麥田雀麥發生動態及其對小麥產量的影響
2019-10-21 06:21:36李琦于金萍劉亦學白霜王金信
雜草學報 2019年1期
關鍵詞:產量
李琦 于金萍 劉亦學 白霜 王金信
摘要:雀麥(Bromus japonicus)是一種越冬性一年生雜草,是我國北方冬小麥田發生最嚴重的禾本科雜草之一,對小麥造成嚴重減產。為明確雀麥在我國冬小麥田的田間發生動態規律,于2013—2015年在山東省泰安市雀麥發生嚴重的冬小麥田進行相關試驗。采用固定樣方和隨機樣方取樣的方法,研究冬小麥田雀麥的出苗規律及其在田間的消長動態,同時研究不同密度雀麥對小麥產量的影響。結果表明,小麥播種后7~30 d為雀麥的出苗高峰期,至12月上旬出苗量占總出苗總量的85.3%。11月中旬,雀麥開始分蘗,平均分蘗數為4.8個/株,比小麥多1.3個/株。3月下旬雀麥生長速度加快,4月下旬平均株高超過小麥,5月中下旬平均株高達到 115.0 cm,高出同期小麥24.1 cm。5月中旬,雀麥與小麥單株平均鮮重趨于穩定,分別為17.24、37.72 g。雀麥與小麥在株高、鮮重等方面的變化趨勢基本一致,與外界溫度的變化密切相關。當雀麥密度為5株/m2時,小麥產量損失率達 5.35%,表明少數雀麥即可對小麥產量造成顯著減產;隨著雀麥密度的增加,小麥產量快速下降,其對小麥產量的影響主要表現在小麥的有效穗密度上,對小麥的千粒質量與穗粒數則影響較小。當雀麥密度為640株/m2時,小麥產量損失率為36.81%,小麥穗密度減少35.69%。
關鍵詞:雀麥;發生動態;溫度;小麥;產量
中圖分類號:S512.11;S451.1 ?文獻標志碼:A ?文章編號:1003-935X(2019)01-0010-07
Abstract:Japanese brome (Bromus japonicus) is a winter annual weed commonly found in wheat fields in China,especially in the northern region where it causes obvious yield losses. To determine the field dynamics of Japanese brome in wheat fields,experimentswere conducted from 2013 to 2015 in a field seriously infested in Taian. The dynamics of emergence,tillering,plantheight,fresh weight and its competitive effect on the yield of winter wheat was surveyed by both fixed and random quadrat sampling. The peak of Japanese brome seedling emergence occurred 7~30 d after wheat seeding. In early December,germinationrepresented 85.3% of the total. Tillering began in midNovember and was completed at the end of March of the following year. Japanese brome had on average 4.8 tillers per plant,1.3 more than wheat. In late March,Japanese brome speeded up growth and became taller than wheat after late April. Japanese brome reached 115.0 cm per plant on average in late May,24.1 cm taller than wheat. The average fresh weight of Japanese brome and wheat estabilized at 17.24 g and 37.72 g,respectively. The dynamics of height and fresh weight of Japanese brome were consistent with wheat,and were closely related to the changing temperature. When Japanese brome density is 5 spikes/m2,the yield losses of wheat are 5.35% indicating that a few Japanese brome could result in a significant reduction of grain yield. With the increase of Japanese bromedensity,wheat yield rapidly declined. Japanese brome had the largest effect on spike density factors in wheat yield with smaller impact on 1000-grain weight and grain number per spike. When the density reached 640/m2,the yield loss rate was 36.81% and spike density of wheat declined 35.69%.
