綿羊GH基因的Pvu Ⅱ 位點的多態性與其生長性狀的關聯分析
2019-09-25 06:44:10白俊艷楊又兵樊紅燈付學言時坤鵬董智豪盧小寧李新月郝偉光李子衡鄭飛揚
浙江農業學報 2019年9期
關鍵詞:質量
白俊艷,曹 恒,王 旭,楊又兵,樊紅燈,付學言,時坤鵬,董智豪,盧小寧,李新月,郝偉光,李子衡,鄭飛揚
(河南科技大學 動物科技學院,河南 洛陽 471023)
生長激素基因(growth hormone,GH)是影響動物生長性狀的主效基因[1],具有提高飼料轉化[2]和促進肌肉中的蛋白質合成[3]等作用。Warwick等[4]首先克隆了綿羊的GH基因并將其定位于11號染色體,綿羊的GH基因由5個外顯子和4個內含子構成。劉士力等[5]研究表明,翹嘴鲌生長激素基因全長5 966 bp,其中轉錄單元長1 648 bp,由5個外顯子和4個內含子組成。牛志剛等[6]研究表明,GH基因的AhⅠ位點在新疆褐牛群體中表現為VV、LV和LL三種基因型,而且14月齡的LV和LL基因型個體的體質量、體長均比VV基因型的高,且差異顯著(P<0.05)。陳清等[7]研究表明,GH基因內含子3在3個地方鵝品種中檢測到AA、AB、BB三種基因型,而且不同基因型與4~10周齡體質量、屠體質量、半凈膛質量等19個性狀呈顯著相關。
許鋒[8]在4個蛋用品系雞中發現GH基因內含子3中存在2個AvaⅠ酶切位點,該酶切位點與蛋型指標關聯顯著(P<0.05)。董飚等[9]研究表明,在黑羽番鴨群體的公鴨中,GH基因第1內含子的CC基因型的宰前活質量、屠體質量、半凈膛質量、全凈膛質量、腹脂質量、腺肌胃質量顯著高于CD基因型和DD基因型(P<0.05)。