短花針茅荒漠草原不同斑塊優勢種植物的C,N,P化學計量特征
2019-10-11 04:38:22于瑞鑫楊新國宋乃平
水土保持通報 2019年4期
于瑞鑫, 王 磊,3, 楊新國, 田 娜, 王 興, 宋乃平
(1.寧夏大學 西北土地退化與生態系統恢復省部共建國家重點實驗室培育基地,寧夏 銀川 750021; 2.寧夏大學 西北退化生態系統恢復與重建教育部重點實驗室,寧夏 銀川 750021; 3.南京大學 國際地球系統科學研究所, 江蘇 南京 210093)
化學元素對植物的生長以及生理機制調節有著極其必要的作用,而其中具備首要作用的化學元素是C,N,P[1]。C是植物體內各種生理過程的能量來源,N,P元素是植物蛋白質和核酸的重要組成元素,且是限制植物生長發育的兩個關鍵因子[2-3],而植物的C∶N和C∶P代表生物量與養分的比值關系,在一定程度上反映植物生長速度與養分利用效率的關系[4],植物葉片N∶P的臨界值則是判斷土壤對植物養分供應狀況的指標[5]。因此,對植物C,N,P化學計量特征的研究在其生長及營養分配等方面均具有重要意義。
生態化學計量成為研究不同尺度植被C,N,P含量及分布的有力工具[6]。He等[7-8]對不同生境、不同群落類型的中國草原植被的C∶N,N∶P化學計量比及其影響因子進行了分析,在大尺度研究草原群落的生態化學計量提供了依據。而小尺度上,生態化學計量特征隨時間的變化過程以與環境因子的響應關系一直是研究的熱點問題,牛得草等[9-10]以荒漠草原灌木植物及典型草原長芒草(Stipabungeana)葉片碳氮磷化學計量特征隨季節變化的響應規律,探討了不同發育階段C,N,P元素含量及其計量比的內穩性特征以及與生長速率的關系。王凱博等[11]分析了黃土丘陵區8種典型植物葉片化學計量隨時間變化的特征,研究了不同植物類型的生長速率與生長適應策略的關系及其差異。通過生長速率理論可知,植物通過調整它們的C∶N∶P比值以適應生長速率的改變,在生活史過程中通過調整生長速率以適應外界環境的變化[12]。而作為斑塊內優勢種,不僅通過生態化學計量的變化改變自身的物質分配,還進一步影響斑塊內植物的生態位策略;因此揭示優勢種的化學計量特征及其生活史策略,對于群落穩定性的評價具有重要的指示價值[13]。
