小麥幾丁質酶基因家族成員結構域信息隱馬爾科夫模型(PF00704和PF00182)均下載自Pfam數據庫(http://pfam.xfam.org/)。利用HMMER 3.1軟件結合幾丁質酶基因家族成員結構信息隱馬爾科夫模型搜索小麥基因組注釋的蛋白質序列文件,E值設置為 1E-10。如果同一個基因對應的多個注釋蛋白質序列被鑒定到,選取長度最長的蛋白質序列。采用Compute pI/Mw tool(https://web.expasy.org/compute_pi/)預測蛋白質的等電點和分子質量。
1.3 小麥幾丁質酶基因家族基因結構和保守基序分析
根據鑒定到的小麥幾丁質酶家族成員蛋白質序列編號和Ensembl Plants中小麥基因組注釋信息文件,提取小麥幾丁質酶基因家族基因的注釋信息,然后將其提交GSDS網站(http://gsds.cbi.pku.edu.cn)來繪制基因結構圖。用MEME軟件(http://meme-suite.org/tools/meme)來預測幾丁質酶家族成員的保守基序。
1.4 小麥幾丁質酶家族成員的系統進化分析
將鑒定到的小麥幾丁質酶家族成員蛋白質序列利用MEGA 7.0軟件包中的MUSCLE軟件進行多重序列比對,然后進行系統進化樹構建。系統進化樹采用NJ(Neighbor-joining)法構建,抽樣次數為1 000[17]。
1.5 小麥幾丁質酶基因家族基因的表達分析
小麥RNA-Seq測序數據來自SRA數據庫[18],登錄號為SRR7000820—SRR7000822、SRR7000826—SRR7000828、SRR7000832—SRR7000834、SRR7000838—SRR7000840、SRR7000844—SRR7000846、SRR7000850—SRR7000852、SRR7000856—SRR7000858、SRR7000862—SRR7000864。采用TopHat 2[19]軟件將其與參考基因組序列比對,運用HTSeq軟件[20]統計每個轉錄本的reads數,然后用DESeq2 R包[21]進行差異分析,保留log2(變化倍數)大于1的轉錄本進行熱圖制作。
2 結果與分析
2.1 小麥幾丁質酶基因家族成員的鑒定及序列特征分析
在小麥基因組中共鑒定到159個幾丁質酶基因家族成員,如表1所示,這些幾丁質酶基因主要分成2個亞家族,即GH-18和GH-19家族,分別有82個和77個基因。小麥幾丁質酶基因編碼蛋白質序列長度為157~739個氨基酸,分子質量為16 648.96~79 774.66 u,理論等電點介于4.10~10.25。對小麥幾丁質酶基因家族成員進行染色體定位分析發現,本研究鑒定到的159個小麥幾丁質酶基因家族中的154個基因不均勻分布在21條染色體上,5個定位在尚未組裝完整的scaffold上。3號染色體上幾丁質酶基因數目最多,共49個,其中3A、3B、3D染色體上分別有18、16、15個;2號染色體次之,含有31個,2A、2B、2D染色體上分別有12、8、11個;7A、7B、7D染色體上分別有8、12、7個; 1A、1B、1D染色體上分別有9、8、6個;5A、5B、5D染色體上分別有7、4、4個;4A、4B、4D染色體上分別有1、1、3個;6號染色體上數目最少,共4個,6A、6B、6D染色體上分別有2、1、1個。從染色體組的角度來看,同一編號染色體A、B、D 3套染色體組含有的幾丁質酶基因數目差別不大,表明幾丁質酶基因在A、B、D 3套染色體組進化過程中沒有發生明顯的基因丟失現象。

