黑米親本總黃酮與花色苷配合力及遺傳力分析
2019-10-08 09:10:06程祖鋅黃昕穎肖長春林荔輝鄭金貴
西南農業學報 2019年8期
關鍵詞:黃酮
程祖鋅,黃昕穎,肖長春,2,林荔輝,2,許 明,鄭金貴*
(1.福建農林大學農產品品質研究所,福建 福州 350002;2.福建農林大學作物科學學院作物遺傳育種與綜合利用教育部重點實驗室,福建 福州 350002)
【研究意義】與普通稻米相比,黑米具有較高的營養和保健價值[1-3],黑米的核心成分為花色苷——以黃酮核為基礎的一類物質,具有抗氧化、抗衰老、降血脂、抗炎以及抑制腫瘤生成等功能[4-6]。但不同黑米品種之間總黃酮、花色苷含量差異極顯著,含量高低不一[7]。選育高黃酮、高花色苷的黑米品種對提高黑米的營養和保健價值具有重要意義,雜種優勢利用是提高黑米功能成分性狀的有效途徑,配合力法是雜種優勢的主要測量方法。因此,研究黑米兩系雜交稻親本總黃酮、花色苷含量的配合力及遺傳力,對合理組配高總黃酮、高花色苷黑米兩系雜交稻組合具有重要意義。【前人研究進展】育種工作者越來越重視特種稻黑米良種的選育,已育成“晚秈紫寶”、“香黑米”等常規品種[8-9],也選育出了血黑A、香血粳A、186A等黑米水稻三系不育系[10],實現了雜種優勢利用,但關于稻米中總黃酮、花色苷含量的配合力及遺傳力的研究較少。伍時照等對2個黑米和2個白米的雜交后代進行研究,結果表明,水稻籽粒種皮的黑(紫)色素沉積是一個由顯性基因控制的質量性狀,受兩對獨立遺傳的互補基因控制[11]。陳廷文分析了水稻種皮黑色素沉積情況,結果發現,黑色素性狀表現受一對顯性色素基因控制,存在加性、顯性和上位性效應[12]。張名位等采用3個黑米水稻與1個白米水稻配制6個雜交組合,對黑米色素進行遺傳分析,結果表明,黑米水稻種皮色素含量的遺傳符合加性-顯性模型,顯性等位基因對提高色素含量起增效作用,隱性等位基因起減效作用,廣義遺傳力和狹義遺傳力均較高[13]。石幫志等對黑米與白米的雜交后代的米色表現進行研究,認為米色性狀為數量性狀,至少受兩對以上顯性重疊基因控制,表現有劑量效應[14]。楊海亮采用7×8不完全雙列雜交研究了特種紅米雜交稻花色苷含量的遺傳,結果發現,花色苷含量受親本一般配合力的影響,受不育系的影響大于恢復系,花色苷含量的廣義和狹義遺傳率均較高[15]。而王強通過對特種紅米雜交稻花色苷含量的遺傳分析,結果發現,花色苷的廣義遺傳力很大,狹義遺傳力卻很低[16]。王詩文以白米不育系為母本,色稻為父本,采用3×6不完全雙列雜交對雜交色稻糙米的花色苷含量進行分析,結果表明,雜交一代花色苷含量偏向于含量低的親本,母本中一般配合力為正的品種與黑米雜交,其特殊配合力為正值;而母本中一般配合力為負的親本與黑米雜交,其特殊配合力為負值[17]。孫明茂等報道,水稻花色苷含量是由多基因控制的數量性狀,受基因型和環境效應的共同影響[18]。【本研究切入點】本研究所選用的黑米不育系和恢復系,為新選育的黑米稻親本,株葉形態良好、糙米黝黑、米質優異,是組配黑米兩系雜交稻的重要種質資源。目前,尚無針對黑米兩系雜交稻親本的總黃酮、花色苷含量的配合力和遺傳力的系統研究,不育系和恢復系在黑米兩系雜交稻的總黃酮和花色苷含量性狀上所起的作用還有待進一步探明。【擬解決的關鍵問題】以6個不育系、5個恢復系及其配制的30個雜交組合為材料,通過不完全雙列雜交試驗,研究黑米兩系雜交稻總黃酮、花色苷含量性狀的遺傳規律,為我國高功能成分黑米兩系雜交稻親本選育、組合測配和評價鑒定等工作提供技術支撐。
