北京油松人工林競爭指數的適用性評價
2019-11-13 02:54:44裴順祥張連金辛學兵
浙江農林大學學報 2019年6期
李 超,裴順祥,張連金,郭 嘉,辛學兵
(中國林業科學研究院 華北林業實驗中心,北京102300)
林木競爭指生長在同一生境中的林木為爭奪共需的有限資源而阻礙其他林木正常生長發育,甚至致使其死亡的行為[1-3]。林木競爭導致個體間差異,進而動態調節林分結構及其空間關系,決定林木競爭格局和空間生態位[4],從而推動群落演替。林木競爭研究能為調節林分結構和保護瀕危物種提供依據,有利于維持生物多樣性和生態系統的穩定性[1,5-6],一直是生態學研究的核心內容之一。競爭指數的提出為定量分析林木競爭強度提供了途徑,根據其與距離的關系分為完備型和非完備型競爭指數[7]。非完備型競爭指數主要有樹高斷面積競爭指數、樹冠表面積外露競爭指數等[8],僅與對象木信息或周圍環境信息有關;完備型競爭指數充分結合對象木信息與周圍環境信息,主要有基于樹冠面積重疊的Bella競爭指數[9]、Hegyi競爭指數[10]和基于交角的競爭指數[11]等。其中,Hegyi競爭指數及其改進模型因簡單易算,適應性較強而被廣泛運用[1,3,12-15],但該指數卻未考慮冠幅和樹高在競爭中的影響;而樹冠面積重疊競爭指數和基于交角的競爭指數則分別涉及了冠幅與樹高的影響,亦被廣泛運用[16-18]。油松Pinus tabuliformis是中國北方重要造林樹種[19],第八次北京市園林綠化資源普查成果報告顯示,按優勢樹種劃分,北京油松林面積和蓄積量分別約8.45萬hm2和213.19萬m3,占北京森林面積與蓄積量的16.76%和15.16%,是北京森林生態系統的重要組成部分。因經營管理不到位,油松林單位面積蓄積量僅為25.22 m3·hm-2,低于北京森林平均水平(27.88 m3·hm-2);林分現存結構不合理,林木競爭激烈,天然更新困難,林分質量、生物多樣性和穩定性均較低[20],亟需開展科學的經營管理。為改善油松人工林結構,首先應篩選一種更適合油松的林木競爭模型來了解該區域油松人工林林內競爭機制,為后期結構調整提供基礎數據和理論支撐。如何在眾多模型中選用最適模型來量化林分林木競爭強度一直是研究的重點[11,21]。目前油松林競爭研究多為競爭模型的構建及改進、種內與種間競爭關系分析、林分結構與生物多樣性探討等[14,19-20,22],鮮有不同類型競爭模型對油松的適用性評價。基于此,本研究以京西九龍山油松人工林為對象,采用回歸分析及k折交叉驗證法[23],開展不同類型競爭指數模型對油松的適用性評價研究,篩選出更適于油松人工林的競爭模型,試圖初步揭示其競爭機制,為后期經營管理提供依據。
1 研究區概況
研究區位于北京市門頭溝九龍山(39°54′~39°59′N, 115°59′~116°07′E), 該區屬暖溫帶大陸東岸半濕潤大陸性季風氣候,年平均氣溫為11.8℃,年平均降水量為623 mm,主要集中在6-9月,年平均蒸發量為1870 mm,無霜期為216 d。土壤屬于山地褐土,土層普遍較薄,含石量高[24]。研究區于20世紀60年代開始封山育林,同期選用2~3 a油松幼苗進行大面積的魚鱗坑整地造林,油松人工林初始密度為5000株·hm-2。在海拔100~997 m分布著典型落葉闊葉林和溫性針葉林[24],主要樹種有油松,栓皮櫟Quercus variabilis,側柏Platycladus orientalis,白蠟樹Fraxinus chinensis,榆樹Ulmus pumila,北京丁香Syringa pekinensis,樸樹Celtis sinensis,臭椿Ailanthus altissima,杜梨Pyrus betulifolia和桑Morus alba等。
2 研究方法
2.1 樣地調查
于2017年6月, 在研究區內設置 1 個 100 m × 100 m 油松人工林樣地(39°55′08″N, 116°01′18″E)。該樣地平均海拔750 m,坡度18°,北坡,林齡約53 a。對樣地內胸徑大于5 cm的林木進行每木檢尺,記錄樹種名稱,用圍徑尺測量胸徑、用皮尺測量冠幅、用激光測高儀測量樹高和枝下高,并用TOPCON全站儀進行定位。樣地基本概況見表1,林木冠幅及位置分布見圖1。

