高效降解海帶芽孢桿菌菌株的篩選及活性成分檢測
2019-10-23 05:44:16張晶晶解學仕周麗李燕王婷
食品研究與開發 2019年20期
張晶晶,解學仕,周麗,李燕,王婷,*
(1.齊魯工業大學(山東省科學院)山東省微生物重點實驗室,山東濟南250353;2.史丹利農業集團股份有限公司,山東臨沂276700;3.山東福瑞達生物工程有限公司,山東濟南250353)
在亞洲國家的沿海地區,海藻作為傳統食物已被消費了幾個世紀,且各種可食用海藻現已越來越多地被用作全球潛在工業應用的成分來源[1]。海藻種類繁多,資源豐富,是重要的海洋資源。在我國,海藻主要以海帶為主。海帶又名昆布、綸布,常作為食品和中藥原料,在《本草綱目》、《神農本草位》、《食用本草》及《中國中草藥匯編》中均有記載。海帶不僅含有豐富的蛋白質、維生素和礦物質,還含有如褐藻酸、褐藻糖膠、褐藻淀粉、海帶多酚、褐藻纖維、甘露醇、高不飽和脂肪酸、巖藻黃質和甾醇類化合物等豐富的生理活性物質[2]。海藻多糖主要包括褐藻膠、褐藻糖膠和褐藻淀粉。作為主要成分的褐藻糖膠已被報道具有的作用包括免疫調節、抗腫瘤、抗病毒、抗菌消炎、抗氧化、抗凝脂、降血糖等[3],其中因褐藻糖膠僅占4%含量就可以阻止癌細胞生長,并引起腫瘤細胞凋亡而引起人們廣泛關注。研究表明,褐藻糖膠抑制了肝癌細胞QGY7703進入對數生長期,從而遏制了腫瘤的增長,褐藻糖膠對Hepes 的抑瘤率可超過50%,且不影響小鼠的正常生長,從而表現出比陽性藥物的優越性[4]。海藻多酚是一大類結構不同的化合物,具有多種生物活性,目前對海藻多酚抗腫瘤活性、抗菌抗病毒活性、抗氧化活性研究較多。其中,鼠尾藻中褐藻多酚對人肝癌細胞株BEL-7402 和人肺癌細胞株A-549 具有較強的抗腫瘤作用,最佳抑制濃度在0.085 mg/L~0.10 mg/L 左右;小黏膜藻粗提物對KB 癌細胞、HT-29 癌細胞具有選擇性細胞毒活性,對正常細胞NIH-3T3 基本上不具有毒性[5]。海藻酸鹽是海藻酸與 Na+、Ca2+、Mg2+結合的線性多糖,有兩個差向異構體組成:β-D-甘露糖醛酸和α-L-古洛糖醛酸[6]。藻酸鹽由于其膠凝、乳化及成膜特性在食品,藥物和醫藥工業中具有應用價值[7]。另外,低分子量的海藻酸鹽具有抗氧化的潛在價值,可增強生物活性[8]。
