飼喂幼蟲糞便提取物對馬鈴薯塊莖蛾生殖的影響
2019-10-29 07:31:34李宗鍇胡純華
江西農業學報 2019年10期
劉 燕,李宗鍇*,韓 瑞,王 靜,胡純華,肖 春**
(1.云南農業大學 植物保護學院,云南 昆明 650201;2.云南農業大學 職業與繼續教育學院,云南 昆明 650201)
0 前言
馬鈴薯塊莖蛾(Phthorimaeaoperculella)是一種典型的寡食性害蟲,寄主范圍僅限于茄科植物,且最嗜食馬鈴薯塊莖[1]。該蟲是馬鈴薯種植業上的最重要害蟲之一,在塊莖貯存期為害尤其嚴重,其中貯存期超過半年以上的塊莖被害率可達100%[2]。因此,研發安全、高效的防治技術,對于控制馬鈴薯塊莖蛾的發生與為害極為重要。
性誘劑應用是防治害蟲的重要技術之一,具有安全、高效、專一性強、不傷害天敵、不污染環境的優勢。馬鈴薯塊莖蛾的性信息素研究早有報道[3],但性誘劑在生產實踐中的應用效果極其少見[4],原因可能和馬鈴薯塊莖蛾存在著孤雌生殖的爭論有關。因為性誘劑的作用方式是阻斷昆蟲兩性之間的交配行為,進而達到降低害蟲后代種群的目的。因此,性誘劑只適合應用于兩性生殖的害蟲種類的防治,對于具有孤雌生殖特性的種類則效果不佳[5]。
關于馬鈴薯塊莖蛾是否存在著孤雌生殖現象一直存有爭論。有報道顯示,馬鈴薯塊莖蛾的孤雌生殖率可達50%以上[6],而有些研究則報道該蟲并不存在孤雌生殖或者發生率極低[7]。最新研究結果顯示,在馬鈴薯塊莖蛾中確實存在著孤雌生殖現象;孤雌生殖的發生與塊莖中的幼蟲密度緊密相關[8]。也就是說誘發孤雌生殖的因素在幼蟲體內就已經形成了。那么,這種因素是否會隨糞便排出體外、并且具有影響雌蟲孤雌生殖發生的活性?孤雌生殖個體的產卵特征是什么?回答這些問題將有助于進一步揭示馬鈴薯塊莖蛾生殖方式轉變及種群自我調節的機制。為此,作者開展了飼喂幼蟲糞便提取物對馬鈴薯塊莖蛾生殖及產卵的影響的研究。
1 材料與方法
1.1 蟲源
馬鈴薯塊莖蛾采自云南省宣威市,幼蟲以正常密度(15頭/130 g塊莖,ND)用馬鈴薯塊莖飼養。當卵孵化后,將初孵幼蟲接入到新鮮塊莖上,之后幼蟲會蛀入到塊莖內取食、生長。將接有幼蟲的塊莖置于養蟲籠(24 cm×24 cm×24 cm)內的細沙上。幼蟲在取食過程中會從塊莖中將糞便排出至塊莖表面形成黑色塊狀物,老熟后從塊莖中脫出并入沙、結繭、化蛹。成蟲羽化后按1∶1配對置于產卵盒(Φ=13 cm,h=14.5 cm)中,成蟲產卵于濾紙表面[9]。每天用10%蜂蜜水(FM)為成蟲提供補充營養,并定時收集蟲卵備用。室內環境條件:溫度(25±2)℃,光周期L∶D = 14 h∶10 h,相對濕度為50%~70%。
按照上述方法,將初孵幼蟲以高密度(45頭/130 g塊莖,HD)接入塊莖中,之后4~6 d內收集幼蟲產生的新鮮蟲糞。取5 g新鮮蟲糞和10 g蜂蜜加蒸餾水配制成100 mL溶液,充分攪拌后靜置5 h得蟲糞提取物(ELF)。將提取物過濾后取濾液于冰箱(<-5 ℃)中保存備用。
1.2 飼喂幼蟲糞便提取物對孤雌生殖發生的影響