表1 林分調查因子匯總Table 1 Stand survey factors

圖1 樣地林木冠幅位置分布Figure 1 Crown width and position distribution of the trees in plots
2.2 數據分析方法
2.2.1 對象木和競爭木的確定 采用8鄰域大樣地平移法進行邊緣校正[15],將原樣地內所有油松作為對象木,并與其4株最近相鄰木構成最佳空間結構單元[8]。
2.2.2 生物量模型及競爭模型 采用裴雅茹[25]的北京油松生物量研究模型對樣地內油松生物量進行估算,公式為:

式(1)中:W為生物量(kg),D為林木胸徑(cm),H為樹高(m)。以Dj為競爭木j的胸徑(cm),Di為對象木i的胸徑(cm),dij為對象木i與競爭木j之間的距離(m),n為競爭木數量,得到Hegyi競爭指數[10]:

式(2)中:CHg_i為 Hegyi競爭指數。 Bella 競爭指數(CIOi)[9]公式為:

式(3)中:Co_i為Bella競爭指數,ZO_ij為對象木i與競爭木j樹冠投影重疊面積(m2),ZA_i為對象木i的樹冠投影面積(m2)。基于交角的林木競爭指數(d_α_CIi)公式為:

式(4)~(6)中:Cd_α_i[17]為總競爭指數,Cd_α_i上和Cd_α_i側則分別為上方遮蓋和側方擠占; α1和 α2為對象木與競爭木之間的高度交角(°), 如果相鄰木j高于對象木i(圖 2A),α1=arctg(Hi/dij),α2=arctg[(Hj-Hi)/dij],cij=1; 否則(圖 2B)α1=arctg(Hj/dij), α2不存在,cij=0。 對象木的各競爭木(n=4)競爭強度的均值為競爭指數,在競爭指數計算公式(2),(3),(5),(6)中,競爭木j的競爭強度可描述為:
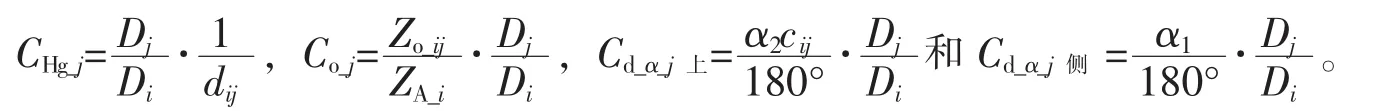
2.2.3 模型適用性評價及油松人工林競爭現狀分析 本研究的油松人工林為同齡林,油松生物量可較好反映油松所受競爭強度。為篩選更適合油松人工林的競爭指數,分別采用指數、對數、多項式、線性和冪函數等模型作油松生物量與該3類競爭指數的回歸分析,獲取決定系數(R2)最大的最優模型,線性轉換后,進行n次k折交叉驗證(n和k均為10),平均n次k折交叉驗證的R2作為最終的驗證R2。k折交叉驗證,即將樣本分為分割成k個子樣本,輪流將(k-1)個子樣本組合作為訓練集,另外的1個樣本作為驗證模型的保留集,k次的平均交叉驗證識別正確率即為模型檢驗結果[23]。k折交叉驗證的驗證R2較回歸R2損失得越少,模型泛化能力越強,以此結合決定系數(R2)來篩選競爭指數,并用該競爭指數對林分現狀作進一步分析。

