馬氏珠母貝DNA甲基化轉移酶Dnmt1的克隆及表達
2019-09-25 11:17:32章佳斌盧曉文羅少杰鄧岳文
廣東海洋大學學報 2019年5期
關鍵詞:分析
章佳斌,盧曉文,羅少杰,焦 鈺,鄧岳文,2
馬氏珠母貝DNA甲基化轉移酶的克隆及表達
章佳斌1,盧曉文1,羅少杰1,焦 鈺1,鄧岳文1,2
(1. 廣東海洋大學水產學院,廣東 湛江 524088;2. 廣東省珍珠養殖與加工工程技術研究中心,廣東 湛江,524025)
對馬氏珠母貝()DNA甲基化轉移酶1(DNA methyltransferase 1,Dnmt1)的基因全長及組織表達作分析,探究馬氏珠母貝甲基轉移酶Dnmt1()參與貝殼礦化調控機理。利用RACE技術獲得的cDNA序列全長,并對的序列特征進行分析,同時利用逆轉錄-聚合酶鏈反應RT-PCR技術檢測馬氏珠母貝不同組織中的表達情況。基因cDNA長4 818 bp,其中,開放閱讀框為4 623 bp,編碼1 540個氨基酸。預測分子質量約為174.55 ku、理論等電點為5.89。結構域分析顯示,PmDnmt1具有DMAP 結合結構域、DNMT1-RFD結構域、鋅指結構、BAH結構域和C5胞嘧啶DNA甲基化酶結構域等。多序列比對結果發現,Dnmt1在不同物種間具有較高保守性。實時熒光定量PCR分析顯示在所有檢測組織中均有表達,在外套膜中央膜部分的表達量最高(< 0.05)。可通過調控外套膜的DNA甲基化修飾參與貝殼的形成。
馬氏珠母貝;DNA甲基化轉移酶;基因克隆;表達模式
DNA甲基化是目前在表觀遺傳學中研究最為成熟的一種表觀遺傳修飾,能夠引起DNA穩定性、構象、染色體結構等多方面的改變,在調控基因表達[1]、維持染色體結構[2]以及基因印跡[3]等方面扮演著重要角色。通常來說,DNA甲基化是由甲基轉移酶(DNA methyltransferase,Dnmt)起介導作用,并利用腺苷酰--甲硫氨酸(SAM)為甲基供體,將胞嘧啶(C)轉變為5-甲基胞嘧啶(5mC)[4]。哺乳動物中,3種DNA甲基轉移酶(Dnmt1、Dnmt2和Dnmt3)參與到DNA的甲基化修飾中。Dnmt3分為Dnmt3a和Dnmt3b兩種,能夠識別半甲基化和未甲基化序列,與DNA序列的從頭甲基化有關;而Dnmt1能夠以Dnmt3a/b修飾產物單鏈甲基化為模板,進而形成雙鏈甲基化,其主要功能是維持基因組已有的甲基化模式,在配子形成期和胚胎早期的甲基化模式的形成發揮重要作用[5-6]。