Key words:Japanese brome;occurrence dymamic;temperature;wheat;yield
雀麥(Bromus japonicus)是一年生禾本科雀麥屬植物,起源于歐亞洲,常見于路邊、低洼濕地以及農田[1]。目前,雀麥廣泛分布于歐洲、北非、澳大利亞、美國、亞洲等地區[2]。近年來,由于氣候變暖,并且隨著國內耕作制度的改變及單一作用機制除草劑的長期使用,導致雀麥在我國多個省份發生嚴重,已成為我國冬小麥種植區發生危害嚴重的雜草之一[3-5]。1株雀麥平均結籽1 885粒,千粒質量約為3.4 g,極易隨風傳播[6-7],雀麥繁殖力強,傳播速度快,且與小麥爭奪光照、水分、養分、生存空間,嚴重影響小麥生長發育,降低其品質并造成減產,在雀麥發生嚴重的地區,小麥減產超過30%[3,8-9]。
趙祖英等研究了氟唑磺隆、啶磺草胺、甲基二磺隆、精喹禾靈、異丙隆、氟噻草胺、磺酰磺隆、丙苯磺隆、嘧啶肟草醚、苯唑草酮、吡氟酰草胺、炔草酯、唑啉草酯等藥劑對雀麥的防除效果,發現氟唑磺隆對雀麥的防效最高,且較為安全,為麥田防除雀麥的理想除草劑[10]。高興祥等明確了雀麥的最佳防除時間是冬前或冬后4月1日之前,越早防除越好[11]。譚金妮等利用MaxEnt模型對雀麥在我國的潛在分布區進行了預測,發現雀麥在我國的適生范圍涵蓋了除東北外的大部分平原地區,在我國冬小麥種植區呈快速蔓延趨勢[12]。
雜草生物學特性研究是所有雜草治理系統的基礎。目前,已有節節麥、播娘蒿、大穗看麥娘在田間發生動態及其對小麥產量影響的報道,為進一步的科學防除提供了理論依據[13-15]。但關于雀麥田間發生動態的相關研究尚未見報道。本研究選擇雀麥發生嚴重的冬小麥地區,采用固定樣方和隨機樣方取樣的方法,研究冬小麥田雀麥的出苗規律及其在田間的消長動態,同時研究不同密度雀麥對小麥產量的影響,旨在明確雀麥在田間的發生動態及生長規律,探究不同密度的雀麥對小麥產量的影響,為科學經濟地防除雀麥提供更加充分的理論依據。
1 材料與方法
1.1 試驗地點與時間
試驗于2013—2015年分別在山東省泰安市岱岳區大汶口鎮(35°57′N,117°3′E)、馬莊鎮(35°59′N,117°0′E)2個雀麥發生嚴重的冬小麥田進行,小麥于10月8日播種,品種為臨麥4號,上茬作物均為夏玉米,整個試驗過程中不使用除草劑,人工拔除其他雜草。
1.2 雀麥田間發生動態調查
選擇試驗地區雀麥發生嚴重的冬麥田,自小麥播種后開始,在每個試驗點選12個樣方插牌固定,樣方面積為0.25 m2(0.5 m×0.5 m)。每周1次拔除其中6個樣方內出現的雀麥,記錄拔除的雀麥株數;同時,每周對不拔除的另6個樣方記錄雀麥的株數。
1.3 田間雀麥與小麥的生物量測定
在田間自然條件下,自小麥出苗后,開始調查雀麥與小麥的生物量變化情況,試驗田對角線隨機取5點,每點各取20株雀麥和20株小麥,分別測量并記錄雀麥與小麥的株高、鮮重、分蘗數,此后每周調查1次,直至收獲,小麥越冬期未進行調查。
1.4 雀麥不同密度對小麥產量的影響試驗