荒漠草原短花針茅草原多以斑塊化鑲嵌的形式存在,受不利環境條件的影響,斑塊中優勢種的生活史策略及養分限制嚴重影響著斑塊化的群落穩定性及生長發育,因此在群落水平上深入研究優勢種的C,N,P的季節變化,對揭示群落的演替及變化過程具有十分重要的意義。本研究選取荒漠草原3種不同典型斑塊中優勢種為研究對象,探討不同斑塊中優勢種在不同發育階段的C,N,P含量的變化特征及元素計量關系之間的關系,試圖從生態化學計量的角度對群落養分的利用策略及生長過程進行初步探討,分析不同群落類型穩定性維持的生態化學計量依據。
1 研究區概況及研究方法
1.1 研究區概況
研究區地處寧夏吳忠市鹽池縣(37°04′—38°10′N,106°30′—107°47′E),總面積約6 744 km2,為典型的中溫帶大陸性氣候,是黃土高原向鄂爾多斯臺地、半干旱區向干旱區、干旱草原向荒漠草原、農業種植區向牧區過渡的4個過渡帶的重疊區[14],這種地理上的過渡性造成了自然資源多樣性和脆弱性的特點[15]。試驗區年均氣溫為8.4 ℃,年日照時數為2 863 h,年均無霜期為160 d,年均降水量250~350 mm,主要集中在7—9月,蒸發量高達2 139 mm,土壤主要以灰鈣土、風沙土、黑壚土為主,試驗區主要為發育在灰鈣土上的以短花針茅(Stipabreviflora)為建群種,小面積斑塊分布的豬毛蒿(Artemisiascoparia)、苦豆子(Sophoraalopecuroides)、牛枝子(Lespedezapotaninii)、老瓜頭(Cynanchumkomarovii)、草木樨狀黃耆(Astragalusmelilotoides),山苦荬(Ixeridiumchinense)等草本植物為主體[16]。
1.2 樣品采集與分析
根據資料收集及現場勘踏,在鹽池縣皖記溝村封育圍欄內選取具有代表性的群落結構斑塊33.34 hm2,確定3類典型斑塊:斑塊A(短花針茅)、斑塊B(短花針茅+草木樨狀黃耆)、斑塊C(豬毛蒿+苦豆子+老瓜頭),每處樣地隨機設置5個1 m ×1 m的樣方分別于2017年的6,7,8,9和10月,采用收割法獲取重要值大于20%的群落優勢種地上部分樣品,每種植物類型獲取5個重復樣品。