圖2 上方遮蓋與側方擠占示意Figure 2 Schematic diagram of the over shading and the lateral extruding
2.2.4 數據分析 采用SVMS 1.0軟件對采集的油松人工林數據進行預處理,獲取對象木與4株競爭木所構競爭單元基本信息,通過Excel 2010完成各對象木生物量和競爭指數的計算,并運用R軟件編程獲取生物量與競爭指數間的最優模型、決定系數(R2)及n次k折交叉驗證R2,在SPSS 20.0軟件中完成對象木徑級和競爭指數的相關分析。
3 結果與分析
3.1 油松對象木與競爭木概況
樣地內胸徑5 cm以上林木共1159株13種,其中油松對象木1054株。油松對象木徑級分布為正態分布(偏度0.093,峰度-0.197;P=0.373>0.05),正態分布曲線略正偏。油松對象木及競爭木概況如表2所示。油松對象木和競爭木胸徑主要集中在10~25 cm,分別占88.24%和85.08%,其中5~10 cm徑級的油松對象木數量較少,僅占8.16%。除大于等于30 cm的徑級外,同徑級對象木平均胸徑大于競爭木,但同徑級對象木平均樹高皆小于等于競爭木。

表2 油松對象木及競爭木的概況Table 2 The status of objective tree of Pinus tabuliformis and competitor
3.2 油松人工林不同競爭指數的適用性分析
林木生物量累積速率受競爭強度影響,所受競爭強度越小,其生物量累積速率越快[18]。
采用回歸分析法對油松生物量與其競爭指數進行分析,結果表明(表3):Hegyi競爭指數和d_α_CIi的最優模型為冪函數,CIOi的最優模型則為指數函數,且均通過模型的顯著性檢驗(P<0.01)。10次10折交叉驗證結果顯示:各模型的交叉驗證R2較決定系數R2損失均小于0.005;其中,d_α_CIi與生物量決定系數R2最高,R2損失量最小,分別為0.7705和0.0010,Hegyi競爭指數次之,CIOi最差。各競爭指數與生物量的關系如圖3所示,油松生物量均隨各競爭指數的增大而減小;其中,CIOi和Hegyi競爭指數較d_α_CIi存在較多的離群點。綜上所述,d_α_CIi能更好反映油松所受競爭強度。

表3 不同競爭指數下的對象木生物量響應模型及交叉驗證結果Table 3 Biomass response model of objective tree and cross-validation results under different competition indices

圖3 對象木生物量與競爭指數的相關性分析Figure 3 Correlation analysis of different competition indices with biomass
3.3 油松對象木的競爭強度分析
3.3.1 競爭強度隨油松對象木胸徑的變化 從圖4可知:小徑級油松受側方擠占和上方遮蓋比例相差不大,隨著徑級的增大,側方擠占逐漸主導競爭,大徑級油松幾乎不受上方遮蓋影響。油松所受平均競爭強度隨徑級增大而減少,上方遮蓋隨徑級的增大而迅速減小。除5~10 cm徑級外,油松種間平均競爭強度均小于種內。對油松對象木的競爭指數作徑級分析,因其徑級競爭指數未滿足方差齊性檢驗,故選用非參數檢驗進行分析。非參數檢驗的Kruskal-Wallis檢驗結果表明油松徑級間競爭指數存在顯著差異(P<0.05),由圖5可知:油松胸徑>20 cm,各徑級間差異均不顯著,具體表現為20~25 cm與25~30 cm,20~25 cm與≥30 cm及25~30 cm與≥30 cm徑級間差異不顯著,其余各徑級間差異均顯著(調整后P<0.05)。這表明油松胸徑<25 cm時,其競爭指數隨胸徑的增大而顯著減少,但胸徑≥20 cm的各徑級油松個體競爭指數隨徑級變化不顯著,占油松總體的25.71%。

圖4 側方擠占和上方遮蓋隨對象木徑級的變化Figure 4 Side and above competition varies with diameter class of objective tree