在試驗田結合人工播種的方法,設置9個處理,雀麥密度分別為0、5、10、20、40、80、160、320、640 株/m2,每個處理重復4次,共36個小區,隨機區組排列,每個小區面積為3 m2(2.0 m×1.5 m)。試驗小區定期定苗,保證每個處理雀麥密度符合試驗要求。小麥收獲時,調查記錄小麥的有效穗密度、千粒質量、穗粒數、產量等指標,比較各處理間小麥產量,分析因雀麥危害對小麥造成的產量損失。
1.5 數據統計和計算方法
2年試驗趨勢一致,對2年試驗數據進行綜合平均分析。試驗數據采用SPSS 19.0軟件進行統計分析,用Duncans新復極差法進行差異顯著性檢驗,并用Sigmaplot 12.5軟件作圖。
2 結果與分析
2.1 雀麥田間發生動態
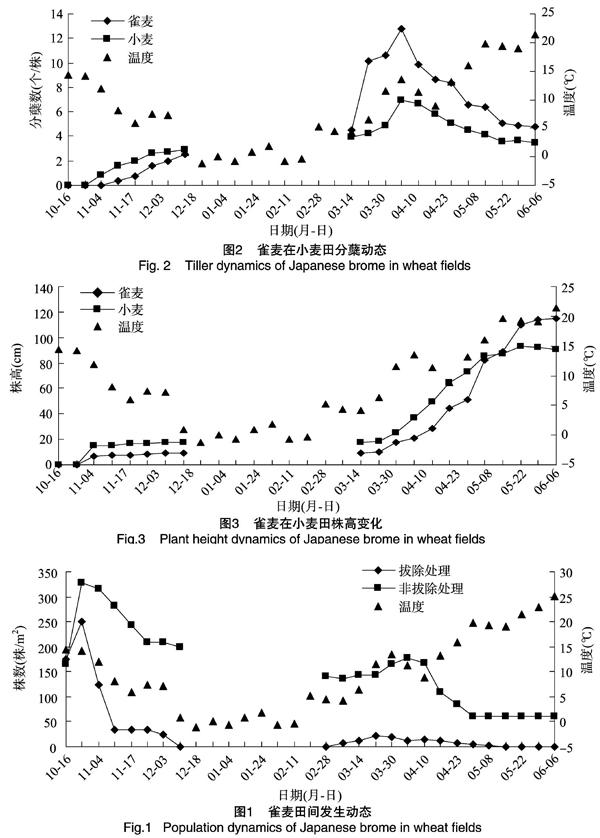
2.1.1 雀麥的出苗動態 冬小麥播種后1周左右,雀麥與小麥開始出苗。由圖1可知,通過雀麥每周拔除處理,10月中旬至11月上旬是雀麥出苗的高峰期,平均每周182.6株/m2,此時周平均氣溫在11.9~14.4 ℃之間。之后隨著氣溫逐漸降低,雀麥出苗量也逐漸減少。至12月上旬,周平均氣溫降至1 ℃以下,雀麥冬前出苗基本結束。翌年1月至2月中旬,周平均氣溫在-1.2~1.9 ℃ 之間,無雀麥出苗;2月下旬,氣溫開始升高,周平均氣溫升至5.2 ℃,雀麥有零星出苗;進入3月份,周平均氣溫在4.3~13.5 ℃之間,處理小區內有少量雀麥出苗;5月份之后,氣溫升高至19.0 ℃,幾乎無雀麥出苗。
從總出苗數可知,雀麥冬前出苗量占總出苗量的85.3%,春季出苗量僅占總出苗量的14.7%。在雀麥不拔除處理中,10月中下旬,雀麥出苗量達到最大值,也達到整個生長周期最大密度值328.7株/m2,至12月中旬,小麥越冬期前,雀麥出苗密度降至200.0株/m2。翌年2月下旬,小麥越冬期后,雀麥密度從200.0株/m2降至140.0株/m2。進入3月份,隨著氣溫升高,雀麥有少量出苗,雀麥株密度上升至176.7株/m2。進入4月份,雀麥與小麥均進入快速生長期,隨著雀麥種內競爭及與小麥的生長競爭,部分雀麥死亡,雀麥株密度由176.7株/m2降低至60.0株/m2。
2.1.2 雀麥的分蘗變化動態 由圖2可知,11月上旬,小麥先進入冬前分蘗期,中旬左右,雀麥開始分蘗;截至12月上旬氣溫降至1.0 ℃以下,冬前分蘗基本結束,雀麥、小麥平均分蘗數分別為2.6、2.9個/株,雀麥略低于小麥。翌年3月上旬,周平均氣溫升至4.3 ℃,雀麥單株分蘗數達到 4.5個/株,比冬前增加1.9個/株。之后隨著氣溫升高,雀麥與小麥進入春季分蘗高峰期,直至3月末分蘗數達到最大值,雀麥平均分蘗數為12.8個/株,比小麥多5.9個/株。此后,雀麥與小麥的分蘗數隨著營養生長的加速而逐漸消亡,直至5月下旬,二者分蘗數趨于穩定,雀麥為4.8個/株,小麥為 3.5個/株,雀麥略高于小麥。由結果可知,雀麥的分蘗能力要強于小麥,越冬前,雀麥分蘗數低于小麥,越冬后,隨著溫度的升高,分蘗數快速增長并超過小麥, 但二者分蘗趨勢一致,和外
界溫度的變化密切相關。
