低溫脅迫對不同基因型茄子種子萌發的影響
2019-10-30 01:21:52朱宗文吳雪霞張愛冬田守波查丁石
江西農業學報 2019年10期
朱宗文,吳雪霞,張愛冬,田守波,查丁石
(上海市農業科學院 園藝研究所/上海市設施園藝技術重點實驗室,上海 201106)
茄子(SolanummelongenaL.)別名落蘇,原產于印度地區,在我國已有1400~1500年的栽培歷史[1-2]。我國栽培的茄子品種眾多,按照居民的栽培方式和飲食習慣的不同,在遺傳變異的基礎上,經過多年的自然選擇和人工選擇,形成了形狀各異、顏色多樣、軟硬度差異明顯的眾多茄子生態類型,具有廣泛的地域性特征。至今在我國的西部偏遠地區還有一些茄屬野生種,這些野生種大多較當前栽培品種生長勢強,抗逆性強,可以作為優良的育種資源[3]。茄子作為一種喜溫性蔬菜,對低溫敏感性較強。其植株生長發育的適宜溫度為22~30 ℃,低于15 ℃時植株生長緩慢,低于5 ℃時莖葉受到傷害,0 ℃以下就會被凍死[4]。在低溫脅迫下,茄子幼苗生長受到抑制,光合作用效率降低[5],植株受到氧化傷害[6],最終茄子的產量和品質大大降低。隨著栽培設施和栽培技術的不斷改進,人們對保護地茄子生產的要求越來越高,對耐低溫性品種的需求也越來越迫切。目前,國內相關科研單位和種子公司很少有耐低溫的保護地專用茄子品種[7-8],而其他茄果類蔬菜像番茄[9]和辣椒[10-11]有較多的保護地低溫專用品種的介紹。在低溫條件下茄子生產主要還是通過嫁接這一途徑來實現[12]。因此,選育出耐低溫性強的保護地專用茄子品種極其重要。我們以本課題組長期保存和積累的各種類型的茄子高代自交系材料為研究對象,通過觀察在不同低溫條件下種子的萌發情況,對低溫脅迫下不同基因型茄子材料種子的耐低溫性做出評價,希望為茄子耐低溫性鑒定及耐低溫材料的篩選提供依據,為茄子低溫育種材料的創制提供一定的幫助。
1 材料與方法
1.1 實驗材料
10份供試茄子材料(表1)為上海市農業科學院園藝研究所茄子課題組保存的高代自交系材料。經過預實驗驗證,同一批種子的發芽率一致,整齊度高。