然而,目前我國海帶的開發主要集中于海帶食品、海藻(帶)肥料和碘、海藻酸、甘露醇等化工原料的提取,海帶中活性成分的提取開發也是只針對一種或一類。例如傳統海帶多糖的提取方法包括熱水提取法、酶提取法、堿提取法,而這些提取方法都存在一定的缺陷。熱水提取的方法采用的溫度是70 ℃~80 ℃,耗能多,提取時間長。酶提取的方法提取的時間長,工業生產需要使用的酶量大,不經濟。堿提取法是現在工業提取海帶多糖的主要方法,但是堿提取方法會造成多糖生物活性的降低。因此,開發一種綠色、高效、環保、全面的提取技術勢在必行。經文獻調研,芽孢菌酶系豐富,具有較強的蛋白酶、淀粉酶和脂肪酶活性,同時還具有降解非淀粉多糖的酶,如果膠酶、葡聚糖酶、纖維素等酶,適用于海帶降解。本研究以天然新鮮海帶為原料,以芽孢桿菌為出發菌株,以海帶發酵黏度、海藻多糖、巖藻糖、海藻酸等海帶活性成分為檢測對象,對能夠高效降解海帶并釋放活性成分的菌株進行篩選,為海帶中活性成分的綜合提取及利用奠定堅實基礎。
1 材料與方法
1.1 材料與試劑
原料:菌株篩選所用新鮮海帶采自渤海灣(山東榮成),發酵用海帶干粉為上述新鮮海帶經自來水洗凈表面泥沙,于55 ℃下干燥48 h,經超微粉碎機粉化后殺菌并儲存。
種子培養基[9],發酵培養基[9],海帶液體培養基[10],LB 固體培養基(除海帶外培養基原料均為分析純):國藥集團化學試劑有限公司。
L-半胱氨酸、無水乙醇、氯化鈣、鹽酸、苯酚(均為分析純):國藥集團化學試劑有限公司;L-巖藻糖:Sigma 公司;細菌基因組提取試劑盒:南京諾唯贊公司;DNA 測序:華大基因。
1.2 儀器與設備
LDZX-50KBS 滅菌鍋:上海申安醫療器械廠;E-clipse E200 顯微鏡:尼康公司;FE20 pH 計:梅特勒-托利多儀器有限公司;CW5-100QA 超微粉碎機:煙臺慧寶設備制造有限公司;SNB-4 數字黏度計:上海精科儀器有限公司;RS232-C 分光光度計:上海元析儀器有限公司;KQ2200B 超聲提取器:昆山市超聲儀器有限公司;HB-100 金屬浴:杭州博日科技有限公司;TProfessional Thermocycler PCR 儀:德國 Biometra 公司;Centrifuge5804R 離心機:德國艾本德公司;Z216M離心機:德國赫姆勒公司。
1.3 方法
1.3.1 菌種篩選
將新鮮海帶加入無菌水中,振蕩培養20 min,取5 mL 水樣加入45 mL 種子培養基,80 ℃處理20 min后 200 r/min 培養 24 h,稀釋 104、105、106倍,平板劃線培養,分離純化后鏡檢。挑取典型菌落接種于海帶培養基,同時設置空白組(未加海帶干粉)和對照組(未加菌種)。37 ℃振蕩培育24 h,分別檢測發酵海帶黏度,使海帶黏度明顯增加的菌株為目的菌株。菌株純化后儲存在30%甘油中于-80 ℃保存。
1.3.2 海帶降解及產物制備
