解淀粉芽孢桿菌G9R-3脂肽類化合物抑制香蕉枯萎病菌機理及防效評價
2019-10-08 09:01:30田丹丹李朝生覃柳燕劉挺燕黃素梅
西南農業學報 2019年8期
周 維,田丹丹,楊 揚,韋 弟,李朝生,覃柳燕,劉挺燕,黃素梅*
(1.廣西壯族自治區農業科學院生物技術研究所,香蕉抗病種質創制與病蟲害防控聯合實驗室,廣西 南寧 530007;2.中國熱帶農業科學院環境與植物保護研究所,農業部熱帶作物有害生物綜合治理重點實驗室,海南省熱帶農業有害生物監測與控制重點實驗室,海南省熱帶作物病蟲害生物防治工程技術研究中心,海南 海口 571101)
【研究意義】由尖孢鐮刀菌古巴專化型4號小種(Fusariumoxysporumf. sp.cubenserace 4,Foc4)引起的香蕉枯萎病被認為是目前全世界危害香蕉種植及產業發展的最嚴重病害。目前,大量研究實踐表明抗病品種結合生物防控已成為防控香蕉枯萎病最切實可行的策略及方法,其中開發利用植物內生菌是目前生物防控研究的熱點。已有研究表明內生菌與宿主能呈現互利共生關系,產生的活性物質能誘導或增強宿主植物對病蟲害及逆境脅迫的抗性[1]。解淀粉芽孢桿菌(Bacillusamyloliquefaciens)是目前生物防控應用較多的一類生防細菌,研究其生防機理發現其能產生多種抑菌物質,非核糖體途徑合成的脂肽類物質就是其中重要一類,主要包括表面活性素Surfactin、伊枯草菌素Iturin、泛革素Fengycin等[2-3];不同類型的脂肽類化合物具有不同的生物活性[4-5]。【前人研究進展】桑建偉等[2]研究發現脂肽類及聚酮類活性物質具有拮抗香蕉枯萎病FOC4活性。羅楚平等[6]和劉郵洲等[7]研究發現解淀粉芽孢桿菌產生的伊枯草菌素、泛革素具有較強的抗真菌活性,而表面活性素具有較強的抗細菌及溶血活性。Xiang等[8]研究發現泛革素及芽孢菌霉素對番茄枯萎病菌有較強的抑制活性。Deleu M等[9]研究認為芽孢桿菌產生的表面活性素雖無直接抗真菌活性但卻能增強伊枯草菌素的抗真菌能力,其能在植物根部表面形成生物膜,阻止病原菌的入侵。【本研究切入點】前期研究發現,G9R-3是一株內生解淀粉芽孢桿菌,該菌株對尖孢鐮刀菌Foc4有較強抑制作用,其能產生β-葡聚糖酶及揮發性抑菌物質(另文發表)。本研究擬對菌株G9R-3產生的脂肽類化合物類型及抑制Foc4活性進行分析,并開展盆栽防效研究。【擬解決的關鍵問題】探究G9R-3脂肽類化合物對 Foc4的抑菌作用機理并評價其防效,為香蕉枯萎病生防菌劑的研發利用提供理論基礎。
1 材料與方法
1.1 試驗材料
供試菌株G9R-3、香蕉枯萎病病原菌Foc4(Fusariumoxysporumf. sp.cubenserace 4)以及香蕉品種“桂蕉1號”香蕉組培盆栽苗均由廣西農業科學院生物技術研究所提供;供試培養基為LB、PDA[10]以及Landy培養基[11];供試儀器設備 Labconco 4.5 L型真空冷凍干燥機;熱電LYNX6000型高速冷凍離心機;基質輔助激光解吸電離-飛行時間質譜儀(Autoflex speedTMMALDI-TOF-MS, BrukerDalton,德國)。
1.2 試驗方法
1.2.1 大孔樹脂抽提法提取G9R-3脂肽類粗提物 參考桑建偉的大孔樹脂抽提法[2]提取G9R-3發酵液中的脂肽類化合物:接種內生細菌G9R-3種子液,使用500 mL Landy培養基進行搖瓶發酵, 30 ℃,180 r/min搖瓶培養 2 d,取發酵液8000 r/min 離心收集上清液。向上清液中加入15 mL XAD-16大孔樹脂,緩慢攪拌12 h后,濾紙片過濾。分別用超純水及無水甲醇洗脫2次,收集全部洗脫液用冷凍真空干燥機進行冷凍干燥,稱重后-80 ℃保存備用。