已有研究表明無脊椎動物基因組的甲基化修飾參與調控轉錄活性[7]、外顯子的選擇性剪接[8]和早期發育活動[9]等。近年來,貝類甲基化模式也受到關注。研究表明不同家系的馬氏珠母貝閉殼肌的甲基化水平與模式存在差異,近交家系的組織甲基化水平顯著高于雜交家系的組織甲基化水平[10]。馬氏珠母貝外套膜不同區域的甲基化存在顯著差異,其組織間的甲基化水平由高到底依次是中央膜>邊緣膜>套膜[11]。對牡蠣發育過程的甲基化研究發現[12],在牡蠣的胚胎發育和幼蟲變態過程中其基因組差異甲基化區域受到強有力的調控,羅少杰等[13]發現牡蠣在感染帕金蟲之后,其鰓與消化道的DNA甲基化水平顯著低于對照組,這表明DNA甲基化在貝類免疫中起到一定的作用。
近些年對貝類甲基化的研究逐漸深入,但貝類基因組甲基化形成與調控的具體機制還尚未闡明,對于在貝類基因組甲基化作用的研究也未見報道。因此,本研究通過首次克隆獲得馬氏珠母貝基因,并檢測其組織表達情況,以期為探究基因與貝類基因組甲基化關系,進一步探索無脊椎動物DNA甲基化修飾對基因表達調控作用提供參考。
1 材料與方法
1.1 材料
選用養殖于廣東省湛江市徐聞縣大井村海域(海域鹽度約為31,pH為8.2)的“海選1號”馬氏珠母貝()為實驗用貝,清除貝體表面附著物暫養2 d后取樣,樣品置于液氮中保存。RACE擴增所用材料為馬氏珠母貝的外套膜,用于熒光定量PCR的材料取自馬氏珠母貝外套膜的邊緣膜(Mantle edge,ME)、套膜(Mantle pallial,MP)、中央膜(Mantle central,MC)、閉殼肌(Adductor muscle,A)、鰓(Gill,Gi)、足(Foot,F)、肝胰臟(Hepatopancreas,He)、性腺(Gonads,Go)、血細胞(Hemocyte,B)。感受態細胞DH5α由本實驗室保存。
1.2 引物信息
見表1。
1.3 方法
1.3.1 總RNA提取和cDNA第一鏈的合成 將馬氏珠母貝各組織參照Trizol試劑說明書進行RNA的抽提;參照寶生物工程(大連)有限公司的Reverse Transcriptase M-MLV(RNase H-)說明書進行cDNA第一條鏈的合成。