表1 小麥幾丁質酶基因家族成員信息
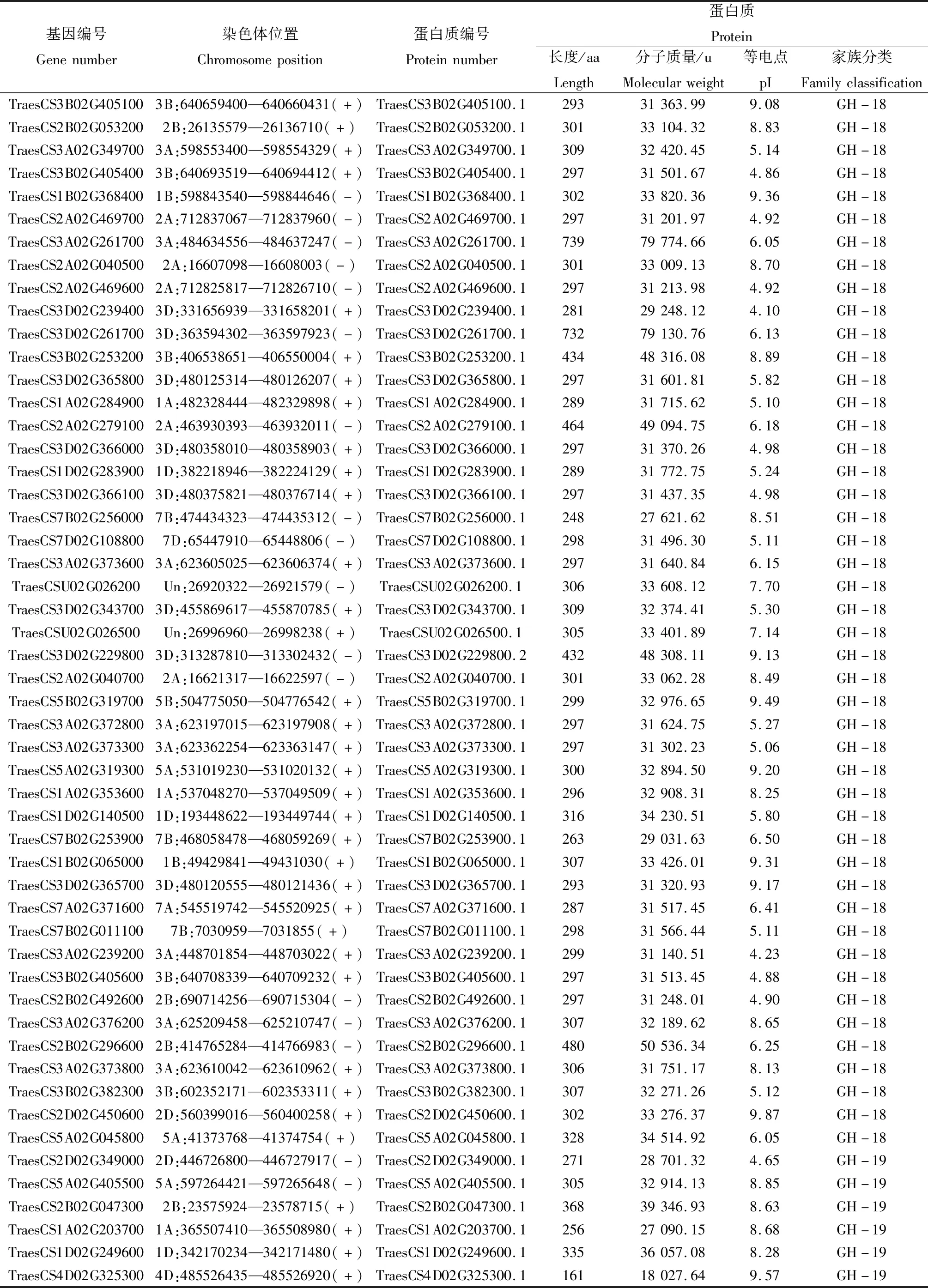
續表1 小麥幾丁質酶基因家族成員信息

續表1 小麥幾丁質酶基因家族成員信息

續表1 小麥幾丁質酶基因家族成員信息
2.2 小麥幾丁質酶家族成員的分類
利用本研究鑒定到的小麥幾丁質酶家族成員蛋白質序列構建系統進化樹,并與已知擬南芥幾丁質酶家族成員蛋白質序列進行比對,然后根據已知擬南芥幾丁質酶家族成員分類信息對小麥幾丁質酶家族基因成員進行分類,結果(圖1)表明,小麥幾丁質酶家族成員分為5類,GH-19和GH-18亞家族分別聚在2個不同的分支。GH-18亞家族包括Ⅲ類和Ⅴ類,其中,Ⅱ類含有34個成員,Ⅴ類含有48個成員。GH-19亞家族中包括Ⅰ、Ⅱ、Ⅳ類,其中,Ⅰ類含有52個成員,Ⅱ類含有13個成員,Ⅳ類含有12個成員。

