煎炸花生油中的總極性組分對脂質代謝的影響
2019-11-20 08:30:56鞠婧捷蘇青峰李有棟李進偉曹培讓劉元法
中國油脂 2019年10期
鞠婧捷,蘇青峰,李有棟,李進偉,曹培讓,劉元法
(1.江南大學 食品學院,江蘇 無錫 214122; 2.重慶紅蜻蜓油脂有限責任公司,重慶 400010)
油脂深度煎炸是最常見的食品烹飪方法之一。食用油在煎炸過程中發生一系列的化學反應,導致煎炸油的理化指標發生變化,產生氧化甘油三酯、聚合甘油三酯和游離脂肪酸等極性大于甘油三酯的組分,這些極性組分是熱氧化油脂劣變最主要的成分[1]。同時,極性組分含量也是衡量油脂質量的主要指標之一[2]。現有關煎炸油的文獻大多圍繞在穩定性好的飽和油脂體系,如棕櫚油[3-4],而對不飽和油脂體系——花生油的相關研究較少。越來越多的人將花生油作為主要的食用油脂,尤其是我國的北方地區,食品攤位甚至某些成熟的食品店、飯店為了降低費用會使用反復煎炸的花生油,煎炸花生油的使用現狀遠比我們想象的要嚴峻。因此,研究煎炸花生油對健康的影響具有一定的現實意義。
目前,煎炸油和極性組分的研究主要集中在開展動物實驗,如Li等[5]發現極性組分對昆明種小鼠的脂質代謝有不利的影響,穆昭[6]研究表明極性組分會造成小鼠的骨髓多染紅細胞的微核率增加,同時引起脂質代謝紊亂。然而,目前有關極性組分對細胞影響的研究,尤其是脂質代謝方面的研究相對較少。脂質代謝是研究非酒精性脂肪肝病(NAFLD)的重要前提條件,正如“雙重打擊”假說所解釋的,NAFLD發生的病理機制第一重“打擊”主要歸因于脂質代謝紊亂[7]。脂質代謝紊亂是指脂質合成和輸出的不平衡,可能會促使甘油三酯在肝細胞內積累[8-9]。脂質代謝基因發生異常是導致肝細胞病變的關鍵控制點。固醇元件結合蛋白-1c(SREBP-1c)驅動肝臟脂肪生成,是一種控制膽固醇、脂肪酸、甘油三酯和磷脂合成所需的堿性螺旋-環-螺旋轉錄因子[10]。SREBP-1c通過激活脂質合成所需的基因脂肪酸合成酶(FAS)和乙酰輔酶A羧化酶(ACC)優先調節從頭脂肪生成,而SREBP-1c通過激活膽固醇合成所需的基因3-羥基-3-甲基戊二酰輔酶A還原酶(HMGCR)來控制膽固醇體內平衡[11]。HepG2細胞來源于肝細胞癌癥患者的肝臟,保留了肝實質細胞的許多形態特征,其特定酶的表達和活性與肝細胞相同,同時HepG2細胞比其他細胞系更敏感[8, 12]。因此,本研究使用煎炸花生油中提取的極性組分,通過HepG2細胞構建脂質代謝的細胞模型,研究極性組分對HepG2細胞的脂質沉積和脂質代謝的相關影響。
1 材料與方法
1.1 實驗材料
1.1.1 原料與試劑
花生油(不含抗氧化劑),無錫德合食品科技有限公司;雞腿,無錫歐尚超市。乙醚、石油醚(沸程30~60℃)、正己烷、甲醇,異丙醇均為分析純;硅膠(200~300目),青島海洋化工有限公司;HepG2細胞,中國上海科學院細胞研究所;油紅O,美國索萊寶生物科技有限公司;胰酶消化液,美國Gibco公司;MEM細胞培養基、UNIQ-10柱式總RNA抽提試劑盒、AMV第一鏈cDNA合成試劑盒、2×SG Fast Qpcr Master Mix試劑盒,上海生工股份有限公司。
1.1.2 儀器與設備
EF-818L單缸單篩電炸鍋,江蘇秉宏生物科技有限公司;Axio Vert A1倒置熒光顯微鏡,德國蔡司公司;CFX96實時熒光定量基因擴增儀,美國伯樂公司。
1.2 實驗方法
1.2.1 煎炸花生油的制備