1.3.2 DNA甲基化轉移酶基因的全長cDNA的獲得 根據馬氏珠母貝基因組數據[14],利用Primer Premier5.0 軟件設計引物,用于中間片段驗證和5′端和3′端的獲得。參照TaKaRa公司SMARTTM RACE cDNA Amptification Kit的說明書獲得用于RACE的cDNA模板。利用3′RACE 的巢式引物依次擴增3′末端序列,第1輪PCR反應體系:Premix TaqTM 5 μL,UPM引物0.4 μL,outer引物0.4 μL,cDNA第一鏈0.4 μL,ddH2O 3.8 μL;第2次PCR反應體系引物更換為NUP引物和inner引物。隨不同引物理論m值調整與其相應的退火溫度,PCR反應條件為94 ℃預變性 5 min;94 ℃變性 30 s,退火30 s,72 ℃延伸 3 min,35 個循環;72 ℃保溫10 min;4 ℃保存。5′末端序列擴增同樣同巢式PCR獲得,方法同3′末端序列。
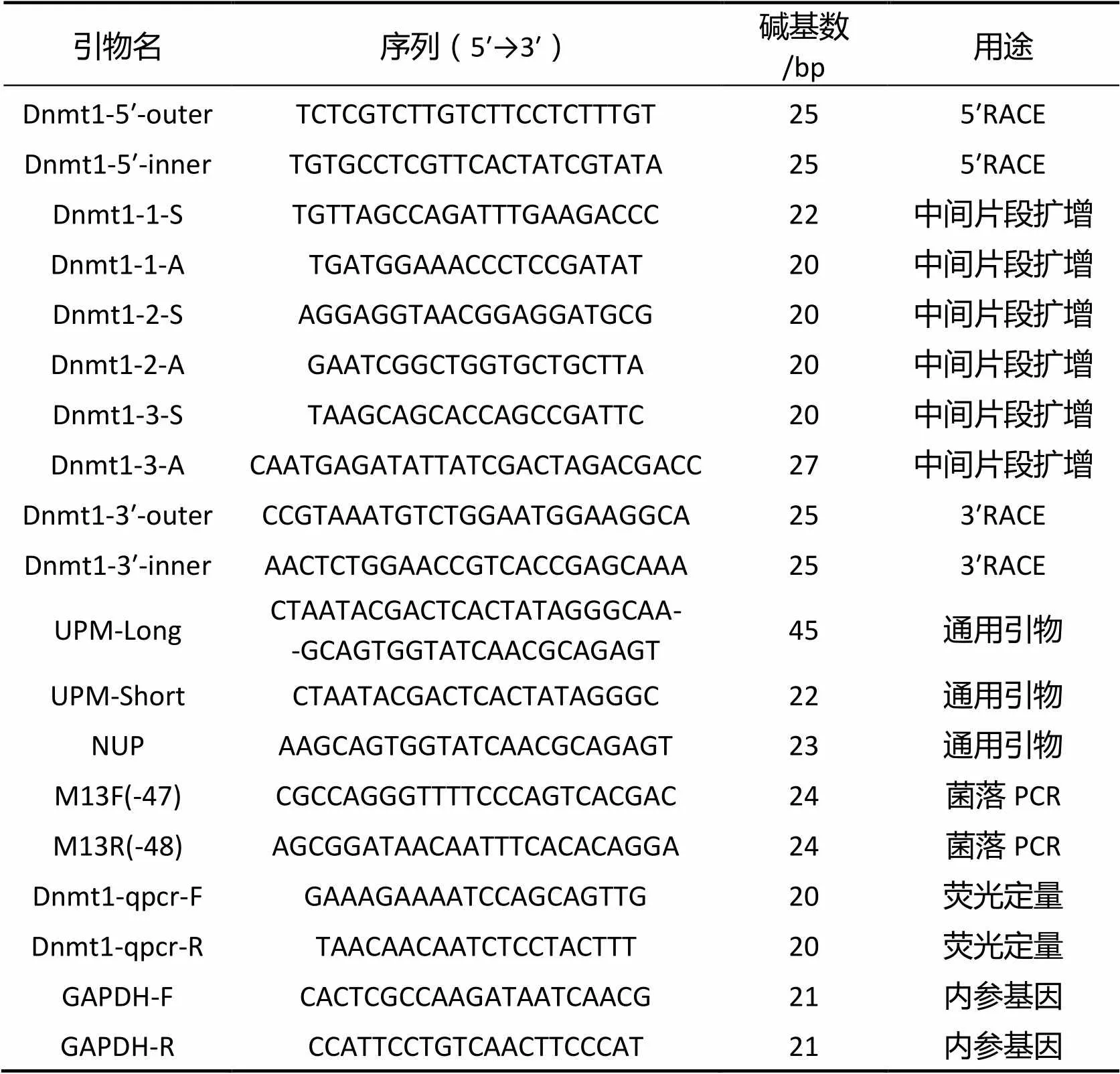
表1 研究所用引物
PCR產物先通過瓊脂糖凝膠電泳檢測產物,后根據純化回收試劑盒說明書純化目的基因片段;純化后的基因片段與PMD19-T載體連接后轉化到DH5α感受態細胞中,搖菌擴增后,經菌液PCR得到陽性單克隆,送至生工生物工程(上海)股份有限公司測序。
1.3.3 基因的生物信息學分析 利用DNAMAN軟件將的中間片段與5′端和3′端片段進行拼接,即獲得了其全長cDNA。根據所獲得的基因全長,使用在線BLAST網站對基因的同源性和相似性進行比對;使用在線網站工具ORF Finder預測基因的開放閱讀框;使用ExPASy分析蛋白的理化性質;使用PSITE V1分析氨基酸序列功能位點;使用SignalP 4.1和Cell-Ploc預測基因的信號肽與細胞定位;使用SMART和ScanProsite預測基因的結構域;使用SWISS-MODEL預測蛋白的空間結構;使用Prabi進行二級結構預測;使用EMBL在線工具Clustal Omega進行多序列比對;使用MEGAX軟件進行進化樹的構建。
1.3.4 熒光定量檢測組織差異表達分析 以GAPDH作為內參基因,采用2-ΔΔCT相對定量分析法,檢測在馬氏珠母貝各組織中的表達,反應體系和程序參照Life Technologies Corporation 公司的SYBR Select Master Mix 說明書。熒光定量PCR結果用ABI step one Software 分析,樣本重復性和組間差異性用SPSS19.0軟件分析。
2 結果與分析
2.1 基因的基本特征