圖5 油松對象木競爭指數隨徑級的變化Figure 5 Competition index of objiective tree of P.tabuliformis varies with diameter class
3.3.2 油松種內、種間競爭隨競爭木徑級的變化 從表4可見:競爭木胸徑越大,其競爭強度越大,上方遮蓋占總競爭強度的比例也越大。當競爭木胸徑為15~20 cm時,其種內、種間競爭強度最大,分別為868.87和32.26,種內競爭遠大于種間。種內競爭的競爭木株數、競爭強度隨徑級呈先增大后減小的趨勢,而種間競爭的競爭木株數、競爭強度總體上隨徑級呈先減小后增大的趨勢,總體上各徑級的種間競爭木株數和競爭強度遠小于種內。
3.3.3 油松所受競爭強度隨樹種的分布規律 由表5可見:油松種內競爭強度為2116.03,占93.64%,遠大于種間,這與其多為純林的特性相符。油松人工林種內及主要伴生種間的競爭強度順序從大到小依次為油松種內,白蠟,榆樹,北京丁香,樸樹,杜梨,臭椿,桑,槐Sohpora japonica,栓皮櫟,山杏Armeniaca sibirica, 山楂Crataegus pinanatifida,白梨Pyrus bretschneideri。
4 討論和結論
4.1 模型適用性
本研究的油松生物量與d_α_CIi,Hegyi競爭指數服從冪函數關系,與CIOi為指數函數。大部分學者認為 Hegyi競爭指數與林木大小參數服從冪函數[1,3,11-12,15,26], 本研究支持了該觀點。 而惠剛盈等[11]建立的基于交角的競爭指數與林木大小參數間擬合的最優模型卻不為冪函數,與本研究結果不同,這可能是本研究選取競爭指數的胸徑比參數替換了其所選用的大小比數造成的,不同結構的競爭指數所擬合的最優模型不同。
d_α_CIi,CIOi和Hegyi競爭指數均反映了油松生物量受競爭強度增大而減小的規律,3類競爭指數的R2損失量均較小,說明其回歸模型泛化能力均較強;其中,d_α_CIi的決定系數最高,損失量最少,與生物量擬合的離群點較少,說明其對油松所受競爭強度的量化穩定,能較好處理如多主干油松所受競爭強度等特殊情況。因此,d_α_CIi能更好量化北京典型油松人工林競爭強度。
CIOi表現最差,這可能受油松樹冠特征影響。油松人工林多為同齡林,林木大小分化不明顯,作為喜陽物種,油松樹冠的中下層側枝易在激烈的競爭中因光環境過弱而枯死,而其上層側枝因生長年限不足而偏短,從而導致其冠幅較小,樹冠重疊面積偏少,所計算競爭指數可能低于林木實際所受競爭強度。且林木間劇烈的競爭易導致樹冠的可塑性變異[27],令測量的冠幅大小和空間位置的準確性偏低,從而使競爭強度量化不夠準確。因此,該競爭指數表現較差。
北京地區油松人工林因長期缺乏經營管理,其林分密度過大,林木間距不足,光環境過弱,為提高競爭力獲取更多光照,林木將更多的能量用于樹高生長[28]。因此,北京油松人工林中林木樹高是競爭力的重要表現形式。本研究中選取的基于交角的競爭指數d_α_CIi優于Hegyi競爭指數,這與惠剛盈等[11]建立的基于交角的林木競爭指數優于Hegyi競爭指數的結果相似,可能樹高與間距比構成的交角因子更好地反映了林木在水平和垂直方向上的競爭壓力,所以競爭指數d_α_CIi效果更為理想。
不同競爭指數側重點和適用性不同[26],d_α_CIi在量化油松純林競爭強度上表現較好,能具體分析其上方遮蓋和側方擠占的競爭強度,但是否對油松混交林也能保持較高的適用性有待進一步深入研究。
4.2 油松人工林競爭現狀及建議
油松對象木徑級正態分布曲線略正偏,這主要是林木對光等有限資源的不對稱競爭引起的[29]。在溫帶森林里競爭壓力通常是阻礙林木生長甚至導致其死亡的主要因素[30],油松人工林過強的競爭壓力,致使油松總體胸徑偏小,小徑級油松受競爭影響不斷死亡。而林內過高的郁閉度、過厚的林下凋落葉層[31],可能共同導致了其林下更新困難,小徑級林木不斷死亡卻難以得到補充,林分難以進一步形成近自然的異林齡結構。