將篩選得到菌株接種至種子培養基中培養至指數期,試驗組(接種量2%)、對照組(接種量為0)分別將種子液轉接至添加5%海帶干粉的發酵培養基中,37 ℃振蕩培育 24 h。將發酵樣品 4 ℃,10 000 r/min 離心10 min。取其上清液,待測。
1.3.3 海帶發酵液中多糖含量的檢測
采用SNT 4260-2015《出口植物源食品中粗多糖的測定苯酚-硫酸法》[11]檢測發酵液上清中的多糖含量。
吸取樣品200 μL 于50 mL 離心管內,提取多糖。提取結束后,4 000 r/min 離心10 min 后棄去上清液。用水將洗滌、離心后得到的不溶物轉移入圓底燒瓶,超聲提取 30 min,重復 2 次。冷卻至室溫(約 25 ℃),過濾,將上清液轉移至200 mL 容量瓶中,殘渣洗滌2 次~3 次,洗滌液轉至容量瓶,加水定容。此溶液為樣品測定液。

式中:c 為標準曲線對應的多糖含量,mg/mL;n 為稀釋倍數;v 為培養基體積,mL;M 為培養基中海帶干粉質量,g。
1.3.4 海帶發酵液中巖藻糖含量的檢測
采用改良Gibbons 比色法[12]檢測巖藻糖。
標準曲線的制備:配置巖藻糖標準液,將20 mg 巖藻糖標準品加少量水溶解,溶液轉移至100 mL 容量瓶中。梯度配置配成1 mL 溶液置于比色管中,每個梯度3 個平行樣,冰水浴中加入 4.5 mL(體積比 6 ∶1)濃硫酸,搖勻。1 min 后,在沸水浴中準確加熱10 min,迅速冷制室溫(25 ℃左右),向每個管中加入100 μL 3%L-半胱氨酸,搖勻,靜置 90 min,分別在 396、427 nm 下測定吸光度。以吸光度值差為縱坐標,以樣品量為橫坐標,繪制標準曲線。樣品檢測以合適稀釋濃度樣品液1 mL 代替1 mL 巖藻糖溶液進行檢測。
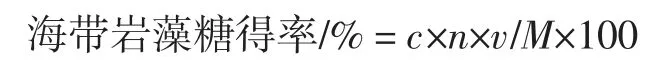
式中:c 為標準曲線對應的巖藻糖含量,mg/mL;n為稀釋倍數;v 為培養基體積,mL;M 為培養基中海帶干粉質量,g。
1.3.5 海帶發酵液中海藻酸含量的檢測
采用鈣凝-酸化法[13]檢測海藻酸含量。
向上清中加入4%的氯化鈣(氯化鈣用少量水溶解),攪拌狀態下逐滴加入,抽濾得沉淀為海藻酸鈣。向沉淀中加入等質量的水,攪拌均勻后,加入與氯化鈣等質量的鹽酸,抽濾,用50%的乙醇將沉淀沖洗兩遍,于烘箱中60 ℃過夜烘干至恒重,稱量烘干至恒重濾紙質量為M1,稱量濾紙及沉淀質量M2,M2-M1即為海藻酸的質量。
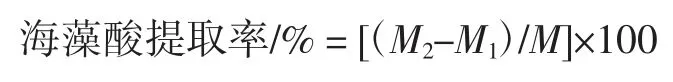
式中:M2為烘干至恒重的濾紙和海藻酸質量,g;M1為烘干至恒重的濾紙質量,g;M 為添加海帶干粉質量,g。
1.3.6 菌種鑒定及進化樹構建