海汀等[10]研究表明,GH基因對麥洼牦牛的體尺性狀無顯著影響。以上眾多研究表明,GH基因與畜禽的生長性狀和屠宰性能有著緊密關系,為此,本研究檢測了GH基因內含子Ⅱ在大尾寒羊、小尾寒羊、豫西脂尾羊、杜泊羊、湖羊群體中的多態性,并與綿羊體質量和體尺等性狀進行了關聯分析,以期為綿羊的標記輔助選擇和進一步選育提供理論依據。
1 材料與方法
1.1 基因組DNA提取
選擇河南省洛陽市馬坡鎮鮮達牧業杜泊羊50只,河南省洛陽市偃師和伊川縣的農戶豫西脂尾羊50只,河南省濮陽市臺前縣的農戶小尾寒羊60只,河南省平頂山市寶豐縣農戶大尾寒羊60只,河南省洛陽市肉羊產業技術體系洛陽綜合試驗湖羊50只,總計試驗綿羊270只。每只羊采集頸靜脈血液10 mL,采用ACD抗凝(1∶6),-20 ℃保存。用上海生工提供的全血DNA試劑盒(SK1262)提取基因組DNA。
1.2 引物設計與PCR擴增
GH基因內含子Ⅱ的引物序列參考施安[11],引物由鄭州鼎國生物技術有限公司合成。引物序列信息具體如下,F: 5′-CTGTTTGCCAACGCTGTGC-3′,R: 5′-CCCAAGCCACGACTGGATAAG-3′,片段大小為693 bp。
PCR擴增反應體系的總體積為15 μL,具體為去離子水3.5 μL,上游引物1 μL,下游引物1 μL,DNA為2 μL,2×TaqPCR Mix為7.5 μL。PCR擴增程序:94 ℃預變性5 min;94℃變性30 s,52 ℃退火30 s,72 ℃延伸30 s,34個循環,72 ℃后延伸7 min;4 ℃保存。
1.3 PCR-RFLP分析