表5 各樹種競爭強度特征Table 5 The competition according to species
采用d_α_CIi對1054株油松對象木和4216株競爭木競爭強度的量化研究表明:油松所受競爭強度隨其胸徑增大而減小,小徑級油松受側方擠占和上方遮蓋的強度均較大,且兩者比例相差不大,隨胸徑的增大,其來自上方遮蓋的比例逐漸減小甚至消失。當胸徑≥20 cm(約油松林中前1/4),油松所受競爭強度隨徑級的增大變化不顯著,這與項小燕等[1]對大別山五針松Pinus dabeshanesis,曾思齊等[3]對青岡櫟Cyclobalanopsis glauca,沈琛琛等[26]對蒙古櫟Quercus mongolica和邢海濤等[13]對馬尾松Pinus massoninana的研究結果相似。這可能是因為小徑級油松獲取光照、水分等資源的能力較弱,所受壓力較大,而大個體油松居于上層林冠,資源獲取能力更強,因而受其他林木競爭影響較小,并能有效抑制競爭木生長,從而進一步穩定其競爭優勢。除5~10 cm徑級外,油松種間平均競爭強度均小于種內,說明油松個體水平種內競爭更激烈。
油松種內競爭強度為2116.03,占93.64%,所受競爭壓力主要來自種內,這與油松林中其他林木數量過少有關。在油松人工林的種內及主要伴生種間的競爭強度排序中,榆樹的數量僅約占北京丁香和樸樹的1/3,但受林木種類各自生長特征影響,其個體較大,競爭強度普遍較高,進而導致榆樹排序反而靠前,說明林木競爭受數量、大小和種類共同影響,這與項小燕等[1]的研究結果相同。
種內競爭的競爭木株數、競爭強度隨競爭木徑級呈先增大后減小的趨勢,而種間競爭的競爭木株數、競爭強度總體上呈先減小后增大的趨勢,且各徑級的種內競爭木株數和競爭強度總體上遠大于種間。這印證了在許多植物群落中種內競爭往往比種間競爭更強是局域尺度上物種穩定共存的重要條件,也是相鄰尺度上穩定共存機制運作的普遍規律[32],亦表明油松在競爭中占主導地位,其他林木受其競爭排擠,大多以小徑級或大徑級的形式在油松林垂直空間的兩端獲取生存空間,兩者形成物種間的空間分離。這種空間分離能減弱種間競爭,阻止或減緩競爭優勢物種對競爭劣勢物種的競爭排除作用,有利于維持物種共存,促進生物多樣性維持[33],這也說明生態位差異在相鄰空間尺度上的群落動態中發揮著重要作用[32]。
林分密度過高易導致生物量及生產力下降[34],為改善油松林結構,提高其質量,針對其缺乏幼苗、林分密度和競爭強度過高的現狀,應合理稀疏和開辟林窗促使幼苗樹種的形成及林木間的空間分離,以降低林木競爭強度、提高生物多樣性。基于油松林競爭強度的空間分布對局部競爭過強的林木進行合理稀疏,重點圍繞胸徑<20 cm的目標樹開展擇伐釋壓經營管理措施,使其在中小徑級時獲得足夠的空間、光照、水分等資源,減少競爭木對有限資源的掠奪,促使小徑級林木的正常生長和林下更新,以改善油松人工林的結構,提高其生態服務功能。
4.3 結論
d_α_CIi競爭指數能比其他競爭指數更準確的量化北京典型油松人工林的競爭強度。林木競爭受數量、大小和種類共同影響。油松種內競爭大于種間及兩者間的垂直空間分離是林內物種共存的重要條件。基于油松林競爭指數的空間分布,對目標樹周圍胸徑<20 cm的競爭木進行合理撫育間伐,能降低目標樹所受競爭強度,改善油松林結構,提高油松林生態系統服務功能。