2.1.3 雀麥的株高變化動態 由圖3可知,在整個生長周期中,雀麥與小麥株高的變化趨勢大致相同,至12月上旬,雀麥與小麥平均株高分別達到9.2、17.3 cm;之后,周平均氣溫降至1.0 ℃以下,小麥進入越冬期,雀麥與小麥的株高均增長緩慢。3月上旬,周平均溫度最低達到4.2 ℃,并逐漸上升,此時雀麥與小麥株高分別為9.2、17.5 cm;3月下旬,周平均氣溫上升至13.5 ℃,雀麥與小麥的生長速度加快,此時雀麥、小麥株高分別為21.4、37.0 cm。5月上旬,隨著進一步的生長,雀麥的平均株高開始超過小麥,分別為88.7、87.4 cm。至5月中下旬,周平均氣溫達到 21.4 ℃,雀麥與小麥株高趨于穩定,雀麥平均株高達到115.0 cm,高出同時期小麥平均株高24.1 cm。
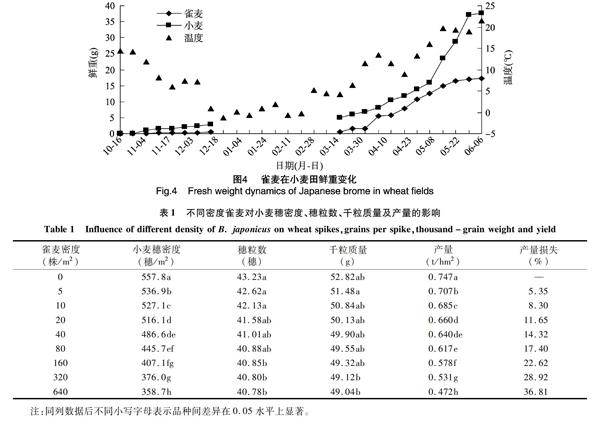
2.1.4 雀麥的鮮重變化動態 由圖4可知,從小麥開始出苗直至收獲,雀麥單株平均鮮重始終低于小麥單株平均鮮重;小麥越冬期前,雀麥與小麥的鮮重增長緩慢,至12月上旬,雀麥單株平均鮮重為0.41 g,小麥平均鮮重為 2.95 g。翌年3月下旬,周平均氣溫升至11.5 ℃,雀麥單株平均鮮重迅速增加,由1.57 g增長至5.59 g;5月上旬,小麥開始進入灌漿期,雀麥與小麥單株平均鮮重的差距進一步拉大。5月下旬,雀麥與小麥單株平均鮮重趨于穩定,分別為17.24、37.72 g,單株雀麥平均鮮重不足小麥的1/2。
2.2 不同密度雀麥對小麥產量及構成因素的影響
2.2.1 不同密度雀麥對小麥產量的影響 由表1可知,當雀麥株密度為5株/m2時,和無雀麥發生對照區相比,小麥產量已顯著降低,損失率達5.35%;當密度為160株/m2時,小麥產量損失率為22.62%;當達到最大密度640株/m2時,小麥產量損失率高達36.81%。對雀麥發生密度與小麥產量進行線性擬合,方程為y=0.480 0+
0.230 0e-0.006 2x,R2=0.948 0,說明小麥產量隨著雜草密度的增加而迅速降低,呈負相關關系。
2.2.2 不同密度雀麥對小麥穗密度、穗粒數和千粒質量的影響 由表1可知,雀麥株密度從0增加至640株/m2,小麥穗粒數從43.23穗減少至 40.78穗,減少5.67%。對雀麥發生密度與小麥穗粒數進行線性擬合,其線性擬合方程為y=40.910 0+2.390 0e-0.045 0x,R2=0.980 9;小麥千粒質量從52.82 g降至49.04 g,減少7.16%,線性擬合方程為y=49.720 0+2.890 0e-0.058 0x,R2=0.916 5,有小幅度的下降。
隨著雀麥發生密度的增加,小麥穗密度則顯著降低,當雀麥株密度為10株/m2,小麥穗密度較無雀麥發生對照顯著降低,減少了5.50%,當雀麥株密度為160株/m2,小麥穗密度減少了27.02%,其受影響程度與小麥產量的變化一致。對不同密度雀麥與小麥穗密度關系進行線性擬合,其線性擬合方程為y=3.740 0+1.550 0e-0.008 0x,R2=0.926 4。上述結果表明,雀麥對小麥產量造成影響時,降低小麥穗密度是主要途徑。
3 結論與討論