1.2.2 G9R-3 脂肽類化合物的種類分析 MALDI-TOF-MS具體方法參考Ongena等[12],乙腈中加入0.1 %三氟醋酸配制提取溶劑,α-氰-4-羥基肉桂酸(α-cyano-4-hydroxycinnamic acid)配制輔助基質溶液,將粗提物母液與提取溶劑、輔助溶液依照3∶1∶1比例混勻稀釋1000倍后檢測。質量掃描范圍為 900~2000 Da。
1.2.3 G9R-3脂肽類粗提物對Foc4 菌落生長的抑制作用 PDA培養基中加入脂肽類粗提物,制備終濃度分別為10、50、100、200、300 μg·mL-1的脂肽類化合物粗提物平板[2],平板中央接種直徑5 mm的Foc4菌餅,無毒 PDA 平板、無水甲醇分別作為空白對照及溶劑對照。每處理3重復,28 ℃培養7 d,十字交叉法測定菌落直徑并依據公式計算菌落生長抑制率:抑制率(%)=(對照菌落直徑-處理菌落直徑)/(對照菌落直徑-對照菌餅直徑)×100。
1.2.4 G9R-3脂肽類粗提物對Foc4孢子萌發的抑制作用 搖瓶發酵培養,過濾菌絲后梯度稀釋制備濃度為103CFU·mL-1的Foc4孢子懸浮液備用。帶毒平板制備方法同 1.2.3。每個平板上涂布100 μl制備的Foc4孢子懸浮液,無毒PDA平板、無水甲醇作為空白對照及溶劑對照。每處理3重復,28 ℃培養3 d,記錄孢子萌發數,并依據公式計算孢子萌發抑制率:抑制率(%)=(對照孢子萌發數-處理孢子萌發數)/對照孢子萌發數×100。
1.2.5 G9R-3 脂肽類粗提物對Foc4菌絲的影響 參照 1.2.3 的方法制備菌株G9R-3脂肽類粗提物帶毒平板,平板中央接種Foc4菌餅培養7 d,插片法觀察菌落邊緣菌絲,無毒PDA平板、無水甲醇作為空白對照及溶劑對照,0.04 %臺盼藍染色10 min后,觀察菌絲形態及染色情況。
1.2.6 G9R-3 脂肽類粗提物對香蕉枯萎病室內盆栽防效評價 依據1.2.1的方法,制備G9R-3發酵液及終濃度為1 mg·mL-1的脂肽類粗提物水溶液;按1.2.4方法,制備濃度為107CFU·mL-1的Foc4孢子懸浮液。選取株高一致,健康的“桂蕉1號”香蕉組培盆栽苗。采用灌根法分別接種上述菌株G9R-3發酵液及脂肽類粗提物水溶液,每株各澆灌20 mL,以清水為對照,每個處理30株。7 d后,每株傷根接種Foc4孢子懸浮液100 mL,60 d后調查植株發病情況。病情指數計算方法及病害分級標準參照農業行業標準(NY/T2248-2012),平均發病率、病情指數及防效(%)計算公式如下。
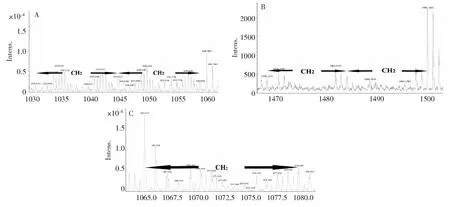
A:表面活性素; B:泛革素;C:伊枯草菌素A: Surfactin;B: Fengycin;C: Iturins圖1 MALDI-TOF-MS 分析菌株G9R-3脂肽類化合物類型Fig.1 Type of lipopeptide of strain G9R-3 analyzed by MALDI-TOF-MS
平均發病率(%)=各處理發病株數/調查總株數×100
病情指數 = {∑(各級病株數×該病級)}/(調查總株數×最高病級) ×100
防效(%)= (對照組病情指數-處理組病情指數)/對照組病情指數×100
2 結果與分析
2.1 G9R-3脂肽類粗提物的提取及脂肽類化合物的種類分析
使用500 mLG9R-3 Landy培養基發酵液,大孔樹脂抽提法提取脂肽類粗提物,冷凍干燥后稱重,經計算得到粗提物樣品濃度為442 mg/L。