將馬鈴薯塊莖(80±10)g置于220 mL紙杯中,并在杯中放置浸有4 mL補充營養液的棉團(Φ=1 cm),將1頭剛羽化的ND雌蟲置于杯中并用尼龍紗網封口以阻隔雌蟲與雄蟲的接觸。8 d后觀察馬鈴薯上有無幼蟲糞便出現,有蟲糞者視為孤雌生殖發生。之后將有蟲糞的塊莖移入到養蟲籠中,直至蛹出現,待成蟲羽化后記錄雌雄比。
利用上述方法,分別測定:ND雌蟲+10%蜂蜜水(對照,ND/FM)、ND雌蟲+蟲糞提取物(ND/ELF)、HD雌蟲+10%蜂蜜水(HD/FM)等3種組合下雌蟲的孤雌生殖率。每處理觀察30頭雌蟲,重復4次。
1.3 飼喂幼蟲糞便提取物對雌蟲產卵量及卵粒的影響
1.3.1 對未交配雌蟲產卵的影響 采用步驟1.2的方法用幼蟲糞便提取物飼喂雌蟲。將5頭剛羽化的ND雌蟲置于產卵盒中,每天飼喂4 mL幼蟲糞便提取物(ELF),分別以喂食4 mL蜂蜜水的ND雌蟲和HD雌蟲為對照。每天收取卵粒,直至雌蟲死亡。使用體視顯微鏡檢查卵的外觀并統計卵量。重復4次。
雌蟲卵粒從外觀上看可分為3類:正常卵、畸形卵和干癟卵。正常卵飽滿,橢圓形;畸形卵表面嚴重凹陷、有皺褶;干癟卵只剩卵殼,也稱無效卵(圖1)。
1.3.2 對已交配雌蟲產卵的影響 將1日齡ND雌蟲、ND雄蟲分別置于產卵籠中喂食幼蟲糞便提取物(ELF)并保持1 d,之后取5對雌、雄蟲(2日齡)按1∶1比例配對置于其它產卵盒中,喂以蜂蜜水。另外分別取5頭雌、雄蟲(ND/ELF)與同日齡異性(ND/FM,僅取食蜂蜜)按1∶1比例配對后置于產卵盒中,每天喂以蜂蜜水。重復4次。
另取5對2日齡ND雌雄蟲(僅喂食蜂蜜,CK1)、5對2日齡HD雌雄蟲(僅喂食蜂蜜,CK2)分別置于產卵盒中以便完成交配、產卵。每天喂以蜂蜜水(FM)。重復4次。次日開始收集卵粒,直到雌蟲死亡,統計單雌產卵量和孵化率。
1.4 數據分析
采用統計軟件SPSS 17.0進行實驗數據分析,利用Duncan檢驗進行差異顯著性分析(ANOVA,α=0.05)。
2 結果與分析
2.1 飼喂幼蟲糞便提取物對馬鈴薯塊莖蛾孤雌生殖發生的影響
實驗結果如表1所示,以正常密度[15頭/(130±2)g塊莖,ND]飼養幼蟲時,在馬鈴薯塊莖蛾中不會出現孤雌生殖現象;而以高密度[45頭/(130±2)g塊莖,HD]飼養幼蟲時,雌蛾的孤雌生殖率可達10%左右,且以孤雌生殖模式所繁殖的后代雌雄比為1.4。用正常密度飼養幼蟲獲得的雌蛾,如果喂以高密度幼蟲的糞便提取物(ELF),則可誘發孤雌生殖的發生,其發生率可達2.5%左右,而且由孤雌生殖而來的后代雌雄比為2.0。
2.2 飼喂幼蟲糞便提取物對馬鈴薯塊莖蛾產卵的影響
2.2.1 飼喂幼蟲糞便提取物對未交配雌蟲產卵的影響 從實驗結果可以看出,未交配雌蛾產的正常卵外觀飽滿,卵黃明顯,呈橢圓形;而畸形卵表面通常有凹陷、皺褶,卵黃明顯;無效卵僅有卵殼,內無明顯卵黃(圖1)。其中無效卵為主要類型,正常卵所占比例最低(表2)。

1.正常卵;2.畸形卵;3.干癟卵。

表1 飼喂幼蟲糞便提取物對馬鈴薯塊莖蛾孤雌生殖發生的影響