GH基因內含子Ⅱ經限制性內切酶PvuⅡ(5′…CAG↓CTG…3′)消化酶切,依照產品說明書上推薦的酶切消化反應4 h,酶切后的產物在3%的凝膠電泳下檢測。酶切反應體系具體為ddH2O 7.4 μL,PCR產物10 μL,10×Buffer 2 μL,限制性內切酶PvuⅡ(10 U·μL-1) 0.6 μL,總計酶切反應體系20 μL。
1.4 關聯分析
GH基因內含子Ⅱ的PvuⅡ位點多態性與綿羊生長性狀進行關聯分析,分析模型如下:
yijkl=μ+Pi+Sj+Mk+eijkl。
(1)
式(1)中:yijkl為性狀表型值;μ為總體均值;Pi為品種效應(i=1,2,3,4,5);Sj為性別效應(j=1,2);Mk為第k種基因型效應;eijkl為殘差效應。結果用平均值±標準差來表示。
2 結果與分析
2.1 綿羊GH基因內含子Ⅱ的PvuⅡ位點的PCR擴增產物檢測
五個綿羊群體的GH基因內含子Ⅱ的PvuⅡ位點的PCR擴增產物的檢測結果見圖1,從圖1可以看出,GH基因內含子Ⅱ的PvuⅡ位點的擴增片段約為693 bp,與目的條帶一致,且條帶清晰、無雜帶,可以進行后續PCR-RFLP分析。
2.2 綿羊GH基因內含子Ⅱ的PvuⅡ位點的PCR-RFLP分析
大尾寒羊、小尾寒羊、豫西脂尾羊、杜泊羊、湖羊的GH基因內含子Ⅱ的PvuⅡ位點進行酶切結果見圖2-圖5,酶切后檢測到AB基因型(264 bp/429 bp/693 bp)和BB基因型(264 bp/429 bp)共兩種基因型。
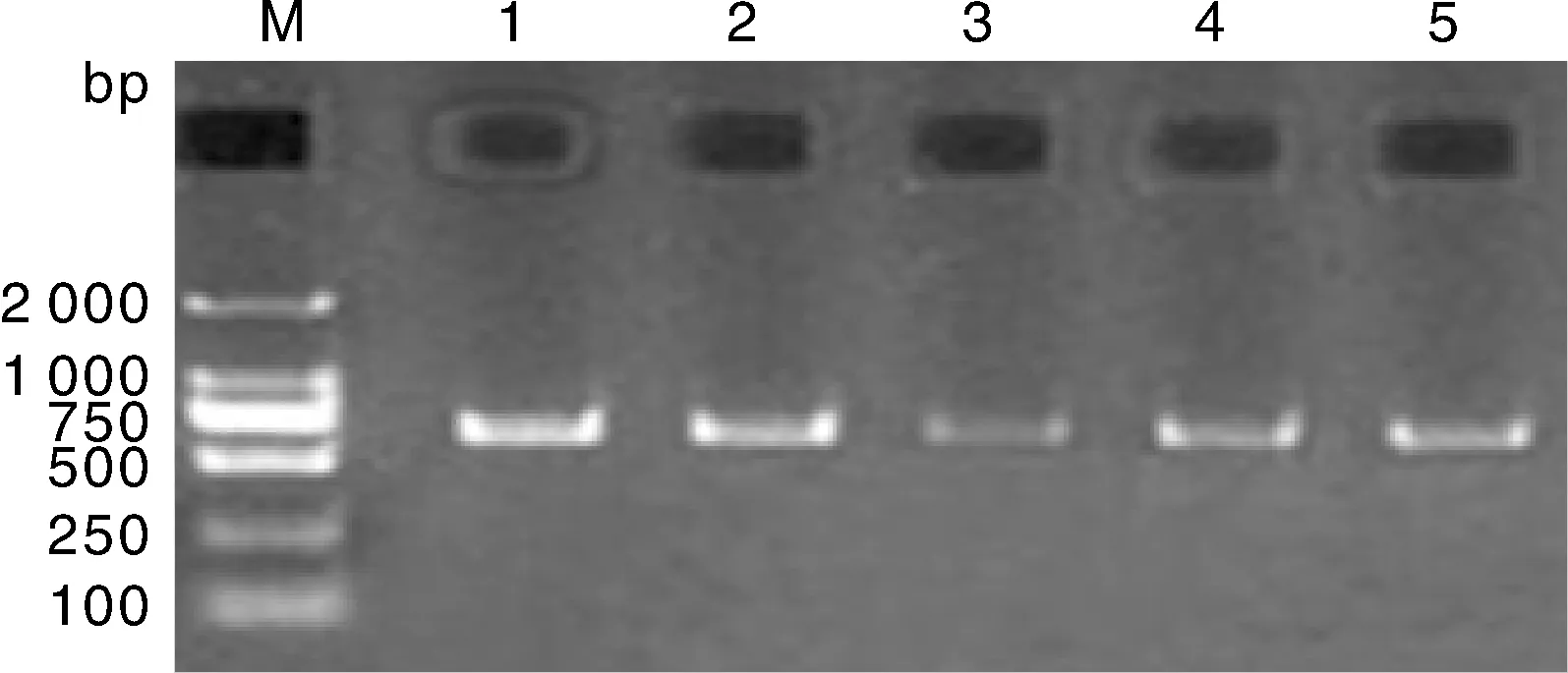