結合MALDI-TOF-MS及LC-MS分析(圖 1),結果表明:菌株G9R-3脂肽類粗提物檢測到含有表面活性素的離子峰,質核比值m/z=1030.63、1044.65、1058.66 Da(圖1-A);含有泛革素的離子峰,質核比值m/z=1470.11、1484.01、1498.16Da(圖1-B);含有伊枯草菌素的離子峰,質核比值m/z=1065.53、1079.63 Da(圖 1-C)。綜合以上質譜分析結果,菌株G9R-3脂肽類化合物種類包括泛革素類群、表面活性素類群和伊枯草菌素類群。
2.2 G9R-3的脂肽類粗提物對Foc4菌絲生長的抑制作用
G9R-3脂肽類粗提物對Foc4菌絲的生長具有較強的平板抑制作用(圖 2、表1)。當終濃度為10 μg/mL時,粗提物對Foc4的菌絲生長產生抑制作用,隨著濃度增加,抑制作用逐步增強,當粗提物終濃度為300 μg/mL時,對Foc4菌絲生長抑制率可達58.91 %;對照溶劑甲醇無顯著抑制作用(P<0.05,下同)。
2.3 G9R-3的脂肽類粗提物對 Foc4 孢子萌發的抑制作用
試驗結果表明(圖3、表 2),G9R-3脂肽類粗提物能顯著抑制Foc4孢子在PDA平板上萌發。當終濃度為50 μg/mL時粗提物對Foc4的孢子萌發開始呈現明顯抑制作用,抑制率為19.47 %;而當粗提物終濃度為300 μg/mL時,對Foc4孢子萌發抑制率可達69.8 %。對照溶劑甲醇無顯著抑制作用。

CK1: 空白對照 (PDA);CK2: 溶劑對照(PDA+ 無水甲醇)CK1:PDA; CK2:PDA+ Anhydrous methanol圖2 不同濃度的G9R-3脂肽類化合物對Foc4菌絲生長的抑制作用Fig.2 Inhibitory effects of different concentrations of G9R-3 lipopeptides on the growth of Foc4 mycelia
2.4 G9R-3的脂肽類粗提物對Foc4菌絲形態的影響
采用臺盼藍染色法顯微鏡觀察。結果表明,G9R-3脂肽類粗提物能嚴重抑制影響Foc4菌絲形態。當粗提物終濃度為300 μg·mL-1時,Foc4菌絲腫大,打結扭曲,呈現明顯畸形。而空白對照和溶劑對照Foc4菌絲無顯著變化(圖4)。鏡檢觀察各濃度粗提物帶毒平板上生長的Foc4菌絲,經臺盼藍染色后,菌絲均被染成藍色,而相應的空白對照和溶劑對照菌絲未見被染色(圖 4)。表明G9R-3脂肽類粗提物能破壞Foc4菌絲細胞膜結構。
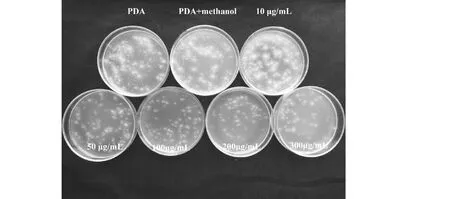
CK1: 空白對照 (PDA);CK2: 溶劑對照(PDA+ 無水甲醇)CK1:PDA;CK2:PDA+ Anhydrous methanol圖3 不同濃度的G9R-3脂肽類化合物對Foc4孢子萌發的抑制作用Fig.3 Inhibitory effect of different concentrations of G9R-3 lipopeptides on the germination of Foc4 spores

表2 不同濃度的G9R-3脂肽類化合物對Foc4孢子萌發抑制率

CK1: 空白對照 (PDA);CK2: 溶劑對照(PDA+ 無水甲醇)CK1:PDA;CK2:PDA+ Anhydrous methanol圖4 臺盼藍染色觀察不同濃度的G9R-3脂肽類化合物抑制Foc4菌絲形態Fig.4 The inhibitory effects of different concentrations of G9R-3 lipopeptides on morphology of Foc4 mycelia observed by trypan blue staining
2.5 G9R-3脂肽類粗提物對香蕉枯萎病的室內盆栽防效