經細菌試劑盒提取基因組,以提取所得DNA 為模板,利用細菌16SrDNA 通用引物27F(AGAGTTTGATCCTGGCTCAG)和 1492R(ACGGCTACCTTGTTACGACTT)擴增16s 序列,擴增后產物進行瓊脂糖凝膠電泳檢測,回收目的條帶送至華大基因測序。進化樹構建:在美國國立生物技術信息中心(National Center for Biotechnology Information,NCBI)采用基本局部比對搜索工具(basic local alignment search tool,BLAST)搜索與目的菌株序列同源性高的序列,經MEGA 6.0 構建系統發育樹。
1.3.7 統計分析
所有試驗重復3 次,結果表示為平均值±標準偏差(standard deviation,SD)。 使用 SPSS 軟件對數據進行方差分析(ANOVA)并使用繪制各菌株發酵后生物活性成分含量柱狀圖以顯示不同菌株對海帶的降解能力差異,篩選出特色益生菌株。
2 結果與討論
2.1 菌種篩選
經富集初篩和劃線純化,獲得20 株芽孢菌,后經黏度檢測篩選,獲得12 株對海帶有明顯降解效果的典型芽孢菌,海帶培養物的黏度及對應菌株形態描述詳見表1 和表2,菌落形態及細胞形態分別見圖1 和圖2。以此為出發菌株對海帶進行發酵降解和活性成分檢測。
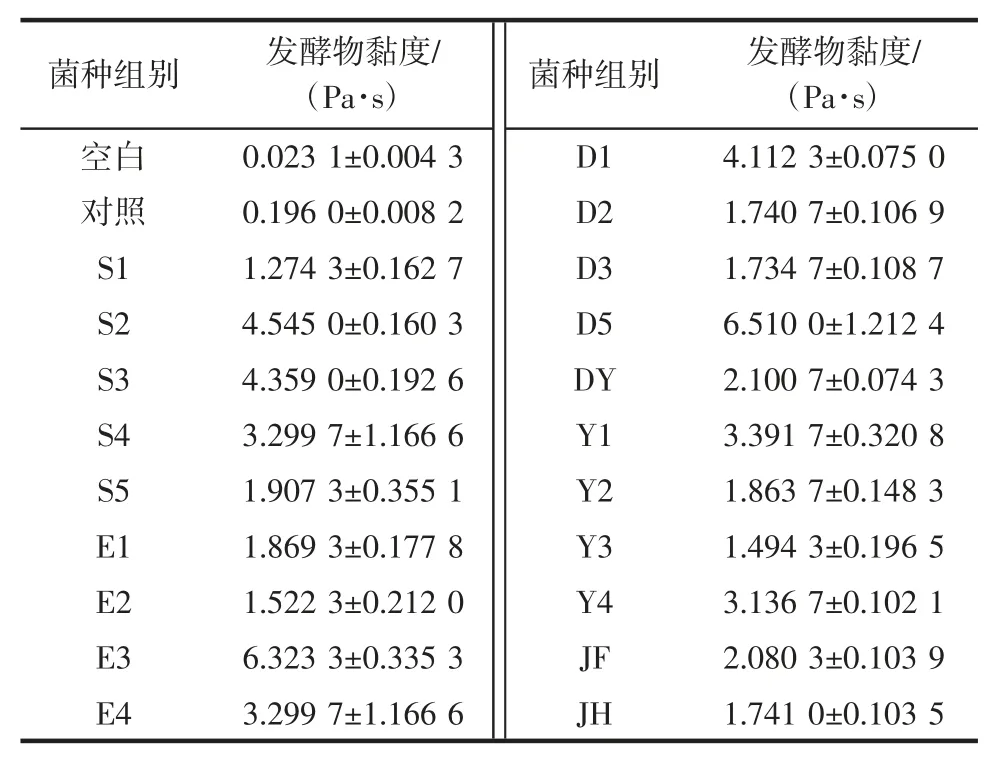
表1 初篩獲得20 株芽孢菌發酵海帶的黏度Table 1 Viscosity of Laminaria japonica fermented by 20 strains preliminary sreening

表2 12 株芽孢菌菌落及細胞形態描述Table 2 Colony and cell morphology description of 12 strains of Bacillus
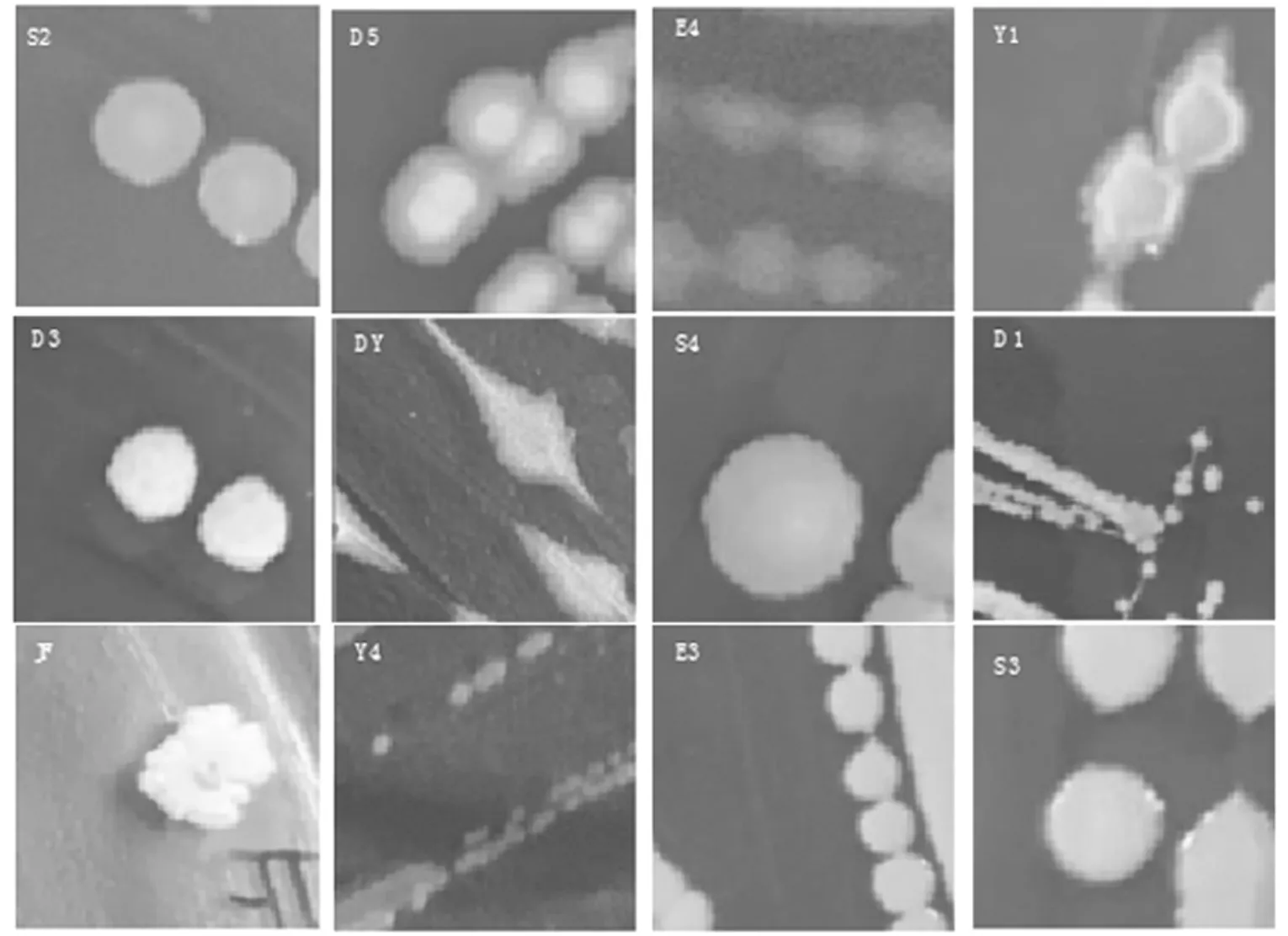
圖1 12 株芽孢菌菌落形態Fig.1 Colony morphology of 12 strains of Bacillus
2.2 海帶發酵液中多糖含量的檢測對比
采用苯酚硫酸法繪制多糖標準曲線:y=12.03x-0.051 6 R2=0.998 9。基于此曲線計算經不同來源菌株發酵后海帶多糖含量如圖3 所示。
結果表明獲得的不同來源菌種均可在發酵過程中增加發酵液中褐藻多糖的含量。菌株D3 在此過程中效果更為明顯,多糖含量為(360.89±6.93)mg/g,提取率達到(36.09±0.07)%,比對照組提高88.0%,高于熱水浸提法的海藻多糖提取率4.99%[14]和纖維素酶法褐藻多糖提取率11.62%[15]。