M,Marker DL2000;1、2、3、4、5分別為大尾寒羊、小尾寒羊、豫西脂尾羊、杜泊羊、湖羊。M, Marker DL2000; 1, 2, 3, 4 and 5 were Large-tailed Han sheep, Small-tailed Han sheep, Yuxi fat-tailed sheep, Dorper sheep and Hu sheep, respectively.圖1 五個綿羊群體的GH基因內含子Ⅱ的PvuⅡ位點的檢測結果Fig.1 Detection of PvuⅡ locus in intron Ⅱ of GH gene in sheep populations
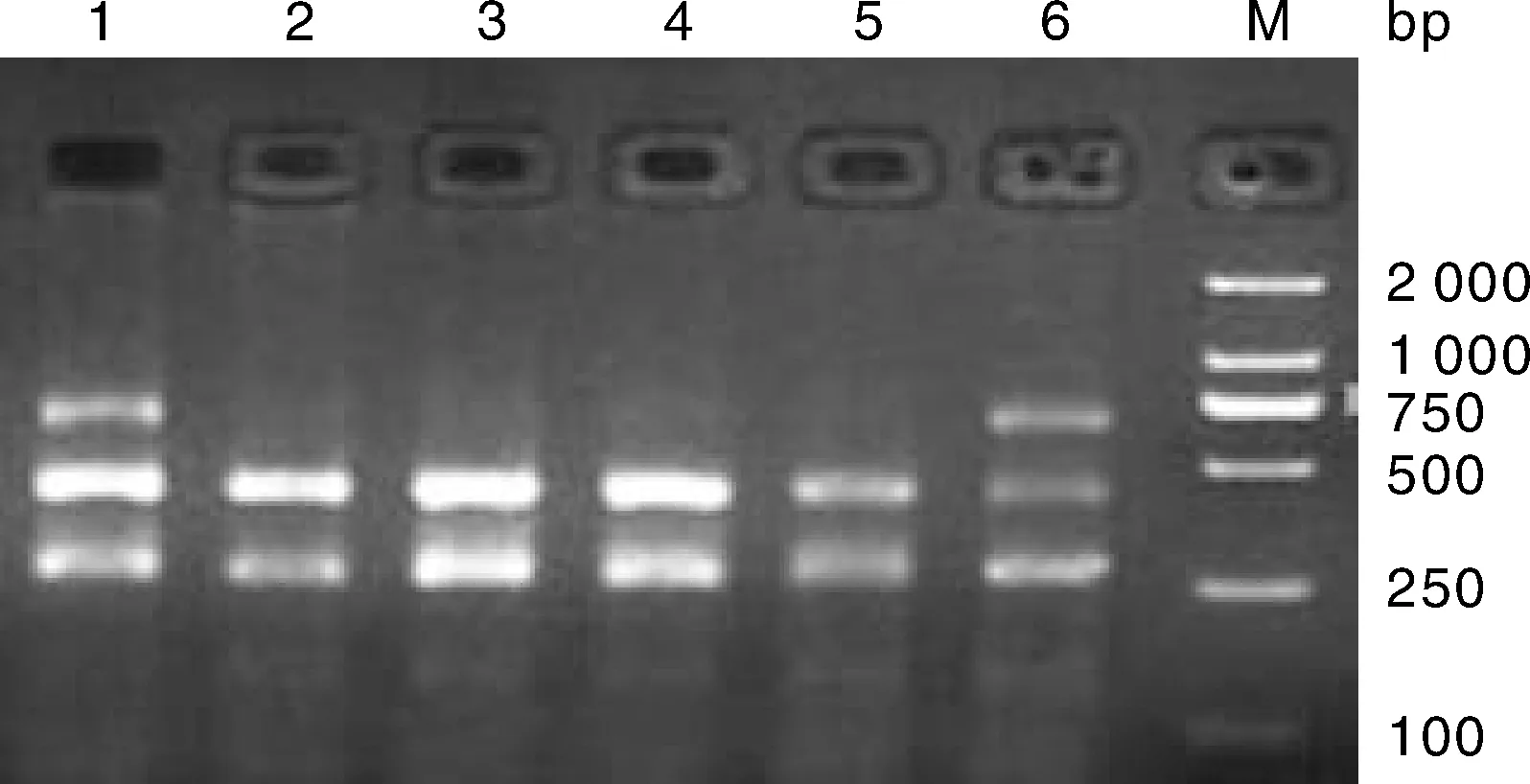
1、6,AB基因型;2、3、4、5,BB基因型;M,Marker DL2000。1, 6: AB genotypes; 2, 3, 4, 5: BB genotypes; M, Marker DL2000. 圖2 小尾寒羊的GH基因PvuⅡ位點多態性Fig.2 Polymorphism of GH gene PvuⅡ locus in Small-tailed Han sheep
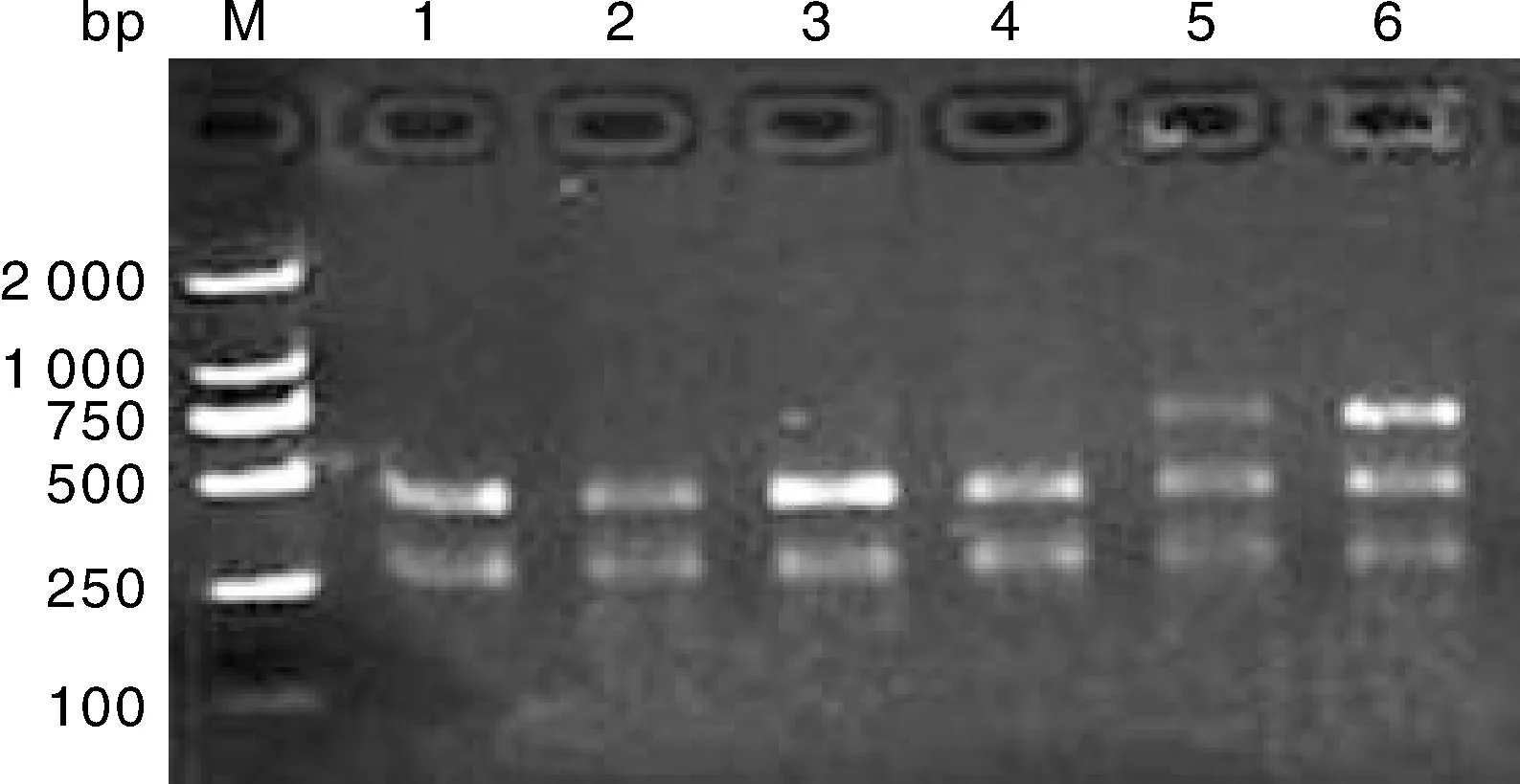
5、6,AB基因型;1、2、3、4,BB基因型;M,Marker DL2000。5, 6: AB genotypes;1, 2, 3, 4: BB genotypes; M, Marker DL2000. 圖3 豫西脂尾羊的的GH基因PvuⅡ位點多態性Fig.3 Polymorphism of GH gene PvuⅡ locus in Yuxi fat-tailed sheep