將采回物樣品清洗后裝入紙袋,于烘箱中105 ℃殺青0.5 h,然后75 ℃烘干至恒重。將烘干的植物樣品經磨碎過0.15 mm篩后測定植物C,N,P的含量。使用元素分析儀(Elemental Analyzer, Vario MACRO)測定植物體中的碳、氮含量,采用鉬銻抗比色法測定植物體中的磷含量[17]。每個樣品測定重復3次,計算出植物C,N,P含量平均值后,再計算植物C∶N,N∶P,C∶P值。
1.3 數據處理
采用Excel軟件對數據進行處理、整理和繪圖;用SPSS 20.0對C,N,P化學計量進行one-way ANOVA方法進行方差分析,均采用LSD方法進行多重比較;采用線性回歸分析法對不同斑塊C,N,P化學計量進行逐步回歸分析。
2 結果與分析
2.1 不同斑塊優勢種植物C,N,P含量季節變化
不同斑塊優勢種植物的C含量隨生長季的推移呈現不同變化特征(圖1)。斑塊A的優勢種短花針茅C含量受季節變化影響顯著(p<0.01),呈現先上升后緩慢下降,在生長季末期顯著上升的趨勢,10月C含量達43.93%。斑塊B的優勢種短花針茅隨時間的變化趨勢與斑塊A基本一致,但隨著生長季的變化不顯著(p>0.05),8月為生長季C含量最高時段;草木樨狀黃耆隨生長季變化總體呈逐漸上升的趨勢,生長季前期C含量顯著低于生長季后期(p<0.01)。斑塊C的優勢種呈現不同的變化趨勢(pKDZ<0.01,pLGT<0.01,p<0.05),老瓜頭隨生長季變化呈現先下降后上升的趨勢,6月達到最低42.05%;苦豆子則呈現逐漸上升的趨勢,9月達到最大45.21%;豬毛蒿呈現先上升后下降的趨勢,9月達到最大45.21%。
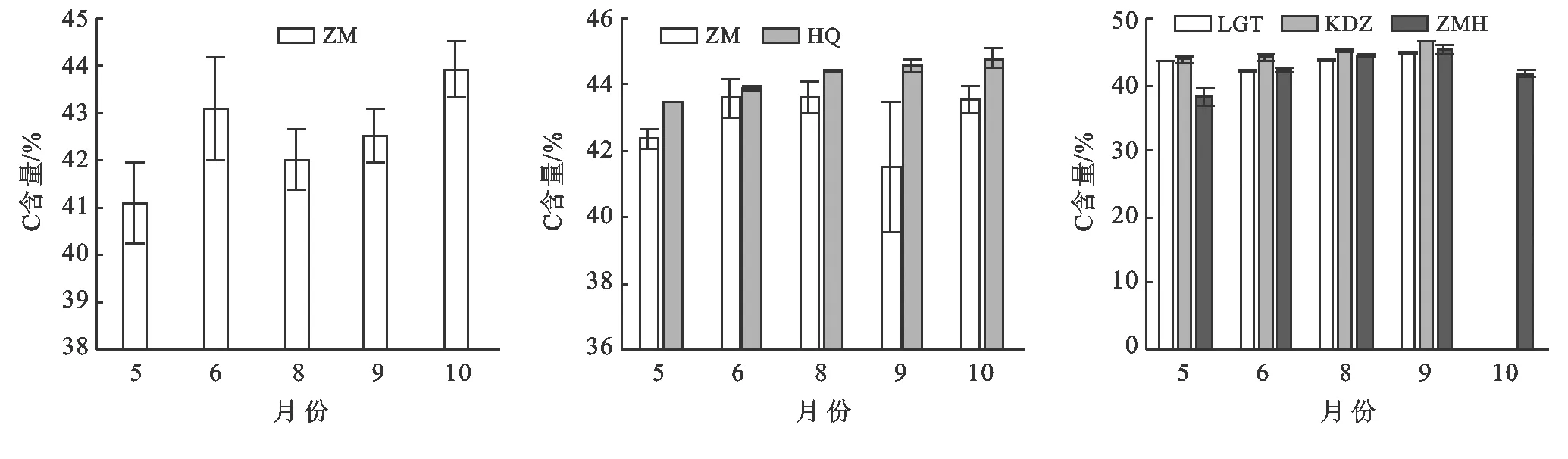
注:ZM:短花針茅;HQ:草木樨狀黃耆;LGT:老瓜頭;KDZ:苦豆子;ZMH:豬毛蒿。下同。圖1 不同斑塊優勢種C含量動態特征
不同斑塊優勢種植物的N含量隨生長季的推移呈現不同變化特征(圖2)。斑塊A優勢種短花針茅呈現先上升后下降,在生長季末期緩慢上升又略有下降的趨勢,6月達到最大(p<0.01)。斑塊B的優勢種短花針茅變化趨勢與斑塊A相同(p<0.05),6月N含量最高1.81%;草木樨狀黃耆N含量隨生長季變化呈現先上升后下降的趨勢(p<0.01),在6月達到最大3.385%后,開始下降繼而緩慢上升。斑塊C優勢種老瓜頭N含量5月最高后逐漸下降(p<0.01),生長季后期略有上升,但變化不顯著;苦豆子生長季內呈現逐漸下降的趨勢(p<0.01);豬毛蒿隨生長季變化呈先下降后上升繼而持續下降的趨勢,但受時間變化的影響不顯著(p>0.05)。