雀麥生長習性與小麥相近,但其繁殖力強,傳播速度快,能與小麥爭奪光照、水分、養分及生存空間,嚴重影響小麥的生長發育,降低其品質并造成減產。本研究明確了雀麥出苗動態以及分蘗、株高、鮮重的變化趨勢和規律;并系統研究了雀麥與小麥的生長變化,同時探究了雀麥不同密度對小麥產量的影響。
結果表明,小麥播種后1周至11月上旬為麥田雀麥的出苗高峰期。12月上旬,周平均氣溫降至1.0 ℃以下,雀麥冬前出苗基本結束,這段時間內雀麥出苗量占整個生長周期出草總量的 85.3%;翌年3月,隨著氣溫的升高,雀麥在田間表現出一個春季的出苗高峰期,該時期占總出苗量的14.7%。節節麥、大穗看麥娘、播娘蒿等一年生雜草在小麥田的出苗高峰期都集中在冬前,占到全年出苗總量的大多數[13-15],本研究結果與之大致相同。李美等認為黃淮海冬麥區,90%以上的雜草冬前都已出苗[16];高興祥等證明雀麥的最佳防除時期是冬前或冬后4月1日之前[11],本試驗也證明了以上結論。
雀麥在田間的分蘗能力強于小麥,冬前雀麥的分蘗數低于小麥,冬后雀麥與小麥進入分蘗高峰期,雀麥分蘗數快速超過小麥,之后分蘗隨著營養生長的加速而逐漸停止。田間條件下,小麥與禾本科雜草的密度都遵循“自疏法則”[17],其有效分蘗數與小麥播種量大小、雜草密度、水肥條件等都有關系[18]。而分蘗冗余可以減少外界不利環境對植株繁殖的影響[19]。雀麥與小麥在株高、鮮重等方面的變化趨勢基本一致,與外界溫度的變化密切相關。此外,每年10月中旬雀麥在冬麥田開始出苗,冬前生長緩慢,3月上旬進入返青期,開始快速生長,至5月上旬,株高由9.2 cm迅速增至109.7 cm,5月末植株成熟,該過程與Baskin等的研究結果[20]相一致。
魏仲塤認為小麥的穗數、穗粒數與千粒質量是衡量小麥產量的三大指標[21],本試驗探究了不同密度雀麥對這三大指標的影響。結果表明,雀麥對小麥產量的影響主要表現在小麥的有效穗密度上,對其千粒質量與穗粒數則影響較小。節節麥、大穗看麥娘、罔草對小麥產量的影響也主要是通過影響小麥的有效穗數來實現的[13,15,22],本研究結果與之一致。當雀麥株密度為5株/m2,小麥產量較對照處理顯著降低,減少了5.35%,表明在田間情況下,少數雀麥即可對小麥產量造成顯著減產。
農田雜草在光照、水分、養分、空間等資源方面與作物存在激烈競爭,其直接危害表現為作物產量損失。同時,該競爭也受氣候、土壤、耕作制度、水肥管理、雜草種群動態及雜草防治策略等因素影響[23-30]。因此,今后研究的重點是雀麥在不同耕作制度、不同小麥播種密度等條件下對小麥生長及產量的影響。
參考文獻:
[1]李揚漢. 中國雜草志[M]. 北京:中國農業出版社,1998:1180-1181.
[2]車晉滇,袁志強,金東紅,等. 雀麥生物學特性研究初報[J]. 北京農業,2010(36):41-43.
[3]魏 敏. 莊浪縣麥田雀麥生物學特性觀察[J]. 甘肅農業科技,2010(8):30-31.
[4]李秉華,王貴啟,蘇立軍,等. 防除雀麥除草劑的篩選及其對冬小麥安全性評價[J]. 雜草學報,2008,26(2):58-59.
[5]江彥軍. 石家莊市麥田雀麥和節節麥的發生特點及防治措施[J]. 河北農業科學,2010,14(8):106-106,147.