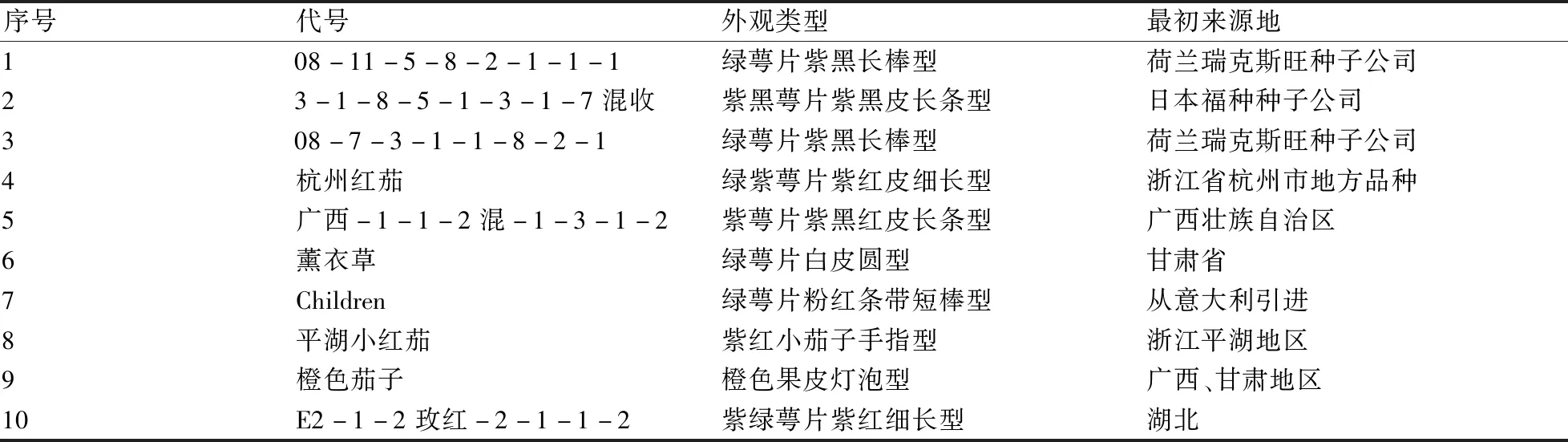
表1 供試茄子品種/材料的情況
1.2 低溫處理
將不同類型的茄子材料種子分別置于發芽盒中,在發芽紙上加入適量蒸餾水后分別放入設定不同溫度的人工智能培養箱進行低溫處理,溫度設定分別為10、15、20、25 ℃。
1.3 種子萌發測定
用透明塑料發芽盒(12 cm× 12 cm× 5 cm)測定種子的萌發情況。將種子放入鋪有兩層發芽紙的發芽盒中,置于不同溫度培養箱內發芽。每個溫度處理100粒種子,3次重復。從第2天起逐日統計種子發芽數。以第14天作為發芽結束時間,發芽結束后統計各項發芽指標。相關發芽指標的計算公式為:發芽率(Gr)=∑Gt/NT×100%;相對發芽率=處理種子的Gr/對照種子的Gr;發芽勢=規定時間內發芽種子總數/供試種子總數×100%;發芽指數(Gi)=∑(Gt/Dt);活力指數(Vi)=Gi×S。其中,Gt表示在t日時的發芽數,Dt表示相應的發芽天數;NT表示種子總數;S表示發芽終期幼苗的高度(cm)。
1.4 幼苗冷害指數和恢復能力的測定
幼苗冷害指數和恢復能力的測定參照王孝宜等[13-14]的方法(作了適當改進)。將發芽結束后的幼苗繼續在人工智能培養箱中進行低溫處理7 d,每日向發芽盒中加入適量的蒸餾水,保證發芽紙濕度適中;7 d后,統計幼苗的冷害指數CI。全部死亡時統計CI為最大值1,全部正常時統計CI為最小值0。統計結束后,將幼苗全部移到正常溫度的培養箱進行1周的恢復培養,結束后統計幼苗的恢復能力,恢復能力以恢復正常的幼苗數占總幼苗株數的百分比表示。對每一幼苗的冷害癥狀進行分級:0級,無冷害癥狀;1級,僅子葉邊緣有輕度皺縮萎蔫;2級,子葉大部分區域變色萎蔫;3級,子葉整體萎蔫;4級,整株死亡。冷害指數(CI)=(1×S1+2×S2+3×S3+4×S4+0×S0)÷(低溫脅迫總株數×4)。式中,Si(i=0,1,2,3,4)為相應冷害等級的苗數。