2.1.1序列特征分析 使用RACE技術獲得基因序列(GenBank登錄號:MN159188),圖1可知,全長為4 818 bp,其中5′UTR為69 bp,3′UTR為126 bp,其中開放閱讀框4 623 bp,共編碼1 540個氨基酸。預測多肽分子質量174.54 ku、等電點為5.89。

起始密碼子(atg)和終止密碼子(tga)用小寫字母表示;下劃線部分代表DNMT1-RFD結構域;雙下劃線部分代表鋅指結構;波浪線部分代表BAH結構域;虛下劃線部分代表DMAP結合結構域;灰色背景部分表示C5胞嘧啶特異性DNA甲基化酶結構域;
2.1.2 PmDnmt1蛋白質理化性質分析 ExPASy預測帶負電荷的氨基酸殘基(Asp和Glu)為253個,帶正電荷的氨基酸殘基(Arg和Lys)為230個;不穩定系數為49.04,為不穩定蛋白;親水性平均系數為-0.763,即為親水性蛋白。通過PSITE V1對氨基酸序列的功能位點進行分析,發現該蛋白含有4個N-糖基化位點,3個cDNA和cGMP依賴蛋白激酶磷酸化位點,26個蛋白激酶C磷酸化位點,31個酪蛋白激酶Ⅱ磷酸化位點,9個酪氨酸激酶磷酸化位點,15個N-豆蔻酰化位點,2個酰胺化位點和23個微體C-端信號序列。SignalP 4.1分析顯示沒有信號肽,且通過Cell-Ploc分析發現該蛋白定位在細胞核。通過SMART和ScanProsite蛋白質分析工具和發現,該氨基酸序列在第10~57位上為DMAP結合結構域,第294~429位上為DNMT1-RFD結構域,第523~569位上為鋅指結構(CXXC-type),第640~766位和821~989均為BAH結構域,第1 034~1 488位上為C5胞嘧啶特異性DNA甲基化酶結構域(圖2)。在Prabi在線網站上對基因進行二級結構預測,結果顯示:α螺旋30.97 %、β折疊4.61 %、延伸鏈15.06 %、無規則卷曲49.35 %。采用SWISS-MODEL軟件預測PmDnmt1蛋白質分子的三維結構,與太平洋牡蠣Dnmt1結構存在較高的相似性(圖3)。
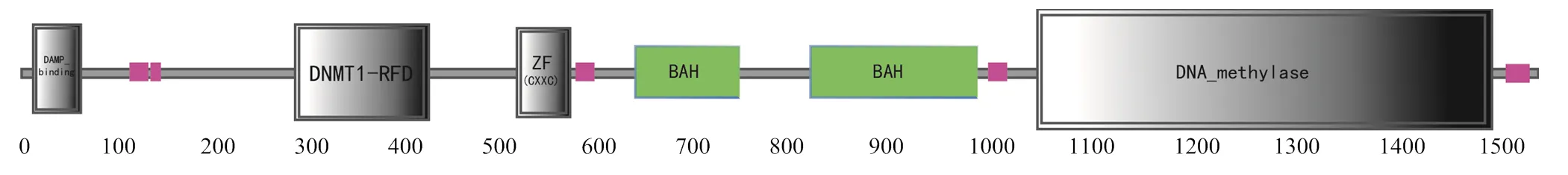
圖2 PmDnmt1結構域示意圖
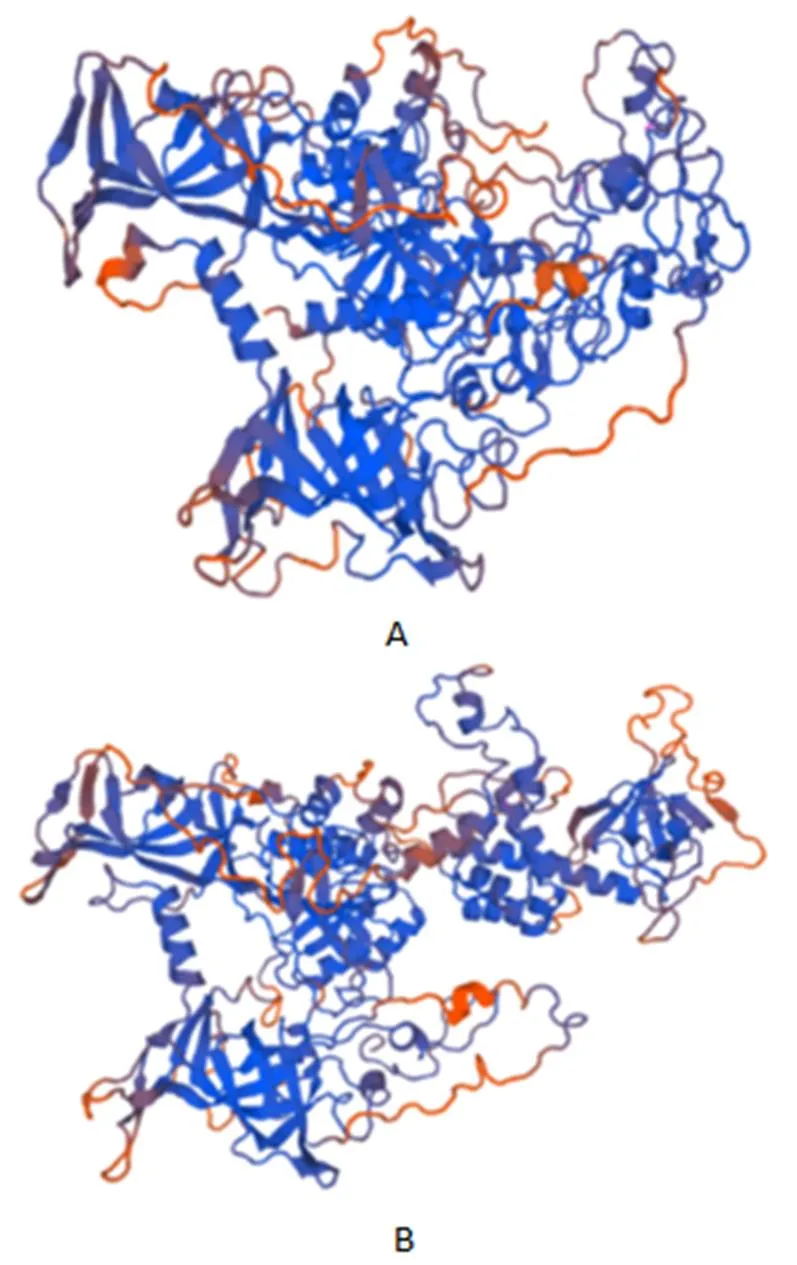
A:馬氏珠母貝;B:太平洋牡蠣