[6]王明發. 麥田雜草雀麥的發生、傳播與防除[J]. 雜草學報,1986,4(2):3-5.
[7]Li Q,Tan J N,Li W,et al. Effects of environmental factors on seed germination and emergence of Japanese Brome (Bromus japonicus)[J]. Weed Science,2014,63(3):641-646.
[8]馬建輝. 莊浪縣麥田雀麥發生特點及防治[J]. 甘肅農業科技,2010(12):49-50.
[9]李 琦,趙 寧,張樂樂,等. 黃淮海地區不同小麥品種對雀麥的化感作用[J]. 麥類作物學報,2016,36(8):1106-1112.
[10]趙祖英,郭文磊,李蓉榮,等. 除草劑對小麥田雀麥的生物活性測定[J]. 植物保護學報,2017,44(5):841-848.
[11]高興祥,李 美,白興勇,等. 雀麥不同防除時間對小麥產量的影響[J]. 植物保護學報,2018,45(2):347-351.
[12]譚金妮,李 琦,白 霜,等. 基于MaxEnt 的農田惡性雜草雀麥的潛在分布預測[J]. 中國農學通報,2016,32(23):49-54.
[13]房 鋒,張朝賢,黃紅娟,等. 麥田節節麥發生動態及其對小麥產量的影響[J]. 生態學報,2014,34(14):3917-3923.
[14]房 鋒,李 美,高興祥,等. 麥田播娘蒿發生動態及其對小麥產量構成因素的影響[J]. 中國農業科學,2015,48(13):2559-2568.
[15]房 鋒,李 美,高興祥,等. 冬小麥田大穗看麥娘種群動態及對小麥產量的影響[J]. 植物保護學報,2018,45(2):340-346.
[16]李 美,高興祥,李 健,等. 黃淮海冬小麥田雜草發生現狀、防除難點及防控技術[J]. 山東農業科學,2016,48(11):119-124.
[17]White J,Harper J L.Correlated changes in plant size and number in plant populations[J]. Journal of Ecology,1970,58(1):467-485.
[18]張永江,王文新,徐 燕,等. 小麥分蘗及成穗特性研究[J]. 現在農業科技,2008,494 (24):168-169.
[19]盛承發. 生長的冗余——作物對于蟲害超越補償作用的一種解釋[J]. 應用生態學報,1990,1(1):26-30.
[20]Baskin J M B C .Ecology of germination and flowering in the weedy winter annual grass Bromus japonicus[J]. Journal of Range Management,1981,34(5):369-372.
[21]魏仲塤. 不同播期生態條件下小麥穗粒數、穗粒重及千粒重性狀分析[J]. 耕作與栽培,1993,5(7):20-24.
[22]褚建君,李揚漢. 罔草對小麥的競爭臨界期[J]. 雜草學報,2003,21(1):21-23.
[23]張朝賢,胡祥恩,錢益新.雜草密度與作物產量損失的預測模型[J]. 植物保護,1997,23(2):6-10.
[24]吳云艷. 雜草稻競爭對栽培稻氮素積累量、氮素利用率及產量的影響[J]. 江蘇農業學報,2018,34(2):241-244.
[25]田欣欣,薄存瑤,李 麗,等. 耕作措施對冬小麥田雜草生物多樣性及產量的影響[J]. 生態學報,2011,31(10):2768-2775.
[26]冒宇翔,沈俊明,王曉琳,等. 不同耕作模式下麥田雜草發生規律[J]. 雜草學報,2018,36(3):5-12.
[27]蔣 敏,沈明星,沈新平,等. 長期不同施肥方式對麥田雜草群落的影響[J]. 生態學報,2014,34(7):1746-1756.
[28]王曉琳,張卓亞,伏 進,等. 秸稈還田條件下不同播種量結合除草劑對雜草和小麥生長的影響[J]. 江蘇農業學報,2017,33(2):307-313.
[29]李秉華,劉小民,許 賢,等. 玉米不同種植密度、耕作模式和水分管理對雜草的影響[J]. 雜草學報,2017,35(3):34-37.
[30]戴曉琴,歐陽竹,李運生. 耕作措施和施肥方式對麥田雜草密度和生物量的影響[J]. 生態學雜志,2011,30(2):234-240.
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