1.5 統計方法
用SPSS 16.0統計軟件和Excel對試驗數據進行統計、作圖及相關分析。
2 結果與分析
2.1 低溫處理對不同類型茄子種子發芽率的影響
不同類型茄子種子經低溫處理后,各溫度處理間種子發芽率存在很大的差異性(表2)。其中1、5、8號三個基因型種子在10 ℃和15 ℃溫度條件下均不發芽,在20 ℃條件下發芽率也較低且顯著低于正常溫度下的;4號和7號種子在10 ℃下不發芽,在15 ℃下發芽率極低;2號和6號種子在15 ℃下的發芽率顯著高于在20 ℃下的;3、6和10號種子在10 ℃下不發芽,在15 ℃下發芽率較高且與正常溫度下差異不顯著;9號種子是唯一在10 ℃低溫條件下還可以發芽的,且在15 ℃和20 ℃條件下發芽很好,與正常溫度下無明顯差異。總體來看:9號發芽最好,發芽耐低溫性最強;其次是3號、6號和10號;發芽耐低溫性較差的是1號、5號和8號。
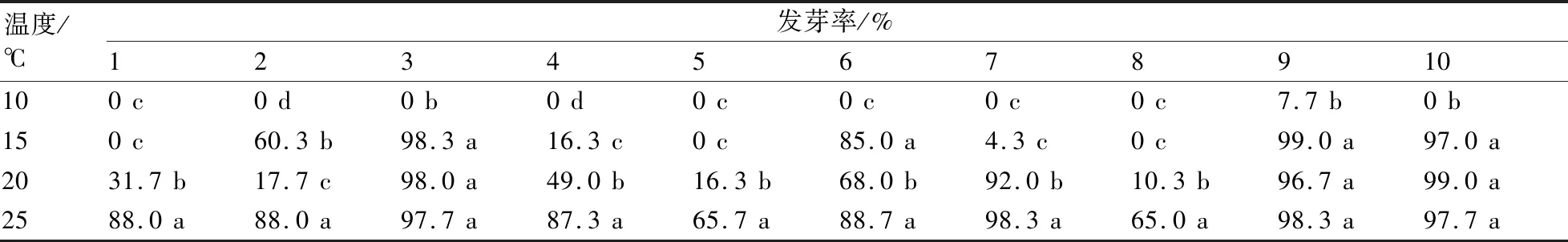
表2 不同低溫處理對茄子種子發芽率的影響
注:同一列數據后附不同小寫字母代表在不同溫度處理間存在顯著差異(P<0.05)。下同。
2.2 不同低溫處理對茄子種子發芽勢的影響
發芽勢反映了發芽初期種子活力的強弱和發芽的整齊度。經低溫處理后,各類型茄子的發芽勢呈現出很大差異性(表3)。發芽勢最高的是9號,它是唯一在10 ℃低溫條件下具有發芽勢的茄子品種,其在15 ℃和20 ℃條件下發芽勢與對照無顯著性差異;6號在15 ℃下的發芽勢與對照無顯著差異,并顯著高于在20 ℃下的發芽勢;10號在20 ℃下的發芽勢與對照無顯著差異;其他類型茄子種子的發芽勢隨著溫度下降均顯著低于前一溫度梯度下的發芽勢。綜合發芽率和發芽勢可以看出:9號不僅發芽率在低溫條件下較高,發芽整齊度也很好,15 ℃的低溫對其發芽勢無顯著影響;6號和10號的發芽勢表現出一定的整齊度,但是總體上弱于9號;1、2、4、5、8號的發芽勢在各低溫條件下表現較弱,在正常溫度條件下也較弱,這說明發芽整齊度與其本身的基因類型和種子存貯狀況存在一定的關系。