不同斑塊優勢種植物的P含量隨生長季的推移呈現不同變化特征(圖3)。斑塊A優勢種短花針茅隨生長季變化呈現先上升后下降生長季末期緩慢上升的趨勢(p<0.01),8月短花針茅的P含量最高為0.60%。斑塊B短花針茅在6月到第一個峰值0.24%,8月略有下降后開始顯著升高(p<0.01);草木樨狀黃耆的P含量隨生長季的推移顯著升高(p<0.01),10月含量最高0.71%。斑塊C中老瓜頭在生長季初期P含量顯著高于其他時段(p<0.01),然后迅下降到生長季最低0.23%,生長季后期緩慢上升;苦豆子在整個生長季范圍內呈顯著逐漸下降的趨勢(p<0.05),5月最高0.41%,9月最低0.17%;豬毛蒿P含量在生長季初期維持在較高的水平后顯著下降(p<0.01),8月為生長季內最低0.11%,生長季后期開始顯著上升并在10月P含量達到了生長季的最高值0.65%。
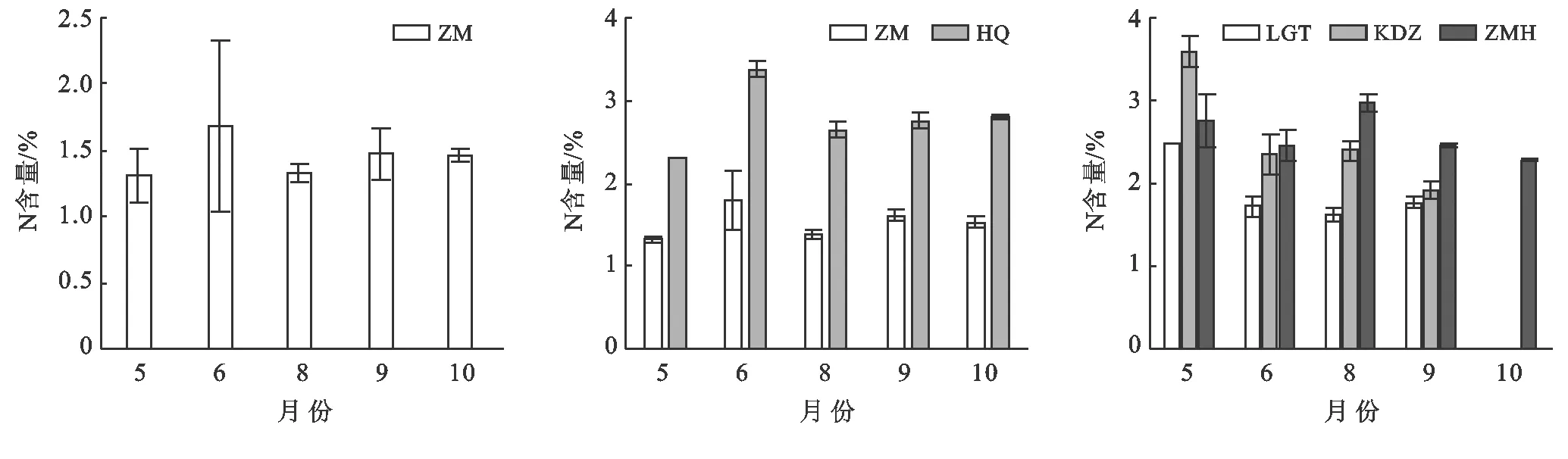
圖2 不同斑塊優勢種N含量動態特征
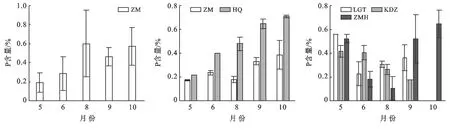
圖3 不同斑塊優勢種P含量動態特征
2.2 不同斑塊優勢種植物C,N,P計量比特征的季節變化
隨著植物生長季的延長,斑塊A內優勢種短花針茅的C∶N變化表現出先降低后上升,8月后再次下降的趨勢,但在生長季內變化不顯著(p>0.05),短花針茅的C∶P表現出持續降低的趨勢,5月和6月顯著高于其他月份(p<0.01),N∶P不同月份間表現出與C∶N相反的規律性,5—6月逐漸增加,6月達到最大值,然后顯著降低(p<0.01),9月略有升高后再次降低(圖4)。斑塊B內優勢種短花針茅C∶N的變化表現出先降低后上升,8月后再次下降的趨勢,且生長季內變化不顯著(p>0.05),短花針茅的C∶P表現出與C∶N相同的變化趨勢,5月和8月顯著高于其他月份(p<0.01),短花針茅的N∶P不同月份間表現出先上升后下降的趨勢,但生長季內變化不顯著(p>0.05);草木樨狀黃耆C∶N的變化呈先下降后上升的趨勢,5月到6月的變化顯著(p<0.01),C∶P,N∶P表現出不斷下降的趨勢,5月顯著高于其他月份(pC∶P<0.01,pN∶P<0.0.