1 材料與方法
1.1 試驗材料
2016年晚造在福建詔安,以新選育的4個黑米稻兩系不育系D18S、D38S、D40S、D43S和2個白米稻兩系不育系品272S、品48S為母本,5個黑米兩系恢復系R48、R92、R149、R156、R401為父本,按照不完全雙列雜交(NCII)設計配制30個黑米兩系雜交稻組合。以上材料由福建農林大學農產品品質研究所提供。
1.2 試驗方法
2016年11月20日30個黑米雜交組合及其親本播種于海南三亞,12月20日移栽。試驗按隨機區組排列,設3個重復,每個小區按8株×8行=64株插秧,株行距20 cm×20 cm,單本插秧,常規管理。每個小區取中間5株測定黑米糙米中總黃酮與花色苷含量,測定方法參考王詩文[17],以小區平均數為計算單位。根據NCII雜交模型進行配合力及遺傳力分析,數據分析采用DPS軟件分析[19]。
2 結果與分析
2.1 各性狀方差和配合力方差分析
供試組合間均方值差異達到極顯著水平(表1),說明供試組合間存在遺傳差異。進一步進行配合力分析表明,不育系對總黃酮、花色苷含量性狀的一般配合力方差均達到極顯著水平,表明所配組合的總黃酮和花色苷含量性狀中不育系起著重要的作用;恢復系對總黃酮含量性狀的一般配合力達到極顯著水平,表明在所配組合的總黃酮含量性狀中父本起著重要的作用。可見,供試組合的總黃酮、花色含量性狀受親本一般配合力和組合特殊配合力共同決定的,這2個性狀的一般配合力和組合特殊配合力的遺傳是由加性和非加性基因共同控制的。
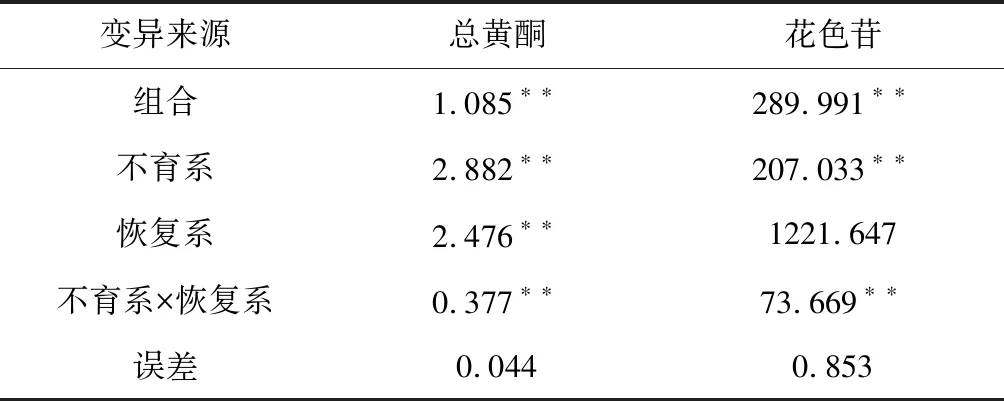
表1 30個組合方差分析及配合力方差分析
注:*、**分別表示P=0.05和P=0.01的顯著水平。

表2 親本各個性狀一般配合力效應值
2.2 親本的一般配合力效應分析
從表2可以看出,供試親本一般配合力(GCA)效應值在2個性狀間存在明顯差異,同一性狀的各個親本的GCA也存在明顯的差異,表明親本在不同的性狀上的加性效應大小不同,同一性狀受到親本的加性效應也不同。
4個黑米兩系不育系D18S、D38S、D40S、D43S在總黃酮和花色苷含量性狀上的GCA都有較大的正效應值,D38S的總黃酮和花色苷含量的GCA分別為12.548、51.780,D40S的總黃酮和花色苷含量的GCA分別為9.742、26.595, D38S、D40S在這2個性狀上的GCA都有很大的正效應值;黑米恢復系R48的總黃酮和花色苷含量的GCA分別為19.650、17.149,黑米恢復系R92的總黃酮和花色苷含量的GCA分別為2.390、14.225,黑米恢復系R48、R92在這2個性狀上GCA都有較大的正效應值。因此,D38S、D40S是總黃酮和花色苷含量性狀一般配合力最好的黑米兩系不育系,R48、R92是總黃酮和花色苷含量性狀一般配合力最好的黑米恢復系。