2.3 海帶發酵液中巖藻糖含量的檢測對比
采用Gibbons 比色法法繪制巖藻糖標準曲線:y=5.028 2x-0.032,R2=0.999。基于此曲線計算經不同來源菌株發酵后巖藻糖含量如圖4 所示。
數據顯示巖藻糖含量與對照組比較,提高16.4%~100.3 %。菌株D1 處理后巖藻糖含量為(138.76±21.11)mg/g,提取率為(13.88±0.21)%,比對照組提高一倍多;S2 與 D3 巖藻糖含量為(134.79±16.42)mg/g和(119.5±2.70)mg/g,提取率為(13.48±0.16)%和(11.95±0.03)%,較對照組增高94.5%和72.5%。結果均高于優化后熱水浸提巖藻糖的得率10.61%[16]。

圖2 12 株芽孢菌細胞形態Fig.2 Cell morphology of 12 strains of Bacillus
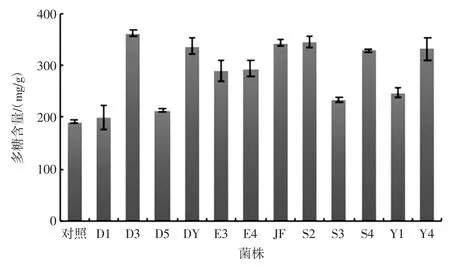
圖3 12 株益生菌發酵后海藻多糖含量Fig.3 The content of seaweed polysaccharide after fermentation of 12 probiotics
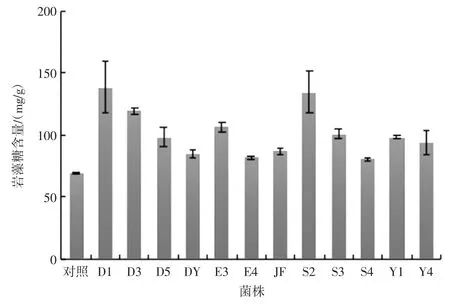
圖4 12 株益生菌發酵后巖藻糖含量Fig.4 The content of fucose after fermentation of 12 probiotics
2.4 海帶發酵液中海藻酸含量檢測對比
利用不同菌株對海帶進行發酵處理前后,海藻酸的產量變化明顯,菌株發酵后海藻酸含量如圖5 所示。
與對照組相比除菌株D5 外,其他菌株對海帶發酵處理后海藻酸產量明顯上升。其中,S2 發酵組提高最為顯著,經 S2 發酵后得率為(49.05±3.03)%,比對照組提高 9.40 倍;Y4 發酵組和 E4 發酵組分別為(38.61±1.79)%和(38.05±0.34)%,分別比對照組提高7.19 倍和7.07 倍。D3 發酵組提取率為(34.68±4.37)%,比未發酵的對照組提高6.36 倍。
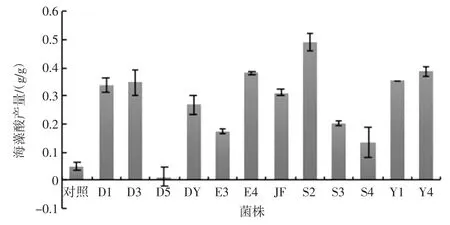
圖5 12 株益生菌發酵后海藻酸含量Fig.5 The content of alginic acid after fermentation of 12 probiotics
2.5 菌種鑒定結果及分析