(1)(圖5)。斑塊C內優勢種老瓜頭C∶N的變化表現出先升高后降低的趨勢,5月顯著低于其他月份(p<0.01),老瓜頭C∶P,N∶P表現出相同的變化趨勢,都在9月達到了最高,分別為202.20,7.99;苦豆子C∶N的變化表現出先升高后降低再升高的趨勢,5月顯著低于其他月份,C∶P呈逐漸上升的趨勢N∶P不同月份間表現出先上升后下降的趨勢,但生長季內變化不顯著(pC∶P>0.05,pN∶P>0.05);苦豆子C∶N的變化呈先下降后上升的趨勢(p<0.01),9月達到最高值,N∶P則呈現出與C∶N相反的趨勢,生長季內變化不顯著(pN∶P>0.05);豬毛蒿生長季內的C∶N,C∶P,N∶P變化不顯著(pC∶N>0.05,pC∶P>0.05,pN∶P>0.05),C∶N呈現出先上升后下降再持續上升的趨勢,C∶P,N∶P呈現出先上升,在8月達到最高后,10月降到生長季最低(圖6)。
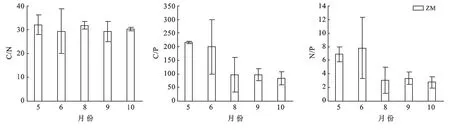
圖4 斑塊A優勢種C,N,P計量比特征
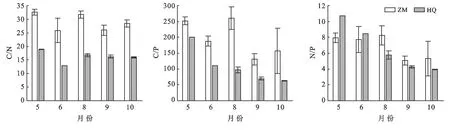
圖5 斑塊B優勢種C,N,P計量比特征
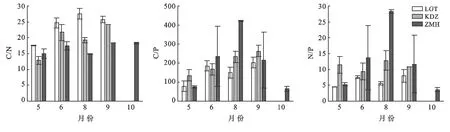
圖6 斑塊C優勢種C,N,P計量比特征
2.3 不同斑塊優勢種植物C,N,P化學計量的相關性
斑塊A優勢種植物C含量、N含量、P含量不相關(圖7);斑塊B優勢種植物C含量與N含量、P含量呈極顯著負相關,N含量與P含量呈極顯著正相關(圖8);斑塊C優勢種植物C含量與N含量、P含量顯著負相關,N含量與P含量不相關(圖9)。
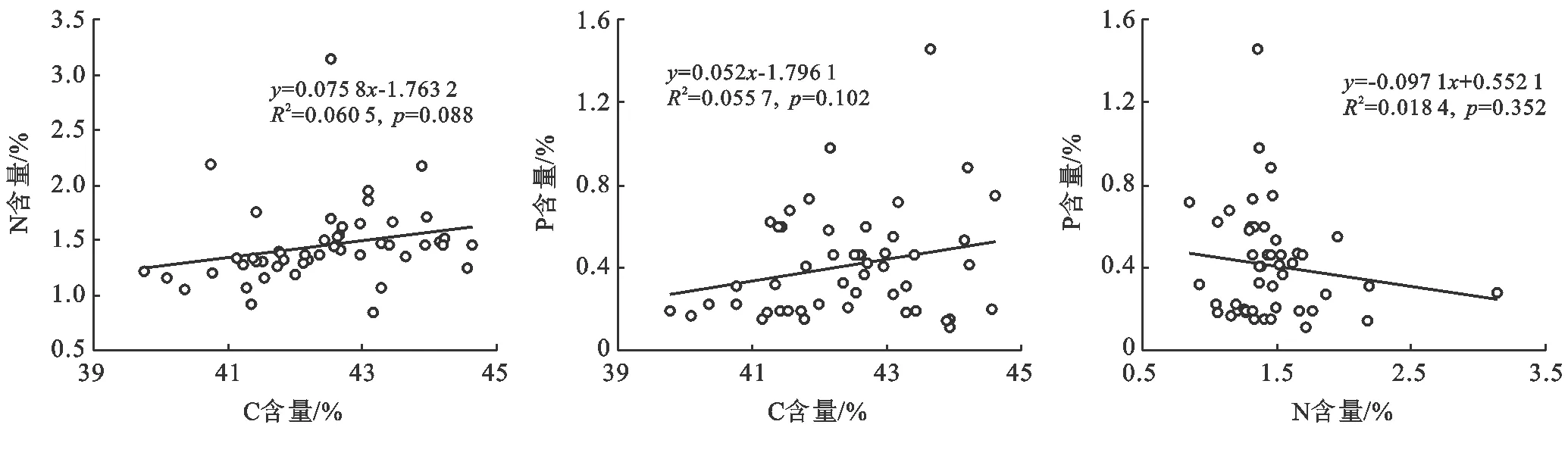
圖7 斑塊A優勢種C,N,P之間的關系(n=49)
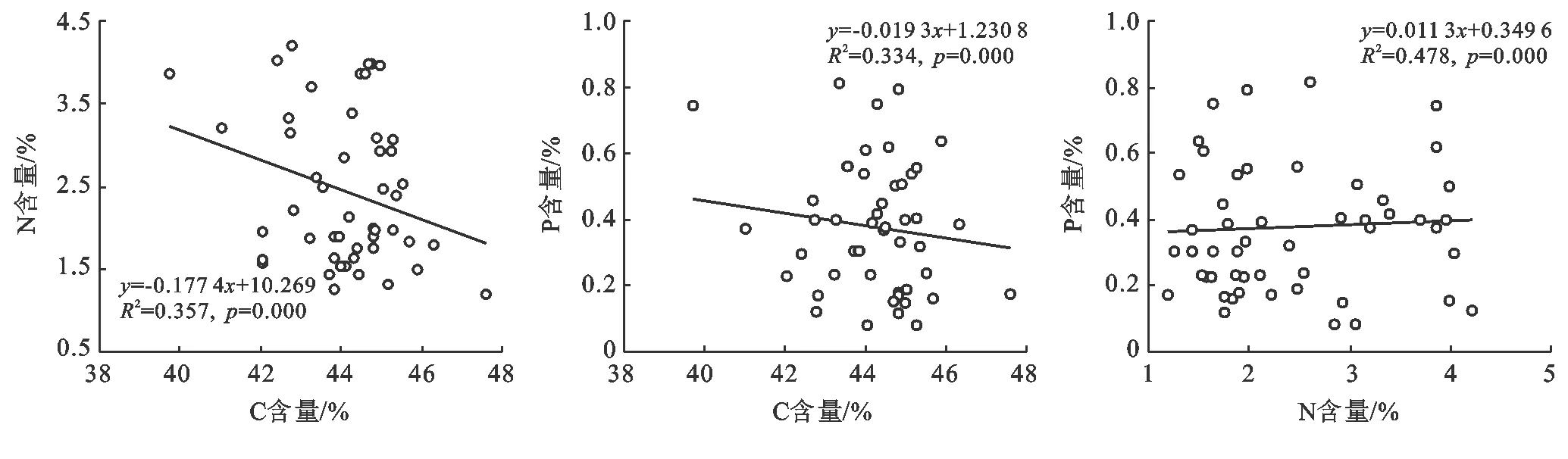
圖8 斑塊B優勢種C,N,P之間的關系(n=54)
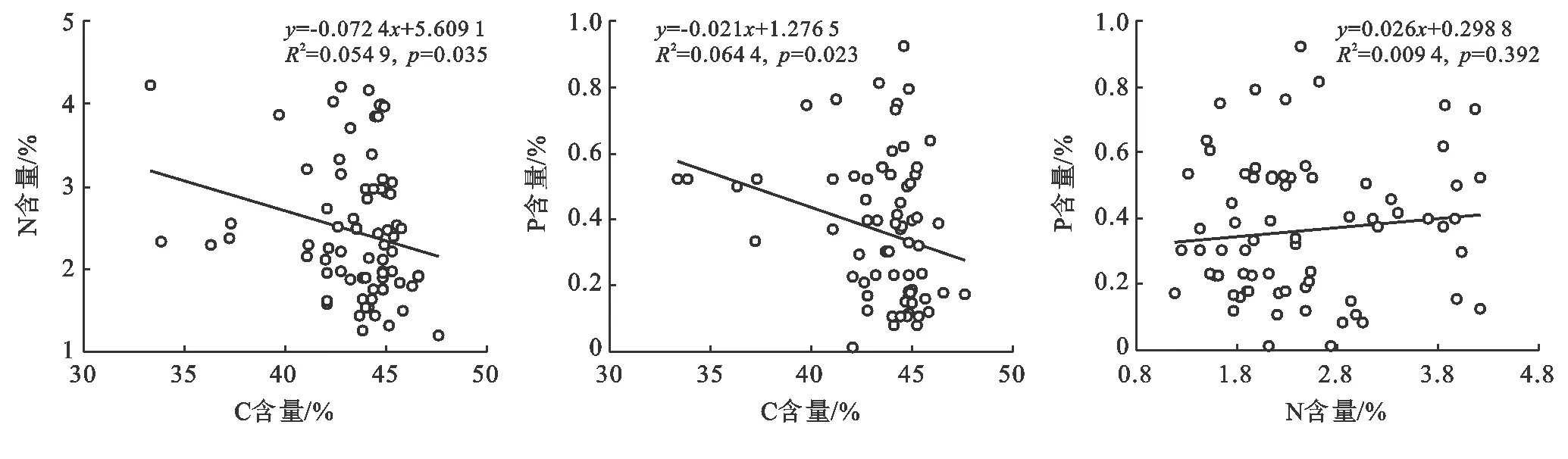
圖9 斑塊C優勢種C,N,P之間的關系(n=80)