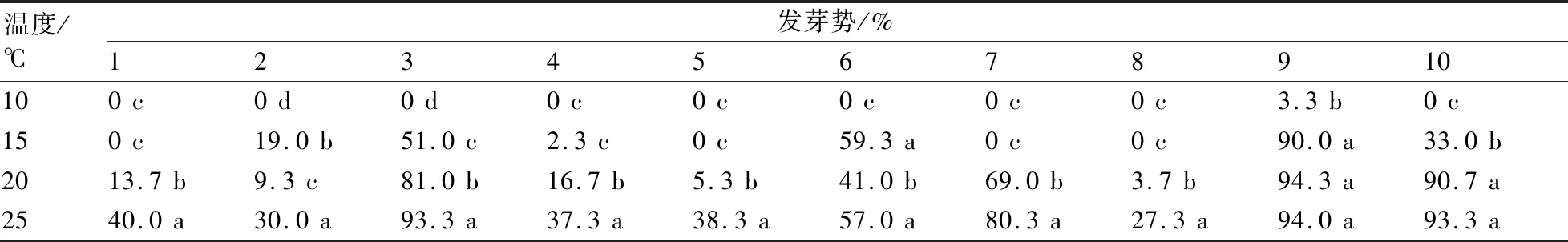
表3 不同低溫處理對茄子種子發芽勢的影響
2.3 不同低溫處理對茄子種子發芽指數的影響
經低溫處理后,不同類型茄子種子呈現出不同的發芽速率,我們測定其發芽指數來衡量其發芽速率。從表4中可以看出:在10 ℃條件下,只有9號存在發芽指數,并與其它溫度條件下存在顯著性差異,而在15、20、25 ℃處理間其發芽指數不存在顯著性差異;3號在20 ℃條件下的發芽指數與對照無顯著性差異,在15 ℃下與正常溫度下存在顯著性差異;6號在15 ℃和20 ℃下的發芽指數沒有顯著性差異,但與對照存在顯著性差異;其它各類型種子的發芽指數隨溫度降低與前一溫度梯度均呈現出顯著性差異。
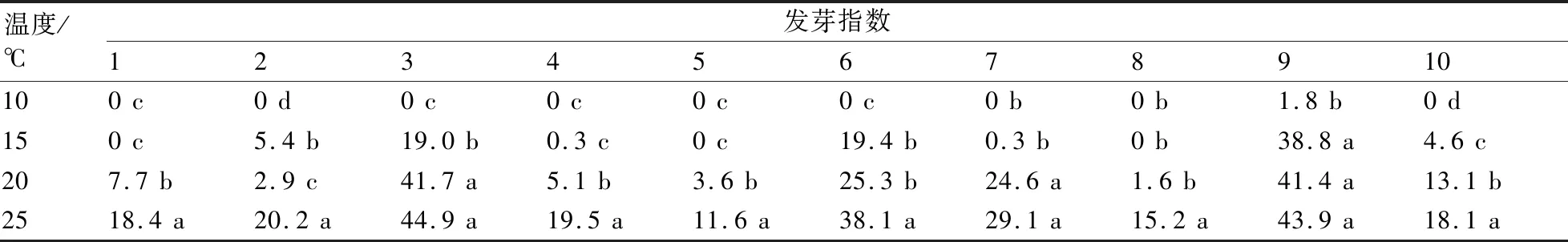
表4 不同低溫處理對茄子種子發芽指數的影響
2.4 不同低溫處理對茄子種子活力指數的影響
如表5所示:9號茄子幼苗具有最強的活力指數,其在10 ℃下活力指數很低,在其他3種溫度處理間也存在顯著性差異,但總體活力指數最高;2號的活力指數在10 ℃和15 ℃條件下差異不顯著,是所有實驗品種中唯一一個在不同溫度梯度間活力指數差異不顯著的;其他各實驗品種的活力指數在不同溫度處理間均呈現出顯著性差異,說明環境低溫對其活力指數下降產生了顯著影響。
2.5 不同低溫處理對茄子種子萌發幼苗冷害指數的影響
經低溫處理的種子萌發后,將幼苗繼續進行7 d的低溫生長觀測,測定各類型茄子幼苗的冷害指數。從圖1中可以看出:9號在各個低溫條件下冷害指數最低,是所有實驗品種中最耐低溫的;2號和10號在15 ℃條件下具有一定的耐低溫性;1號、5號和8號在15 ℃條件下不發芽或幼苗完全死亡;在20 ℃條件下各品種的冷害指數較低,5號和8號受影響較大。綜合來看,9號最耐低溫,10號、2號次之,5號和8號最不耐低溫。
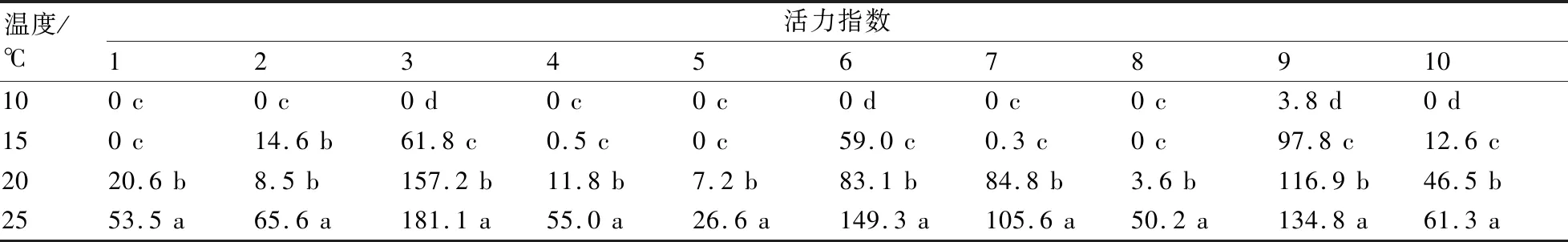