2.3 各性狀特殊配合力效應分析
從表3可以看出,同一性狀不同組合間的特殊配合力(SCA)也存在明顯差異。30個組合中,總黃酮、花色苷含量性狀的SCA變異幅度分別為-16.955~28.741、-39.106~58.746,組合D38S/R92的總黃酮、花色苷含量的SCA效應值分別高達21.675、35.283,組合D40S/R48的總黃酮、花色苷含量的SCA效應值分別高達28.741、58.746,這2個組合是花色苷、總黃酮含量性狀SCA效應得到最好的組合。雖然2個白米不育系的一般配合力都很低,但與黑米恢復系雜交,也能組配出特殊配合力較高的雜交組合,組合品48S/R149的特殊配合力比一些2個親本都是黑米的組合的特殊配合力都要高;2個都是黑米的親本也可能組配出特殊配合力低的雜交組合,但一般配合力都高的2個黑米親本所組配的雜交組合D38S/R92、D40S/R48具有更強的雜種優勢。因此,在育種中要特別重視一般配合力高的親本的選擇及其選配。

表3 各性狀特殊配合力效應值

表4 各性狀的基因型方差
注:δ1:母本GCA基因型方差;δ2:父本GCA基因型方差;δ12:父母本互作基因型方差;Vg:GCA基因型方差占總方差比例;Vs:SCA基因型方差占總方差比例;Vg1:母本GCA基因型方差占總方差比例;Vg2:父本GCA基因型方差占總方差比例。

表5 各性狀的遺傳力
2.4 各性狀群體方差和遺傳力估計
由表4可以看出,在總黃酮、花色苷含量的遺傳中一般配合力所占的比重大,分別為71.53 %、77.57 %(Vg>50 %),表明在這2個性狀中,親本的基因加性效應對雜種一代性狀的形成起主導作用。在總黃酮含量中,不育系的一般配合力基因型方差所占的比重為35.85 %,父本為35.67 %,兩者差異不大;在花色苷含量中,不育系的一般配合力基因型方差所占的比重為70.72 %比父本6.85 %大,說明在花色苷含量性狀上,不育系基因型方差對一般配合力方差貢獻更大,在育種過程中要特別注重不育系的選擇。
由表5可以看出,這2個性狀的狹義遺傳力大小順序為:花色苷>總黃酮。總黃酮含量、花色苷含量的遺傳力較高,說明在育種過程中,這2個性狀可在早代選擇以提高育種效率。
3 討 論
目前,未見有關稻米中總黃酮含量性狀配合力與遺傳力的研究,有關花色苷含量性狀配合力與遺傳力的報道也較少,未見黑米兩系雜交稻中關于這兩個性狀配合力及遺傳力的系統研究。
本研究供試組合的總黃酮、花色苷含量性狀由親本一般配合力和組合特殊配合力共同決定的,這兩個性狀的一般配合力和組合的特殊配合力的遺傳由加性和非加性基因共同控制的,這與楊海亮[15]、王強[16]等關于紅米中花色苷含量的研究結果相似。黑米花色苷含量性狀的遺傳中一般配合力所占的比重大,親本基因的加性效應對這兩個性狀的形成起主導作用,這與楊海亮研究結果相一致[15],而王強研究認為特殊配合力對紅米花色苷影響較大,親本的非加性效應對紅米花色苷含量性狀的形成起主導作用[16]。不同人研究結果不一樣,可能是由于研究的材料不同造成的。