2.1.3 PmDnmt1與其他物種的同源性分析 以EMBL在線工具Clustal Omega將PmDnmt1與太平洋牡()、美洲牡蠣()、蝦夷扇貝()、擬球海膽()密西西比鱷()的Dnmt1的氨基酸進行多序列比對分析,結構顯示物種間的Dnmt1具有較高的相似性(圖4)。以馬氏珠母貝、太平洋牡蠣、蝦夷扇貝、擬球海膽、家蠶(、斑馬魚()、密西西比鱷、綠海龜()、褐家鼠()和人()的Dnmt1氨基酸序列為基礎,使用NJ法構建系統進化樹。結果顯示,馬氏珠母貝和太平洋牡蠣聚為一簇,并與蝦夷扇貝聚在同一分支上;人和褐家鼠聚為一簇,并于密西西比鱷、綠海龜和斑馬魚聚在同一分支上(圖5)。
2.2 組織表達差異分析
應用實時熒光定量PCR技術分析在組織中的表達量,所用組織分別為:外套膜的邊緣膜(ME)、套膜(MP)、中央膜(MC)、閉殼肌(A)、鰓(Gi)、足(F)、肝胰臟(He)、性腺(Go)、血細胞(B)等九個組織。實驗結果表明在馬氏珠母貝的各個組織中均有表達,其中在中央膜上的表達量最高(< 0.05),在血淋巴中表達量最低(< 0.05),在余下7個組織中表達水平較為一致(如圖6)。
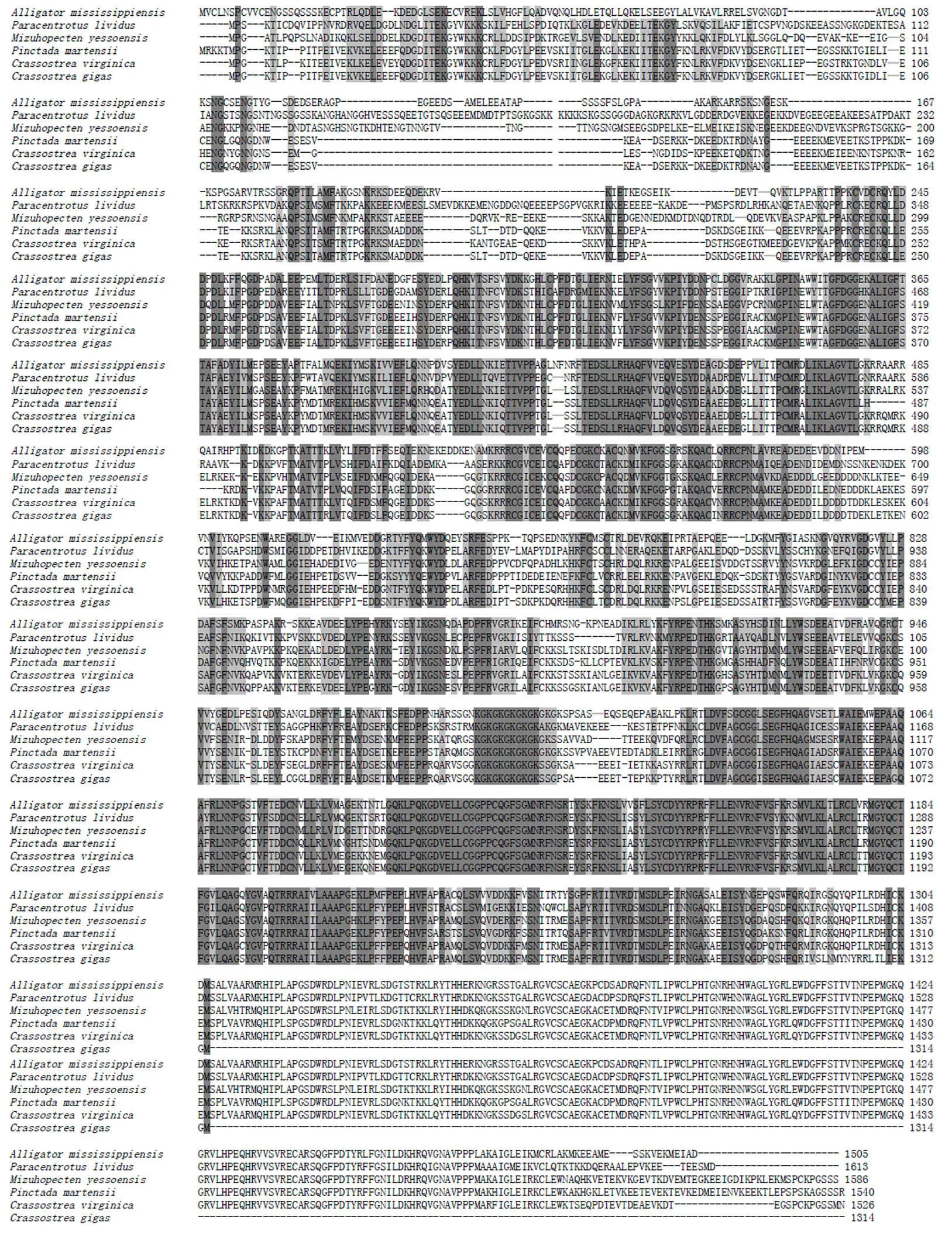
保守氨基酸序列以藍色陰影標準
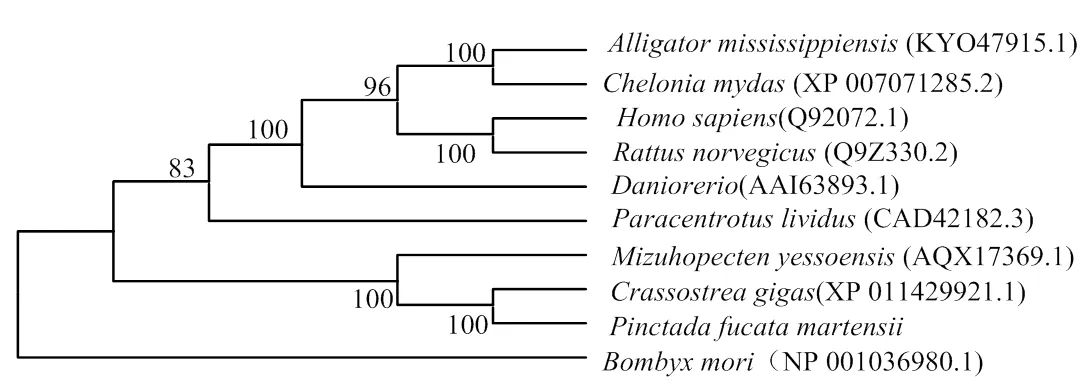
圖5 PmDnmt1氨基酸序列聚類分析
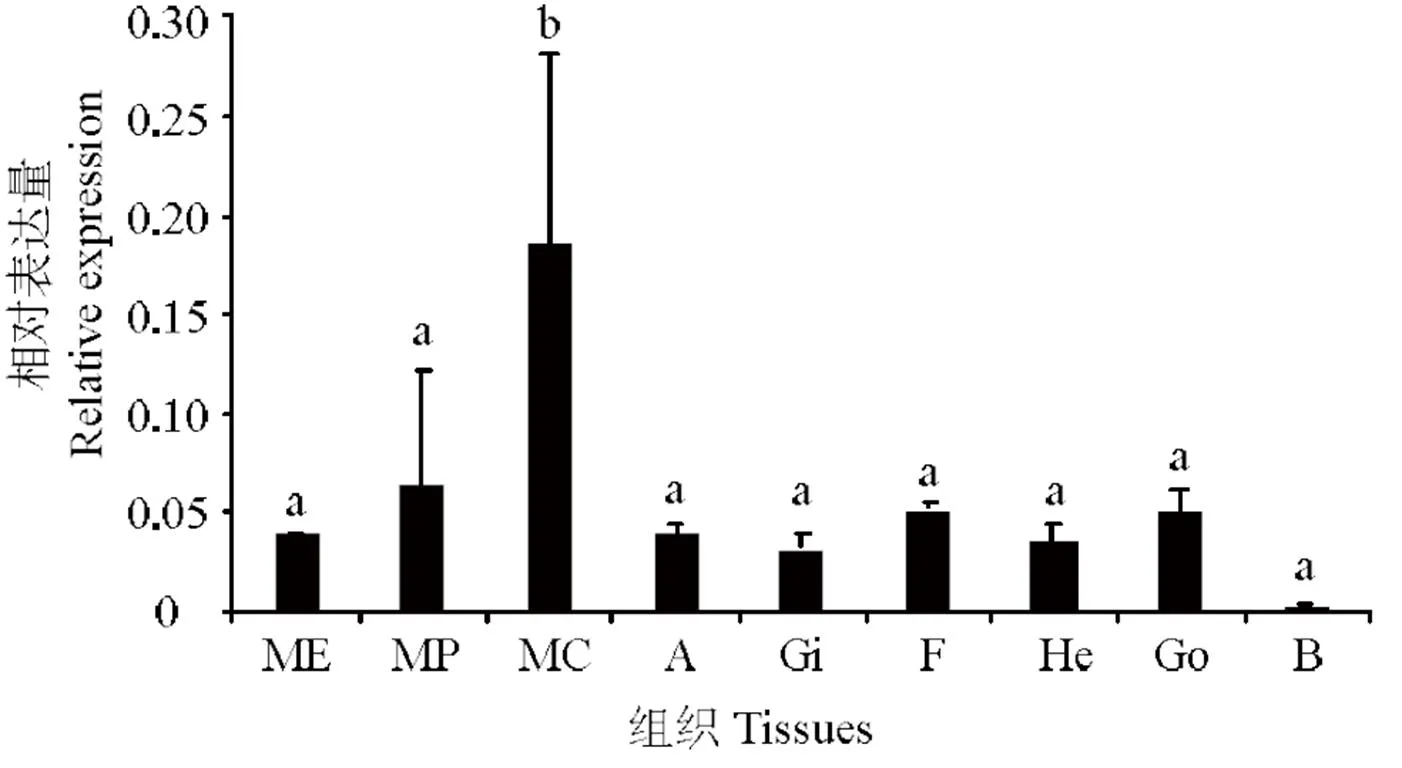
ME:邊緣膜;MP: 套膜;MC:中央膜;A:閉殼肌;Gi:鰓;F:足; He: 肝胰臟;Go:性腺;B:血細胞;
3 討論
DNA甲基化是一種常見的復制后修飾,參與生物機體內諸如調控基因表達、染色體結構維持、生長發育等多種重要生理過程的現象,主要由甲基轉移酶行使DNA甲基化修飾功能。本實驗通過RACE技術成功獲得基因序列全長,為4 818 bp,共編碼1 540個氨基酸。多序列比對分析結果顯示PmDnmt1與不同物種中的Dnmt1的比對結果中呈現較高的保守性。一般認為Dnmt1序列可以分為兩個區域N端蛋白識別區、C端催化區,其余位置上包含有半胱氨酸位點、BAH結構、鋅離子結合點等起到錨定蛋白和形成DNA甲基化等作用[15-16]。SMART預測分析發現具有DMAP結合結構域,DNMT1-RFD結構域、鋅指結構、BAH結構和C5胞嘧啶特異性DNA甲基化酶結構域等。DMAP結合結構域位于PmDnmt1的N端,在哺乳動物中,Dnmt1通過該位點與DMAP1(一種轉錄抑制因子)形成轉錄抑制復合物從而介導轉錄抑制[17],因此,可以推測PmDnmt1也具有轉錄抑制的功能。