將7 L花生油置于電炸鍋中并保持溫度在(180±2)℃,油炸過程中未補充新鮮油。煎炸實驗每天開展10 h,連續4 d,每隔1 h換500 g雞腿進行煎炸,第4天結束時收集煎炸花生油樣品,在-20℃避光保存。
1.2.2 極性組分的制備
根據GB 5009.202—2016《食品安全國家標準 食用油中極性組分(PC)的測定》第二法柱層析法制備極性組分,并在此基礎上進行改良。先用50 g硅膠裝柱,再用洗脫劑A(石油醚-乙醚,體積比87∶13)洗滌硅膠柱。將溶于洗脫劑A的5 g花生油樣品倒入柱中同時用洗脫劑A洗脫花生油樣品直至非極性組分洗脫完畢,再用750 mL洗脫劑B(乙醚)和250 mL洗脫劑C(甲醇)收集極性化合物,洗脫速度15 mL/min。最后,通過旋轉蒸發(40℃)和真空干燥(40℃,0.01 MPa)收集溶解在洗脫劑B和洗脫劑C中的極性化合物。為了更有效地分離極性化合物,使用薄層色譜進行實時監測。
1.2.3 HepG2細胞培養
HepG2細胞均勻分散在含有10%胎牛血清(FBS)、100 U/mL青霉素和100 μg/mL鏈霉素的MEM細胞培養基的完全培養基中,置于37℃的CO2培養箱中培養。待HepG2細胞融合度達到80%左右,吸除原培養液,用1 mL的磷酸鹽緩沖溶液(PBS,pH=7.4)清洗2遍,加入1 mL胰酶消化液,放入CO2培養箱中,37℃消化1 min,加2 mL完全培養液終止消化,輕緩地用吸管反復吹打細胞,再全部轉移至另一個15 mL離心管中,800 r/min離心5 min,吸除培養液,再加入3 mL完全培養液重新懸浮細胞。細胞計數后取相應數量細胞進行培養。
1.2.4 HepG2細胞的處理
采用細胞計數板計數20×104個HepG2細胞2 mL 并將其接種于六孔板中,培養1 d后細胞完全貼壁,加入質量濃度分別為0、0.1、0.5、1.0、2.0 mg/mL的極性組分2 mL,對照組不加入極性組分,用無血清的培養基代替,孵育24 h后,用PBS清洗獲得極性組分處理和對照組的HepG2細胞。
1.2.5 油紅染色
通過油紅染色評估HepG2細胞內脂質堆積的情況。將1.2.4獲得的HepG2細胞,用10%甲醛固定30 min,然后油紅O染料進行染色,PBS洗掉染料后,采用倒置熒光顯微鏡拍照觀察。
1.2.6 實時定量PCR(RT-PCR)
將1.2.4獲得的HepG2細胞,使用UNIQ-10柱式總RNA抽提試劑盒提取RNA、AMV第一鏈cDNA合成試劑盒進行反轉錄和2×SG Fast Qpcr Master Mix試劑盒進行PCR擴增。引物通過Primerbank設計,經上海生工科技股份有限公司合成。引物信息如表1所示。內參基因為β-actin,將每個基因的表達水平進行平均計算,用2-ΔΔCt方法[13]計算不同基因相對于對照基因的相對轉錄水平的倍數變化。
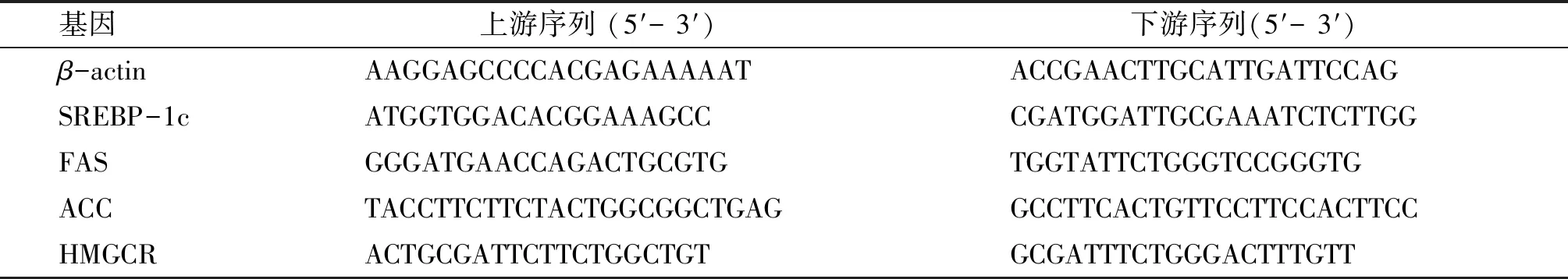
表1 RT-PCR的引物序列
1.2.7 統計學方法
采用SPSS7.0統計軟件進行Duncan’s檢驗和Anova分析,數據用“平均值±標準差”表示,P<0.05表示差異具有統計學意義。
2 結果與分析
2.1 極性組分對HepG2細胞脂質沉積的影響
圖1是用不同質量濃度的極性組分處理HepG2細胞并油紅染色的40×顯微鏡的放大圖。

注:A.0 mg/mL; B.0.1 mg/mL; C.0.5 mg/mL; D.1.0 mg/mL; E.2.0 mg/mL。