圖1 小麥幾丁質酶家族成員的系統進化樹Fig.1 The phylogenetic tree of chitinase family members in wheat
2.3 小麥幾丁質酶基因家族成員的基因結構和保守基序分析
為了進一步研究小麥幾丁質酶基因家族成員的進化關系,根據小麥基因組參考序列的功能注釋信息并繪制其外顯子-內含子結構圖(圖2—3),結果表明,GH-18亞家族基因含有1~8個外顯子,其中,大部分只有1個外顯子,不含有內含子;少量含有1~2個內含子;TraesCS3D02G229800.2、TraesCS3A02G223700.1和TraesCS3B02G253200.1三個基因含有7~8個內含子。GH-19亞家族基因含有1~5個外顯子,大部分成員含有1~2個內含子,基因長度差別不大,其中,TraesCS3A02G415700.1基因內含子長度比較長;TraesCS3D02G229800.2、TraesCS3A02G223700.1、TraesCS3B02G253200 .1和TraesCS3A02G415700.1四個基因長度比其他幾丁質酶基因長很多,而且內含子長度比外顯子長度長,說明在進化過程中這4個基因內部可能發生了大片段的插入。對GH-18和GH-19亞家族成員中的保守基序進行分析(圖4—5)發現,GH-18亞家族82個成員中有79個包含保守基序5,67個成員中包含保守基序1、保守基序2、保守基序3、保守基序4;GH-19亞家族77個成員中有74個包含保守基序1、保守基序4,其中,GH-19亞家族的第Ⅳ亞類不含有保守基序2,剩下65個成員中62個含有此基序。綜上所述,相同亞家族的幾丁質酶基因幾乎具有相似的外顯子-內含子結構和保守基序。

圖2 小麥GH-18亞家族成員基因結構分析

圖3 小麥GH-19亞家族成員基因結構分析

圖4 小麥GH-18亞家族成員保守基序分析
2.4 小麥幾丁質酶基因家族成員的表達分析
通過分析4個小麥品系的RNA-Seq數據,得到幾丁質酶基因家族成員在小麥接種禾谷鐮刀菌前后的表達譜。由圖6可知,雖然幾丁質酶基因在不同小麥品系中被誘導表達的程度存在差異,但是在4個小麥品系被禾谷鐮刀菌侵染后,一些基因(TraesCS1A02G203600.1、TraesCS1D02G249600.1、TraesCS3A02G260100.1、TraesCS6D02G199500.1、TraesCS6A02G216100.1、TraesCS2D02G349400.1、TraesCS2A02G350900.1、TraesCS2A02G350800.1、TraesCSU02G026200.1、TraesCS7B02G256000.1、TraesCS1B02G158000.1、TraesCS1D02G140500.1、TraesCS1A02G141500.1、TraesCS3A02G373800.1、TraesCS3B02G406000.1)的表達均明顯上調,說明幾丁質酶基因在小麥應答禾谷鐮刀菌過程中起重要的作用。

圖5 小麥GH-19亞家族成員保守基序分析
3 結論與討論
本研究從小麥基因組中共鑒定到159個幾丁質酶基因,分為GH-18和GH-19兩個亞家族,相同亞家族成員幾乎具有相似的外顯子-內含子基因結構和保守基序,并且聚類在同一分支上。此外,在A、B、D 3套染色體組中鑒定到的幾丁質酶基因家族基因數目差別不大,說明幾丁質酶基因在A、B、D 3套染色體組長期進化中沒有發生明顯的基因丟失現象。
近年來,植物幾丁質酶基因在很多農作物抗逆抗病過程中具有重要的作用[9-16]。本研究分析了幾丁質酶基因家族成員在3個小麥抗病品系和1個小麥感病品系被禾谷鐮刀菌侵染前后的表達變化,結果表明,一些幾丁質酶基因家族成員的表達明顯被禾谷鐮刀菌誘導,而且同一家族的不同基因成員的表達量也存在明顯的差異。這種現象可能由于基因家族在擴張的過程中同一家族不同基因成員負責不同生物或者非生物脅迫應答功能不同所致。