N端的DNMT1-RFD結構域與亞細胞定位、蛋白質結合和催化功能有關,研究表明,Dnmt1與裸露DNA和多核小體的結合活性受到DNMT1-RFD結構域的抑制,DNMT1-RFD結構域通過結合催化域而排除DNA發揮作用[18]。此外,PmDnmt1的鋅指結構為CXXC類型,該結構域包含8個保守的半胱氨酸殘基,能與2個鋅離子結合,是與非甲基化CpG二核苷酸結合的位點[19]。這些典型結構域使得甲基轉移酶具有穩定而廣泛的作用位點,進而更好地調控基因表達。
為了更好探究馬氏珠母貝甲基轉移酶在組織中的作用,使用實時熒光定量PCR結果顯示該基因mRNA在各個組織中均有表達,在中央膜中的表達量最高。外套膜是貝殼形成的主要器官,其不同區域所具有的結構特征和功能具有差異。貝殼按照形成的方式和組織的結構不同可分為3層,由內到外依次是珍珠質層,棱柱層及角質層。珍珠層是由中央的上皮細胞分泌形成,以文石為主;棱柱層結構由邊緣膜分泌形成,以方解石為主;角質層結構由外套膜的外、中褶殼皮溝細胞分泌形成。研究發現與貝殼棱柱層形成有關的基因通常在邊緣膜高表達,與貝殼珍珠層形成相關的基因則在中央膜和套膜區高表達[20]。馬氏珠母貝外套膜不同區域其DNA甲基化水平不一[11],而在維持基因組的甲基化水平起到重要作用,且影響到胚胎發育[21]、器官功能分化[22]與轉錄調控。結合與貝殼珍珠層形成相關的礦化基因普遍在中央膜和套膜上高表達[22],推測礦化基因的高表達需要DNA甲基化等表觀遺傳修飾參與調控,因而有可能通過維持甲基化修飾參與調控貝殼珍珠層的形成。
4 結論
本研究首次利用RACE技術從馬氏珠母貝克隆獲得基因cDNA全長序列,該基因在馬氏珠母貝外套膜中央膜中表達量最高,推測可能參與外套膜貝殼礦化基因的調控。
[1] MEEHAN R R. DNA methylation in animal development[J]. Seminars In Cell & Developmental Biology. 2003, 14: 53-65.
[2] RAZIN A. CpG methylation, chromatin structure and gene silencing-a three-way connection[J]. The EMBO Journal. 1998, 17(17): 4905-4908.