注:數字后面的字母a、b、c表示不同處理間的差異顯著性(方差齊性檢驗,df1=2,df2=9,P=0.184;ANOVA,F=31.978,P<0.001)。
在沒有交配的條件下,不論是由正常密度還是高密度幼蟲飼養而獲得的雌蟲產卵量均較低,單雌產卵量不超過20粒;而在飼喂ELF后,ND雌蟲產卵量也極低(平均19.6粒),與前兩者不存在顯著差異(表2)。
在僅喂食蜂蜜時,HD雌蟲產生的卵粒中正常卵粒所占比例顯著高于ND雌蟲。而ND雌蟲在取食ELF后正常卵粒所占比例得到顯著提高(表2)。

表2 飼喂幼蟲糞便提取物對馬鈴薯塊莖蛾未交配雌蟲產卵的影響
注:字母a、b、c表示不同處理間的差異顯著性(α=0.05)。其中,正常卵(方差齊性檢驗,df1=2,df2=9,pP=0.619;ANOVA,F=6.523,P=0.018),無效卵(方差齊性檢驗,df1=2,df2=9,P=0.995;ANOVA,F=5.056,P=0.034)。
實驗結果也同時顯示,在未交配狀態下ND雌蟲(僅喂食蜂蜜)所產出的卵中無效卵比例高達77.6%,顯著高于HD雌蛾(僅喂食蜂蜜)的66.4%。而喂食ELF的ND雌蛾產下的卵粒中無效卵占73.6%,與另外兩種處理均無顯著差異(表2)。畸形卵在總卵粒中所占比例在3種處理間均不存在顯著差異(表2)。
2.2.2 飼喂幼蟲糞便提取物對交配雌蟲產卵的影響 從單雌產卵量來看,HD雌蟲(CK2)在交配后的產卵量最低,僅為142±16.7粒,顯著低于ND雌蟲(CK1)的211±20.3粒。如果單獨給ND雌蟲、ND雄蟲喂ELF,或者同時給ND雌雄蟲喂食ELF,則雌蟲交配后的落卵量與對照(CK1)相比均無顯著差異。但如果讓ND雌雄蟲同時取食ELF后再進行交配,則雌蟲單雌產卵量可達233.0±18.0粒,顯著高于HD雌蟲(CK2)的產卵量,而與ND雌蟲(CK1)無顯著差異(表3)。
在僅喂食蜂蜜的條件下,HD雌蟲(CK2)卵的孵化率最低,僅為(81.8±1.4)%,顯著低于ND雌蟲(CK1)的(92.6±0.9)%。與對照(CK1)相比,ND雄蟲在取食ELF后若再與雌蟲(僅喂食蜂蜜)交配,則卵孵化率僅為(82.0±2.3)%,顯著低于CK1的(92.6±0.9)%,與CK2的(81.8±1.4)%無顯著差異。如果讓ND雌蟲取食ELF后與ND雄蟲(僅喂食蜂蜜)、或者ND雌雄蟲同時取食ELF,則卵孵化率均與CK1無顯著差異(表3)。
至于雌蟲所產畸形卵量,在各處理間均無顯著差異。
3 討論
孤雌生殖是昆蟲中比較常見的一種生殖方式。有許多種類既可以行兩性生殖,也可行孤雌生殖[10,11]。許多研究結果表明,孤雌生殖的發生是對極端環境的一種適應,比如應對食物/寄主匱乏[12,13]、適應高溫[14]等。
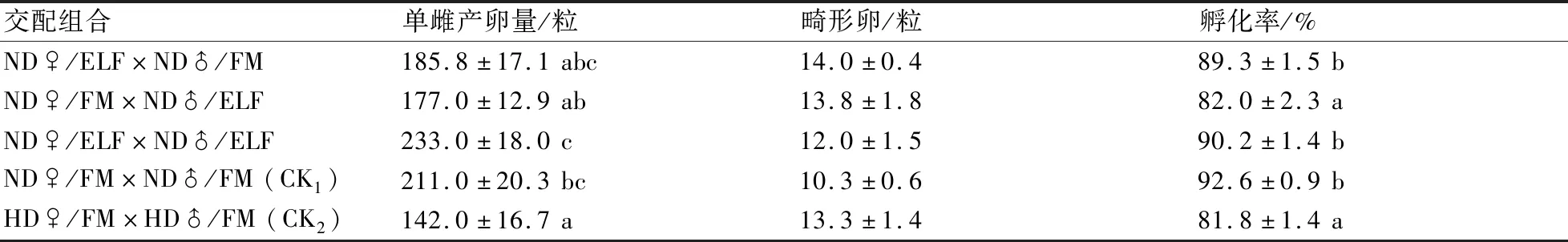
表3 飼喂幼蟲糞便提取物對馬鈴薯塊莖蛾交配雌蟲產卵的影響