綜合12 株典型芽孢菌株發酵后成分檢測分析,結果顯示D3 的對海帶的綜合降解提取效果最好。經擴增得到長度約為1 500 bp 的D3 菌株16S rDNA 序列,結果如圖6 所示。測序結果用BLAST 進行序列同源性比對,并利用MEGA 6.06 構建系統發育樹,結果見圖7。
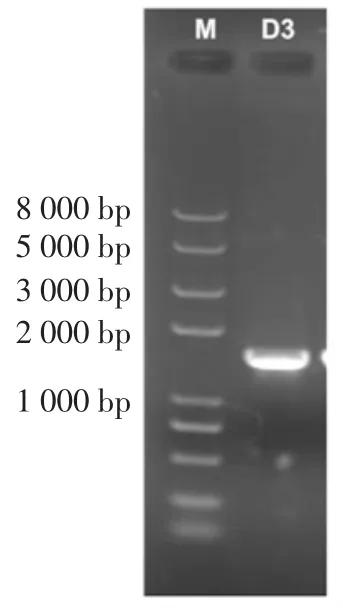
圖6 菌株D3 16S rDNA 經聚合酶鏈式反應(polymerase chain reaction,PCR)擴增結Fig.6 Electrophoresis of PCR amplified 16S rDNA
由系統發育樹可知,D3 菌株與Bacillus siamensis PD-A10(NR 117274)處于同一個分支,親緣關系最近,因此將菌株D3 歸屬于Bacillus siamensis。暹羅芽孢桿菌(Bacillus siamensis)是 2010年報道的一個新種,截至目前屬于該種的菌株報道不多。

圖7 菌株D3 基于16S rDNA 系統發育樹Fig.7 Phylogenetic tree of strain D3 based on 16S rDNA gene sequences
3 結論與討論
海帶發酵液多糖的主要成分是褐藻多糖,是海帶中廣泛存在的一大類具有生物活性的成分,在免疫調節、抗疲勞、抗凝血、抗輻射、抗氧化、抗衰老、抗腫瘤、抗病毒等方面的研究[17-18]不斷深入。人體產生的活性氧(reactive oxygen species,ROS)攻擊自身大分子化合物從而導致炎癥、糖尿病和癌癥等疾病,褐藻多糖具有良好的抗氧化活性,可以保護人體免受活性氧的損傷。低濃度的褐藻多糖能直接清除多形核白細胞呼吸爆發產生的活性氧自由基[19],高濃度的多糖能阻止活性氧的生成。在體外活性研究中巖藻多糖表現出對腫瘤細胞的殺傷力,抑制并有效控制腫瘤轉移。另外巖藻多糖的在抗流感、人乳頭瘤病毒(human papillomavirus,HPV)、單純皰疹病毒(herpes simplex virus,HSV)等的能力已得到驗證。小鼠實驗證明巖藻多糖可增強小鼠抗疲勞能力,增強耐缺氧能力,對受輻射損傷的的免疫功能調節作用呈量效關系。黎靜等研究發現海帶多糖可以調節皮膚基質金屬蛋白酶活性,進而調節光老化皮膚膠原蛋白的代謝,減少日光照射對皮膚的損傷[20]。L-巖藻糖可看做一種甲基戊糖,其較大量地存在于海藻及樹膠中,也發現于某些細菌的多糖中。L-巖藻糖具有免疫增強功能,并能夠降膽固醇、降血脂。一方面L-巖藻糖通過對腸道微生物的調整,增強腸道機能。另一方面,L-巖藻糖還可以與病毒、細菌、毒素等結合,防止其感染細胞,增強機體免疫力。