表5 不同低溫處理對茄子種子活力指數的影響
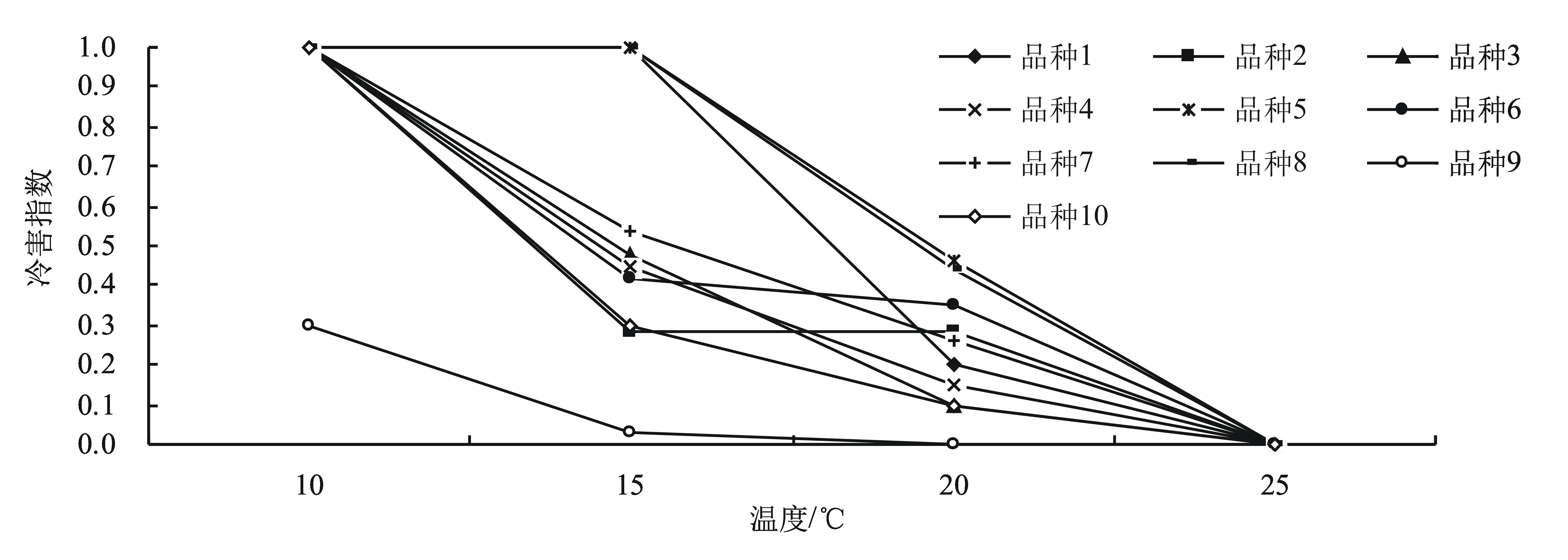
圖1 低溫處理后各類型茄子幼苗受冷害情況
2.6 不同低溫處理對幼苗恢復能力的影響
將經低溫處理的各類型幼苗移入正常溫度下進行幼苗恢復能力的測定。從圖2可以看出:在10 ℃條件下只有9號的幼苗具有一定的恢復能力;經15 ℃處理后,6號和9號幼苗的恢復能力超過50%,而4號的恢復能力最低,在25%左右;經20 ℃低溫處理后,各品種幼苗的恢復能力較強,只有4號低于50%;1號、5號和8號由于其種子在10 ℃和15 ℃下不發芽,因而沒有存活幼苗。從幼苗的恢復情況可以看出,9號的耐低溫性最強,6號和10號次之,1、4、5和8號最差。

圖2 低溫處理后各類型茄子幼苗的恢復情況
3 討論
當植物受到低溫脅迫時,在各種環境信號傳導物質作用下植株體內會發生一系列的生理生化反應。這些生理生化反應是植株本身對逆境一種自我保護的應激反應,更多是存在于生理生化物質的積累和代謝上[15-17]。張曉艷等研究表明,在所有受低溫脅迫的茄子嫁接苗中,耐低溫材料的超氧化物歧化酶(SOD)、過氧化物酶(POD)和過氧化氫酶(CAT)活性均高于不耐低溫材料的[18]。吳雪霞等發現經低溫處理后茄子幼苗的丙二醛(MDA)含量顯著增加,葉片的SOD、POD、CAT、抗壞血酸過氧化物酶(APX)活性均顯著升高,且其脯氨酸(Pro)和可溶性蛋白質含量均顯著增加[19]。董愛玲等研究發現低溫馴化顯著增加了茄子幼苗的可溶性糖、可溶性蛋白含量,提高了SOD、POD、CAT活性[20]。這些研究結果均表明茄子在低溫脅迫下,其植株體內會發生一系列的生理生化反應,且在不同的品種材料之間,變化存在較大差異。因而選育耐低溫性強的茄子材料對茄子育種及了解相關基因的功能有著極其重要的作用。本實驗通過對大量的高代自交系茄子材料進行低溫種子活力測定,從中找到了一些萌發耐低溫性較強的材料,其中9號表現最為突出,在各種溫度梯度下,其發芽率、發芽勢、發芽指數和活力指數均表現出很高水平,甚至在10 ℃低溫條件下仍然有部分種子可以成功發芽且幼苗具有較強的活力和恢復能力,可以作為今后茄子耐低溫育種的優良高代自交系材料。