盆栽防效試驗結果表明(表3),菌株G9R-3脂肽類粗提物(濃度為1 mg·mL-1)對香蕉枯萎病防效達76.53 %,平均發病率和病情指數分別為16.67 % 和12.58,而對照平均發病率高達93.33 %,防控效果明顯,差異達極顯著(P<0.01)。而脂肽類粗提物與發酵液防效差異不顯著(P>0.05),對比分析可知,脂肽類化合物的抑菌活性是內生解淀粉芽孢桿菌G9R-3防控香蕉枯萎病的重要生防機制,化合物的純化及可能產生的誘導植株抗性增加了防控效果。發酵液中活性物質相對濃度低,但菌株依靠自身拮抗病原活性能有效占據植株根際土壤生態位,降低了Foc4孢子濃度。
3 討 論
大量研究表明,解淀粉芽孢桿菌能夠促進植物生長及拮抗病原菌,因此通常被作為研究植物-微生物互作的模式菌株,其有8.5 %的基因組控制產生生物活性物質,諸如抗菌蛋白、脂肽類以及聚酮類物質[13-14]。脂肽類抗生素包括伊枯草菌素家簇 iturins、泛革素家簇 fengycins 和表面活性素家簇 surfactins[15-18]。除產生直接抗菌物質外,解淀粉芽孢桿菌還能誘導植株系統抗性(ISR),通常是提前激活植株茉莉酸及乙烯防御反應信號通路,從而有效的抵御外界病原菌及寄生菌[19]。本研究通過MALDI-TOF-MS和LC-MS分析發現菌株G9R-3主要能產生Surfactin、Fengycin 和Iturin 3類脂肽類化合物。活性分析發現G9R-3脂肽類化合物能夠抑制Foc4菌絲生長和孢子萌發,致畸菌絲并破壞細胞膜結構。這與國內外報道脂肽類物質Fengycin、Iturin抗真菌生物活性研究結果相似[2, 20-21]。Xiang 等報道 bacillomycin L對番茄枯萎病菌具有明顯的抑制作用,可導致細胞膜穿孔和活性氧積累; Surfactin具有抗細菌活性,其能改變細菌的表面疏水性,從而破壞生物膜完整性[22-24]。向亞萍等通過室內平板拮抗試驗發現Surfactin對番茄枯萎病菌生長的抑菌活性較弱,但其在防效評價試驗中表現出良好的生防潛力,這與Surfactins 具有較強的表面活性及在植株根部形成能夠抵御病原真菌侵入的生物膜有關[25-26]。Ongena等[27-28]還發現,surfactins 可以誘導大豆和番茄產生抗病性。本研究針對香蕉進行了枯萎病盆栽防效評價,結果表明菌株G9R-3脂肽類化合物能有效降低感病品種“桂蕉1號”枯萎病發病率及受病原侵染程度,分析可知菌株G9R-3抑菌活性是其防控香蕉枯萎病的重要機制,根據當前研究推測其產生的Fengycin、Iturin類物質抑制了 Foc4 的生長,降低了病原孢子濃度,同時推測其產生的surfactins類物質誘導了香蕉的抗病性并促進了菌株在植株根部的定殖,相關的誘導抗性機制有待進一步試驗證實。G9R-3是一株分離來自香蕉抗病品種“桂蕉9號”健康植株根部的內生菌,其產物是否參與香蕉協同抗病還不得而知,依據MALDI-TOF-MS分析還不能確定具體的效應物質,利用HPLC-MS進一步分離鑒定效應化合物,并探明其與香蕉協同抗病機制是今后研究的重點。

表3 G9R-3脂肽類化合物室內盆栽防效評價結果
注:不同字母表示差異達顯著水平(P<0.05)。
Note: Different lowercase letters represented significant difference at 0.05 levels.
4 結 論
香蕉內生解淀粉芽孢桿菌G9R-3能產生Surfactin、Fengycin 和Iturin 3類脂肽類化合物。其對香蕉枯萎病菌(Fusariumoxysporumf. sp.cubenserace 4,Foc4)具有較好的抑制活性。其抑菌機理為破壞Foc4 細胞膜并引起菌絲腫大和畸形,并抑制其孢子萌發。盆栽防效評價表明菌株G9R-3脂肽類粗提物(濃度為1 mg·mL-1)對香蕉枯萎病防效達76.53 %。