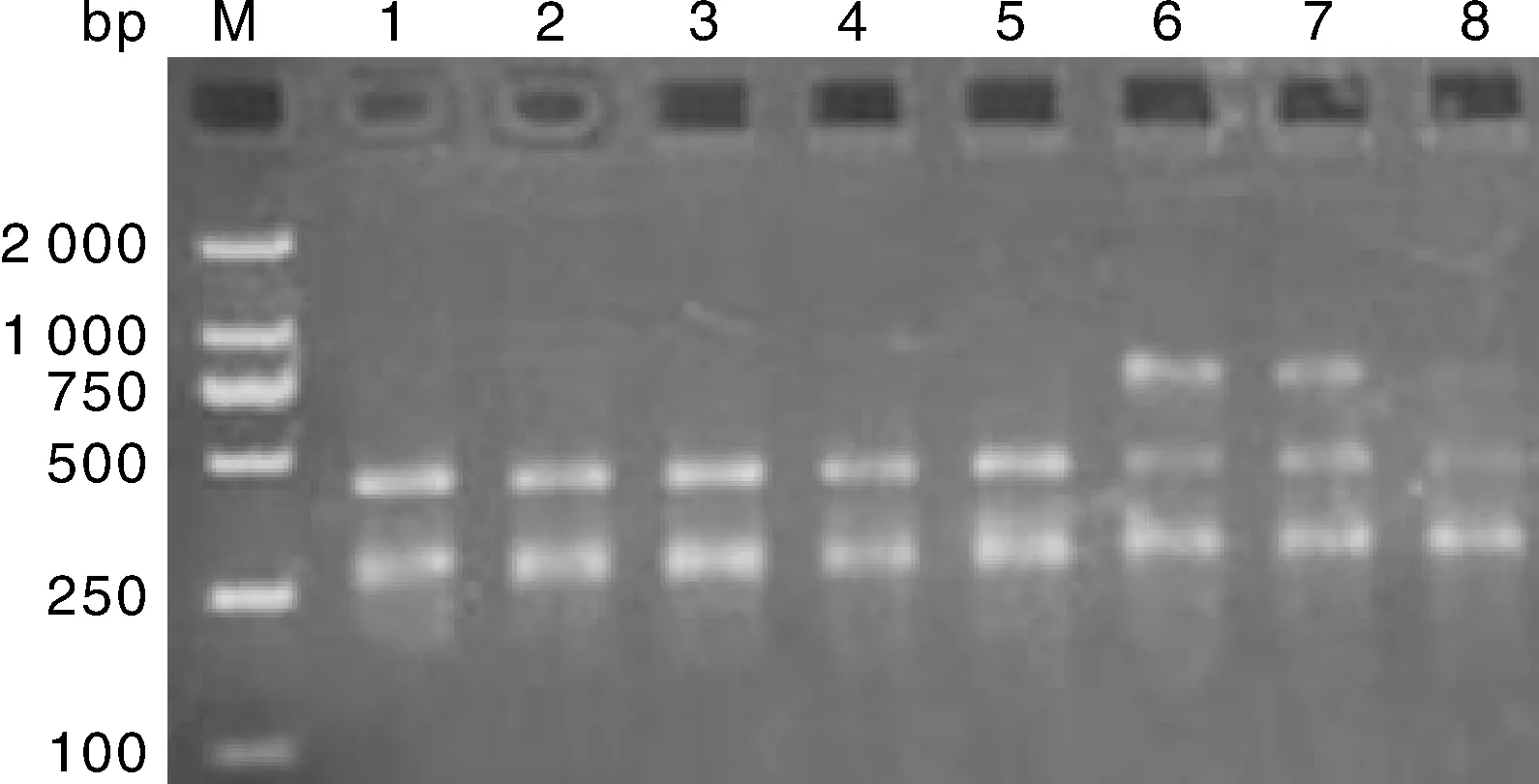
6、7、8,AB基因型;1、2、3、4、5,BB基因型;M,Marker DL2000。6, 7, 8: AB genotypes; 1, 2, 3, 4, 5: BB genotypes; M, Marker DL2000.圖4 湖羊的GH基因PvuⅡ位點多態性Fig.4 Polymorphism of GH gene PvuⅡ locus in Hu sheep
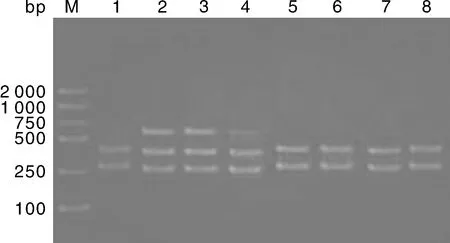
2、3、4,AB基因型;1、5、6、7、8,BB基因型;M,Marker DL2000。2, 3, 4: AB genotypes; 1, 5, 6, 7, 8: BB genotypes; M, Marker DL2000.圖5 杜泊羊的GH基因PvuⅡ位點多態性Fig.5 Polymorphism of GH gene PvuⅡ locus in Dorper sheep
2.3 綿羊GH基因內含子Ⅱ的PvuⅡ位點的群體遺傳學分析
綿羊GH基因內含子Ⅱ的PvuⅡ位點的群體遺傳學分析結果見表1,可以看出,GH基因內含子Ⅱ的PvuⅡ位點在大尾寒羊、小尾寒羊群體中AB基因型頻率最高(0.786和0.750),而在豫西脂尾羊、湖羊、杜泊羊群體中以BB基因型頻率最高(0.576、0.529和0.541)。GH基因內含子Ⅱ的PvuⅡ位點在5個綿羊群體中均以B等位基因頻率為最高。GH基因內含子Ⅱ的PvuⅡ位點在5個綿羊群體中的有效等位基因數為1.497~1.908,多態信息含量(PIC)為0.277~0.363,呈現為中度多態性(0.25