3 討 論
N,P是限制陸地生態系統植物生長的主要元素[2,18-20],也是植物體內容易發生短缺的元素[21],植物體中的N∶P作為判斷環境對植物生長的養分供應狀況和植物的生長速率的重要指標[5],研究區域、生長階段、植物種類的不同,都會影響N∶P臨界值的變化[18]。
研究表明,當植物的N∶P<14時,群落水平上植物生長主要受N限制;N∶P>16時,植物生長主要受P限制;當14 植物葉片C含量與N含量、P含量的顯著負相關性以及葉N含量與P含量的正相關性,是高等陸生植物C,N,P化學計量的普遍特征之一,許多學者[21,25-26]已對不同地域植被化學計量特征進行研究,并驗證了這一規律,但斑塊A中短花針茅C,N,P含量之間無相關性,其原因可能是斑塊A中不能給優勢種群提供植物生長發育的穩定環境,而斑塊B,C內優勢種植物生長情況更穩定[26]。植物葉片C含量高則意味著其比葉重大,光合速率較低,生長速率慢,對外界不利環境的防御能力強,而N,P含量高,意味著其光合速率較高,生長速率快,對生長所需資源的競爭能力強[27-29]。 因此,群落優勢種C∶N∶P比值的季節變化,反映不同時段群落的生活史策略。不同斑塊的植物在生長季初期體內C含量基本都處在整個生長季的較低水平,斑塊A,B植物的N,P含量也基本處于相對較低的水平,斑塊C植物的N,P含量則處于較高水平,說明不同斑塊類型在生長季的初期對荒漠草原區的不利的氣候變化都具有較高的抵抗能力,與此同時,斑塊C中的苦豆子、豬毛蒿、老瓜頭能更早的適應環境的變化,早于短花針茅、草木樨狀黃耆進入的生長繁殖期,占據生長所需的資源。 5—6月水分的蒸騰速率及消耗開始加快,斑塊A,B的C,N,P含量都不同程度的開始上升,一段時間內保持在一個較高的水平。短花針茅與草木樨狀黃耆開始進入生長繁殖期,斑塊C優勢種C含量開始上升,N,P含量降低,生長速率降低,對資源的競爭能力開始減弱。斑塊A優勢種短花針茅的C,N含量在6—8月這個時段開始下降,P含量仍然保持上升趨勢,C∶P不斷下降,生長速率減弱,競爭能力減弱;斑塊B中短花針茅C含量則繼續保持相對較高的水平,N,P則開始下降,造成這種原因可能是由于短花針茅與草木樨狀黃耆之間相互作用的結果,使斑塊B此時對外界不良環境仍保持較高的抵御能力,在此過程中,短花針茅C∶P上升,草木樨狀黃耆C∶P下降,說明短花針茅的競爭能力高于草木樨狀黃耆;斑塊C中優勢種C含量上升、C∶P基本維持在一個叫高水平,具有較高的競爭能力。