[3] AUTRAN D, HUANCA-MAMANI W, VIELLECA- LZADA J P. Genomic imprinting in plants: the epigenetic version of an Oedipus complex[J]. Current Opinion In Plant Biology. 2005, 8(1): 19-25.
[4] 張海燕, 趙洪斌, 田亞平, 等. DNA甲基化檢測技術與應用前景[J]. 標記免疫分析與臨床. 2013, 20(5): 348-351.
[5] SUBRAMANIAM D, THOMBRE R, DHAR A, et al. DNA methyltransferases: a novel target for prevention and therapy[J]. Frontiers In Oncology. 2014, 4: 80.
6] CUI D, XU X. DNA methyltransferases, DNA methylation, and age-associated cognitive function[J]. International Journal of Molecular Sciences. 2018, 19: 13155.
[7] SUZUKI M M, BIRD A. DNA methylation landscapes: provocative insights from epigenomics[J]. Nature Reviews Genetics. 2008, 9(6): 465-476.
[8] LYKO F, FORET S, KUCHARSKI R, et al. The honey bee epigenomes: differential methylation of brain DNA in queens and workers[J]. Plos Biology. 2010, 8(11): e1000506.
[9] RIVIERE G, WU G, FELLOUS A, et al. DNA methylation is crucial for the early development in the oyster[J]. Marine Biotechnology. 2013, 15(6): 739-753.