注:孵化率是指全部落卵的孵化率。字母a、b、c表示不同處理間的差異顯著性(α=0.05),其中單雌產卵量(方差齊性檢驗,df1=4,df2=15,P=0.822;ANOVA,F=4.059,P=0.02),畸形率為(方差齊性檢驗,df1=4,df2=15,P=0.161;ANOVA,F=1.524,P=0.245),孵化率(方差齊性檢驗,df1=4,df2=15,P=0.605;ANOVA,F=9.965,P<0.0001)。
在馬鈴薯塊莖蛾中,孤雌生殖只有在塊莖幼蟲數量超過一定限度時才會產生[8]。這說明孤雌生殖產生主要是為了應對食物資源匱乏而采取的一種生殖策略。我們的實驗結果顯示,喂食高密度幼蟲產生的糞便提取物ELF能夠誘發孤雌生殖現象的發生(表1)。這一事實說明孤雌生殖誘發因子(Parthenogenesis-inducing factors, PIF)在幼蟲期內即開始發揮作用。這也同時說明,這些誘發因素(PIF)可能隨幼蟲糞便排除體外。這顯示誘發因素可能存在于腸道中,甚至可能與腸道中的微生物有關[14,15]。實驗結果顯示,在未交配時,不論是正常密度幼蟲飼養、還是高密度幼蟲飼養所獲得的雌蟲,其產卵量均很低;在給正常密度雌蛾喂食幼蟲糞便提取物ELF后,未交配雌蟲的產卵量也與前兩者沒有顯著差異(表2)。這說明,PIF沒有刺激雌蟲產卵的功效。
那么,當環境改變后、昆蟲體內的誘發因素PIF被激活時,會使馬鈴薯塊莖蛾產卵發生哪些變化呢? 從實驗結果(表2)可以看出,幼蟲高密度飼養時,未交配雌蛾產下的正常卵量顯著高于正常密度飼養的雌蛾,由高密度幼蟲羽化而來的處女雌蛾(HD♀)產下的正常卵粒可達總卵量的4.5%左右,遠高于正常密度雌蟲(ND♀)的1.7%。更進一步觀察顯示,當誘發因子PIF進入ND♀體內后,則可誘使未交配雌蛾體內正常卵粒的比例顯著增加。如ND♀在取食幼蟲糞便提取物(ELF)后,正常卵粒比例亦得到顯著升高,達到3.9%(表2)。這在一定程度上說明,誘發因子PIF具有促進卵粒發育的作用,而正常卵粒的增加正是孤雌生殖發生的物質基礎。這說明蟲體內誘發因子PIF可能參與了卵母細胞發育的調控。這與有關報道的結果較相似[16]。
更進一步的結果顯示,與對照(CK1)相比,在交配前,不論是給ND♀還是ND♂,或者同時給ND♀、ND♂喂食幼蟲糞便提取物,則ND成蟲交配后的單雌產卵量均無顯著變化(表3)。這進一步說明,PIF不會刺激雌蛾產卵。
有意思的是,喂食ELF雖然不會刺激雌蟲產卵,但是實驗結果證實,給予ND♂喂食ELF后再與未接觸過ELF的ND♀交配,則造成卵孵化率顯著下降(表3)。這說明,PIF在一定程度上具有抑制雄蟲生殖力的作用。這種現象在其它昆蟲的孤雌生殖現象中也存在[17]。
綜上所述,孤雌生殖的發生是馬鈴薯塊莖蛾對不利環境的一種適應性表現。即在食物極端匱乏的情形下,為了降低種內競爭、保持種群順利繁衍,種群必須降低自身繁殖力,以實現有限資源的高效利用[18]。而降低種群自身繁殖力是通過PIF來進行調節的,PIF被激活后,可以促進卵粒的發育,以提高孤雌生殖的效率;且交配會顯著提高雌蟲的產卵量,而PIF被激活后,雄蟲的生殖潛力會受到顯著抑制,從而導致種群繁殖力下降。但馬鈴薯塊莖蛾體內的PIF究竟是什么,尚需要進行更深入的研究。