因此,處于生長旺盛期的植物群落,其優勢物種競爭能力與防御能力對群落的穩定性起著十分重要的作用。 隨著植物的成熟和衰老,斑塊A中短花針茅的C含量持續升高,對外界的抵御能力增強,9—10月C∶P小幅上升后降到最低,此時短花針茅開始出現返青現象,生長速率略有加快;斑塊B中草木樨狀黃耆C含量持續升高,C∶P下降,在整個生長季草木樨狀黃耆都保持著較高的防御能力,而短花針茅由于繁殖策略的改變,9月C含量、C∶P降到生長季的最低,生長速率減緩,競爭能力下降;斑塊C中苦豆子和老瓜頭在前期以較高的防御能力快速完成生長繁殖,占據生長空間,來適應貧瘠的土壤環境,這與崔東等[30]研究結果一致,并在9月完成生長周期,豬毛蒿在整個生長季對環境的適應性都保持在很高的水平,在8月生長旺盛期過后,生長速率逐漸減緩。 整體而言,5種優勢種植物的限制性生長主要元素為N元素,C,N,P含量存在一定相關性。以短花針茅為單優優勢種的群落采取的是防御性生活史策略。短花針茅+草木樨狀黃耆的群落則表現為競爭性策略。以豬毛蒿+苦豆子+老瓜頭為優勢種的群落,豬毛蒿表現為競爭性策略,苦豆子、老瓜頭表現為防御性策略。 (1) 不同類型斑塊C、N、P含量及C∶N∶P化學計量比在生長季內表現出不同的變化趨勢,優勢種植物的限制性生長主要元素為N元素,C,N,P含量存在一定的相關性。 (2) 不同斑塊的優勢種C、N、P化學計量存在顯著的季節差異,使其表現為不同的生活史策略,表明荒漠草原地區水土要素及其時空動態對植被的約束顯著,不同優勢種對外界因子產出了不同的適應策略。 (3) 荒漠草原干旱的環境不利于枯落物的分解,土壤有機質含量低,N元素成為該區域植物生長速率和養分供應狀況的主要限制因子,進而對不同群落的多樣性及穩定性產生影響。4 結 論
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
趣味(語文)(2020年3期)2020-07-27 01:42:46
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