[10] 羅少杰, 張鵬飛, 焦鈺, 等. 馬氏珠母貝近交與雜交家系的 DNA 甲基化多態性比較[J]. 廣東海洋大學學報. 2016, 36(6): 9-15.
[11] 羅少杰, 鄧岳文, 鄭哲, 等. 馬氏珠母貝外套膜不同區域基因組DNA甲基化MSAP分析[J]. 中國水產科學. 2016, 23(6): 1227-1235.
[12] SONG K, LI L, ZHANG G. The association between DNA methylation and exon expression in the Pacific oyster[J]. Plos One. 2017, 12(9): e185224.
[13] FARIAS N D, DE OLIVEIRA N F P, DA SILVA P M. Perkinsus infection is associated with alterations in the level of global DNA methylation of gills and gastrointestinal tract of the oyster[J]. Journal of Invertebrate Pathology. 2017, 149: 76-81.
[14] DU X, FAN G, JIAO Y, et al. The pearl oystergenome and multi-omic analyses provide insights into biomineralization[J]. Gigascience. 2017, 6:1-12.
[15] 蘇玉, 王溪, 朱衛國. DNA甲基轉移酶的表達調控及主要生物學功能[J]. 遺傳. 2009, 31(11): 1087-1093.
[16] 王志剛, 建新. DNA甲基轉移酶分類、功能及其研究進展[J]. 遺傳. 2009, 31(9): 903-912.
[17] ROUNTREE M R, BACHMAN K E, BAYLIN S B. DNMT1 binds HDAC2 and a new co-repressor, DMAP1, to form a complex at replication foci[J]. Nature Genetics. 2000, 25(3): 269-277.
[18] FARISA S, FAGAN R L, MATTHEW W, et al. The replication focus targeting sequence (RFTS) domain is a DNA-competitive inhibitor of Dnmt1[J]. Journal of Biological Chemistry. 2011, 286(17): 15344-15351.
[19] CROSS S H, MEEHAN R R, NAN X, et al. A component of the transcriptional repressor MeCP1 shares a motif with DNA methyltransferase and HRX proteins[J]. Nature Genetics. 1997, 16(3): 256-259.
[20] FUNABARA D, OHMORI F, KINOSHITA S, et al. Novel genes participating in the formation of prismatic and nacreous layers in the pearl oyster as revealed by their tissue distribution and rna interference knockdown[J]. Plos One. 2014, 9: e847061.
[21] RAI K, NADAULD L D, CHIDESTER S, et al. Zebra fish Dnmt1 and Suv39h1 regulate organ-specific terminal differentiation during development[J]. Molecular and Cellular Biology. 2006, 26(19): 7077-7085.
[22] ANDERSON R M, BOSCH J A, GOLL M G, et al. Loss of Dnmt1 catalytic activity reveals multiple roles for DNA methylation during pancreas development and regeneration[J]. Developmental Biology. 2009, 334(1): 213-223.
Molecular Cloning and Expression Analysis of DNA Methyltransferase 1 () from
ZHANG Jia-bin1, LU Xiao-wen1, LUO Shao-jie1, JIAO Yu1, DENG Yue-wen1,2
(1.,524088,;2.,524025,)
【】In order to offer a theoretical basis for exploring the regulation of DNA methyltransferase 1 of() in biomineralization, the full-length gene was obtained and the tissue expression pattern ofwas elucidated. 【】The cDNA sequence ofwas obtained by rapid amplification of cDNA ends technology (RACE). The characteristics ofwere analyzed, and the mRNA expression ofin different tissues ofwere detected by fluorescence quantitative PCR. 【】The results showed that the length ofwas 4 818 bp,including ORF 4 623 bp which encoded 1 540 amino acids. The predicted molecular weight was about 174.55 ku and the theoretical isoelectric point was 5.89. The result of domain prediction analysis showed thathad DMAP binding domain, Dnmt1RFD, zinc finger structure, BAH domain and c-5 cytosine-specific DNA methylase domain profile. The multi-sequence alignment results showed that Dnmt1 was highly conserved across different species. Real-time fluorescence quantitative PCR analysis showed thatgene was expressed in all test tissue, with the highest expression in the mantle central (< 0.05).may be involved in shell formation through regulating the DNA methylation.
; DNA methyltransferase; gene clone; expression pattern
Q78
A
1673-9159(2019)05-0016-08
10.3969/j.issn.1673-9159.2019.05.003
2019-04-20
國家自然科學基金(31672626)
章佳斌,男,碩士研究生,研究方向為珍珠培育與加工。E-mail:874788088@qq.com
鄧岳文,教授,研究方向為珍珠貝遺傳育種與養殖。E-mail:dengyw@gdou.edu.cn
章佳斌,盧曉文,羅少杰,等. 馬氏珠母貝DNA甲基化轉移酶的克隆及表達[J]. 廣東海洋大學學報,2019,39(5):16-23.
(責任編輯:劉朏)
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