種子活力主要受自身基因遺傳性、種子發育成熟度和貯藏環境因子三方面的影響[21]。自身的遺傳性決定了種子活力強度的可能性,其他環境因素也會對種子活力最終的表現起到重要的作用。我們可以采取一定的措施來促使種子表現出其強的活力,有研究表明預冷處理可以有效地破除一些種子的生理休眠,如麥類種子發芽前放入濕潤的發芽床上,在5~10 ℃下處理3 d,可以達到破除生理休眠的效果[22];喬海明發現用5 ℃處理野豌豆種子可以顯著提高其種子活力[23];憨宏艷等研究表明不同土壤濕度條件下低溫處理會增強水青樹種子的發芽率,并對種子活力起到一定的促進作用[24]。這些研究都表明在環境低溫條件下,種子活力受到了正面影響,這可能是由其遺傳物質的特異性所決定的。種子萌發是一個極其復雜的過程,不僅涉及到各種貯藏物質的分解,還涉及到許多萌發過程中相關基因的調控。白雪等以冷敏感作物蓖麻為實驗材料,構建15 ℃(低溫)和25 ℃(適溫)兩種萌發條件下完全展開的子葉cDNA文庫,利用Illumina測序技術進行轉錄組測序(RNA-Seq),篩選出了低溫上調差異表達基因243個[25]。包崇來等從大量茄子育種材料中選出了耐低溫基因型材料E659,其幼苗在4 ℃低溫下處理10 d后仍能保持正常的生長狀態,沒有受到明顯傷害[26]。本實驗中2號和6號茄子材料在15 ℃條件下表現出了較強的耐低溫能力,其發芽率、發芽勢均顯著高于20 ℃條件下的;9號材料則在15 ℃和20 ℃溫度下表現出了和對照一致的種子活力,并在10 ℃低溫條件下仍然表現出了較強的種子活力。這些表明不同基因型材料的特定遺傳物質決定了其耐低溫能力的強弱。目前對茄子的耐低溫生理機制研究較多,但獲得極耐低溫茄子基因型材料的報道較少。本實驗為茄子耐低溫抗性種質資源的創制和耐低溫新品種的選育提供了新的基因源,可作為茄子耐低溫分子育種的新材料。