肖瑤等實驗證明高劑量巖藻糖對免疫性肝損傷具有保護作用[21]。海藻酸極易形成鈉鹽,海藻酸鈉是一種高粘性的高分子化合物,具有優良的分散性、保濕性、成膜性、抗菌性、可生物降解、生物相容性好等特點[22],由于其極強的親水特性,當環境濕度較大時,海藻酸鈉可吸收外界水分,保持內部的相對濕潤,具有較強的吸濕作用,且成本較低,廣泛應用于食品保鮮。具有高古洛糖醛酸含量的藻酸鹽可形成堅固且剛性的凝膠,并且適用于食品和化妝品工業[6]。例如趙珊等以此為原料制備櫻桃保鮮劑,明顯延長保存期和貯藏品質[23]。陳妮娜等制備可降解改性復合膜應用與綠色食品包裝以代替傳統塑料膜[24]。機體產生的氧自由基使氧化反應增強,產生大量自由基,對機體組織的毒害很大[25]。海帶中的褐藻糖膠、多酚類物質,對清除人體內自由基保護機體具有極大的作用。數據表明經不同芽孢菌發酵,提高了海帶中抗氧化物質活性。而Bacillus siamensis D3 在其中具有顯著優勢,在食品領域具有重要的價值,應用潛力巨大。
微生物發酵是一種有前景的生物過程,通過增加發酵提取物中的酚類物質,蛋白質,還原糖和脂肪酸可提高海藻的營養價值及其生物功能活性[26]。對比各指標活性物質常用的提取方法及提取效率,區別于傳統酶法、水提法等方法對海藻中單一組分的提取及酸堿和高溫等處理過程,本研究中芽孢菌發酵能夠綜合性提高海藻中各項活性成分的種類和含量,綜合利用價值提高且經濟環保可循環。Hifney 等試驗證明海帶上清液中的海藻多糖、巖藻糖增多可能是菌株生長過程中分泌的胞外酶對海帶的降解作用[10],研究人員證實一些藻類真菌在生長中伴隨巖藻多糖酶和藻酸酶的產生,本研究以多指標評價篩選得到的降解海帶并提高其活性成分的芽孢菌,而試驗結果表明12 株不同芽孢菌株對海帶發酵后各項指標均有提高但是能力存在較大差異,推測是生長中伴隨產生降解海帶的生物酶的種類和活力不同導致的差異或者不同菌株發酵周期長短存在差異而導致的不同。具體促進海帶降解和釋放生物活性物質的酶類及具體降解機制有待進一步深入研究。
目前,食品工業中使用最廣泛的抗氧化劑是合成化合物,包括丁基化羥基甲苯,丁基羥基茴香醚,叔丁基氫醌和沒食子酸丙酯,可能會對健康造成不良影響和致癌性[27]。因此,本研究所篩選所得菌株提取海藻多糖和海藻酸鹽低成本、高效益、環保可再生,使海藻多糖和海藻酸鹽等有望作為天然、安全、可再生的抗氧化劑應用于各領域。另外,海帶發酵產物中富含的多糖、酚類化合物、氨基酸和脂肪酸等成分賦予海帶更高的營養保健功能,既可單獨提取或純化作為功能性食品、保健品和化妝品原料,使海帶活性物質作為新資源食品進一步開發,也可直接當作為復合活性營養應用于食品或化妝品中綜合利用。