分析表明,不育系總黃酮含量的一般配合力基因型方差所占的比重與父本相差不大,但不育系花色苷含量的一般配合力基因型方差所占的比重比父本大,不育系基因型方差對花色苷一般配合力方差貢獻更大,因此,在育種過程中,要注重不育系對花色苷含量的選擇[15]。廣義遺傳力反映了遺傳變異和環境變異的作用,狹義遺傳力度量加性遺傳效應[20]。總黃酮、花色苷含量的廣義遺傳力與狹義遺傳力都較高,說明這2個性狀的加性效應作用突出,受環境影響相對較小,因此,在育種過程中,這2個性狀可在早代選擇以提高育種效率[21]。
在水稻雜交育種中,性狀優良的親本,所配雜交組合的性狀未必優良;性狀一般的親本,所配雜交組合可能有很好的雜種優勢,這主要由于親本間的配合力差異引起的[22]。本研究也存在類似的情況,總黃酮、花色苷含量性狀一般配合力高的親本并不能保證其所配的雜種總黃酮、花色苷含量就高,但一般配合力都高的兩個親本,要獲得富含總黃酮、花色苷的雜交組合的概率要更高。
根據GCA和SCA,可以將親本分成四種類型:第一類,GCA高,SCA大,為最理想的親本;第二類,GCA高,SCA小,為較好的親本;第三類,GCA低,SCA大,為有一定的利用價值的親本;第四類,GCA低,SCA小,為基本無利用價值的親本[23]。綜合黑米不育系和恢復系的配合力表現,可以發現,黑米兩系不育系D38S、D40S,黑米恢復系R48、R92在這2個性狀的GCA都高,所配組合D38S/R92、D40S/R48的SCA也大,說明D38S、D40S、R48、R92是理想的富集總黃酮和花色苷的黑米兩系不育系和恢復系,在生產上有較大的應用前景,通過大量測交組配有望選育出農藝性狀、總黃酮和花色苷的特殊配合力好的即高產、高功能成分的強優勢組合。好的親本能夠聚集較多的有利基因,兩個都是好的親本進行組配,利用它們加性效應和非加性效應能夠配組出優勢強的黑米雜交稻新品種,因此,為了提高黑米雜交稻的育種效率,可以在一般配合力高的親本基礎上組配特殊配合力高的黑米雜交稻,同時進行廣泛的測交,這樣更容易獲得強優勢的黑米雜交稻組合。
4 結 論
本研究采用6×5不完全雙列雜交(NCII)模式對黑米的總黃酮和花色苷含量的性狀進行了配合力及遺傳力分析。結果表明,總黃酮、花色苷含量性狀存在加性效應和非加性效應,親本基因的加性效應對這2個性狀的形成起主導作用,這2個性狀遺傳力高,可在育種早期世代進行選擇以提高育種效率,D38S、D40S、R48、R92是富集總黃酮、花色苷的優異親本,應用前景較好。
猜你喜歡
四川蠶業(2021年2期)2021-03-09 03:15:32
四川蠶業(2021年3期)2021-02-12 02:38:46
中成藥(2018年11期)2018-11-24 02:57:00
中成藥(2017年8期)2017-11-22 03:19:40
中成藥(2017年10期)2017-11-16 00:50:13
中成藥(2017年4期)2017-05-17 06:09:50
哈爾濱醫藥(2016年1期)2017-01-15 13:43:16
天然產物研究與開發(2016年11期)2016-06-15 20:29:17
湖南師范大學自然科學學報(2015年1期)2015-02-27 14:50:04
安徽醫藥(2014年12期)2014-03-20 13:15:15