圖1 不同質量濃度的極性組分對HepG2細胞脂質沉積的影響
由圖1可見,當不加入極性組分時(圖1A),細胞界限較為明顯,細胞漿充足,含有少許紅色圓形結構可能是細胞自身存在的脂質,幾乎沒有油紅O的附著,說明不加入極性組分的HepG2細胞產生的脂質很少。隨著極性組分含量逐漸增加,脂質沉積情況逐漸加重。當極性組分的質量濃度為0.1 mg/mL時(圖1B),脂質沉積與對照組存在顯著不同,但是與極性組分質量濃度為0.5 mg/mL時(圖1C)差異不大,紅色小泡狀的脂滴主要附著在細胞的邊緣,細胞并沒有大部分被聚積的脂質占滿,細胞膜的界限開始不明顯,體現了小泡狀脂肪生成特征。然而,當極性組分的質量濃度增加到1.0 mg/mL時,被油紅O附著的區域進一步擴大,以不均勻的形態分布在胞漿中。當HepG2細胞用2.0 mg/mL的極性組分處理時,不僅是脂質聚積的現象變得更加明顯,而且分不清細胞膜的位置,細胞的形態也發生了很大的改變,說明當極性組分的質量濃度達到一定程度時,不僅僅會造成脂質積累還會導致細胞形態的改變,并且造成一定程度的細胞變性。目前鮮有文章報道花生油的極性組分對細胞脂質沉積情況的影響。因此,我們以梯度劑量處理HepG2細胞的方式,觀察極性組分對脂質沉積的作用,與極性組分應用到老鼠中的研究結果一致[5],極性組分不僅會引起老鼠肝臟脂質堆積,也會造成HepG2細胞脂質沉積,這是引發肝臟等相關疾病的第一重關鍵因素,為證明極性組分引起NAFLD等疾病提供了強有力的證據。
2.2 HepG2細胞的脂質代謝基因表達
研究表明,FAS、ACC和HMGCR是SREBP-1c的下游靶基因[14]。其中,FAS催化乙酰輔酶A和丙二酸單酰輔酶A合成軟脂酸,ACC催化乙酰輔酶A生成丙二酰輔酶A,都是控制脂質合成的關鍵限速酶。這4種脂質合成基因對于調控細胞的脂質代謝有很大的影響,為了檢測極性組分對HepG2細胞調控脂質代謝相關基因表達的影響,測定了這4種基因的mRNA水平,結果見圖2。

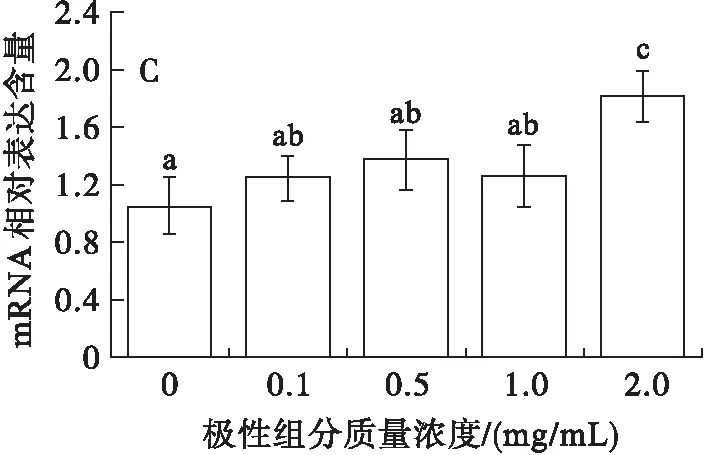
由圖2可見,與對照組比較,SREBP-1c和FAS的mRNA的表達均隨極性組分質量濃度的增加出現顯著性差異(P<0.05)(圖2A和圖2B)。盡管ACC的mRNA的表達在極性組分質量濃度低于1.0 mg/mL時與對照組差異不顯著,但是當極性組分質量濃度進一步增加時,同樣出現了顯著性差異(P<0.05)(圖2C)。一定程度上證明了SREBP-1c對其下游基因FAS和ACC起到了上調作用。由圖2D可知,不同質量濃度的極性組分對HMGCR的mRNA的表達水平沒有影響,證明了盡管HMGCR基因同樣是SREBP-1c的下游靶基因,但是其對HepG2細胞的脂質代謝沒有起到調控作用。由此可以認為極性組分在HepG2細胞中通過上調SREBP-1c、FAS、ACC途徑引發脂質代謝紊亂。
3 結 論
研究界普遍認為極性組分對健康存在潛在危害,但是造成危害的細胞學的現象和原因卻不清楚。本文研究表明:在病理現象方面,極性組分導致HepG2細胞內甘油三酯的積累;在分子水平上,極性組分上調HepG2細胞的SREBP-1c、FAS、ACC脂質合成基因途徑,引起脂質合成速率和游離脂肪酸水平的增加,從而導致了脂肪酸合成和分解的不平衡。本研究是首次解釋不飽和體系的植物油——花生油產生的極性組分可以導致脂質代謝紊亂,可以用作預防脂質代謝紊亂相關疾病(如NAFLD)的新靶點,并且為研究煎炸油對肝臟疾病的影響提供了新的切入點。