2.4 綿羊GH基因內含子Ⅱ的PvuⅡ位點與生長性狀的關聯分析
GH基因內含子Ⅱ的PvuⅡ位點的多態性與綿羊生長性狀的關聯分析結果見表2,從表2可以看出,AB基因型的體質量、體長、胸寬、臀端高、管圍、尻高、頸長上顯著高于BB基因型(P<0.05),GH基因內含子Ⅱ的PvuⅡ位點對其他性狀無顯著影響(P>0.05)。
3 討論
由于生長激素基因在動物生長發育中有著重要的生理作用,對GH基因的研究就成為同類研究中的一個重要課題。Valinsky等[12]對綿羊GH基因第2內含子進行擴增,結果發現了2個等位基因。Bastos等[13]檢測到葡萄牙綿羊品種中GH基因第4、5外顯子的多態情況。任春環[14]通過PCR-SSCP的方法對4個綿羊品種的GH基因進行了多態性檢測,結果表明,在所檢測的第1內含子、第2外顯子、第2內含子、第3外顯子、第3內含子、第4外顯子、第4內含子、第5外顯子位點中,僅在第3內含子和第4外顯子檢測到SNP,其余位點均未檢測到SNP。于洪川等[15]利用限制性片段長度多態性技術在不同灘羊群體中發現酶切位點并命名為AB、BB基因型。馬麗娜[16]對GH基因在灘羊群體中多態性研究表明,GH基因在灘羊群體中具有多態性,出現AB、BB兩種基因型,頻率為0.362 3、0.637 7。本研究采用PCR-RFLP的方法在大尾寒羊、小尾寒羊、豫西脂尾羊、杜泊羊、湖羊5個綿羊群體中,對GH基因內含子Ⅱ的PvuⅡ位點只檢測到了AB基因型和BB基因型,這與施安[11]、于洪川等[15]、馬麗娜[16]等的研究結果相一致,與任春環[14]的研究結果不符。
表1 綿羊GH基因內含子Ⅱ的PvuⅡ位點的遺傳多態性
Table1Genetic polymorphism ofPvuⅡ locus in intron Ⅱ ofGHgene in sheep

遺傳多態性Genetic polymorphism頻率Frequency大尾寒羊Large-tailedHan sheep小尾寒羊Small-tailedHan sheep豫西脂尾羊Yuxi fat-tailedsheep湖羊Hu sheep杜泊羊Dorper sheep基因型頻率Genotype frequencyBB0.2140.2500.5760.5290.541AB0.7860.7500.4240.4710.459基因頻率Gene frequencyA0.3930.3750.2120.2350.229B0.6070.6250.7880.7650.771雜合度Heterozygosity0.4760.4660.3320.3640.354多態信息含量Polymorphic information content0.3630.3580.2770.2980.291有效等位基因數Number of effective alleles1.9081.8761.4971.5751.547χ2值3.5717.4588.7275.8827.149P0.0590.0080.0030.0150.008
表2GH基因內含子Ⅱ的PvuⅡ位點與綿羊生長性狀的關聯分析
Table2Association ofPvuⅡ locus in intron Ⅱ ofGHgene with growth traits in sheep
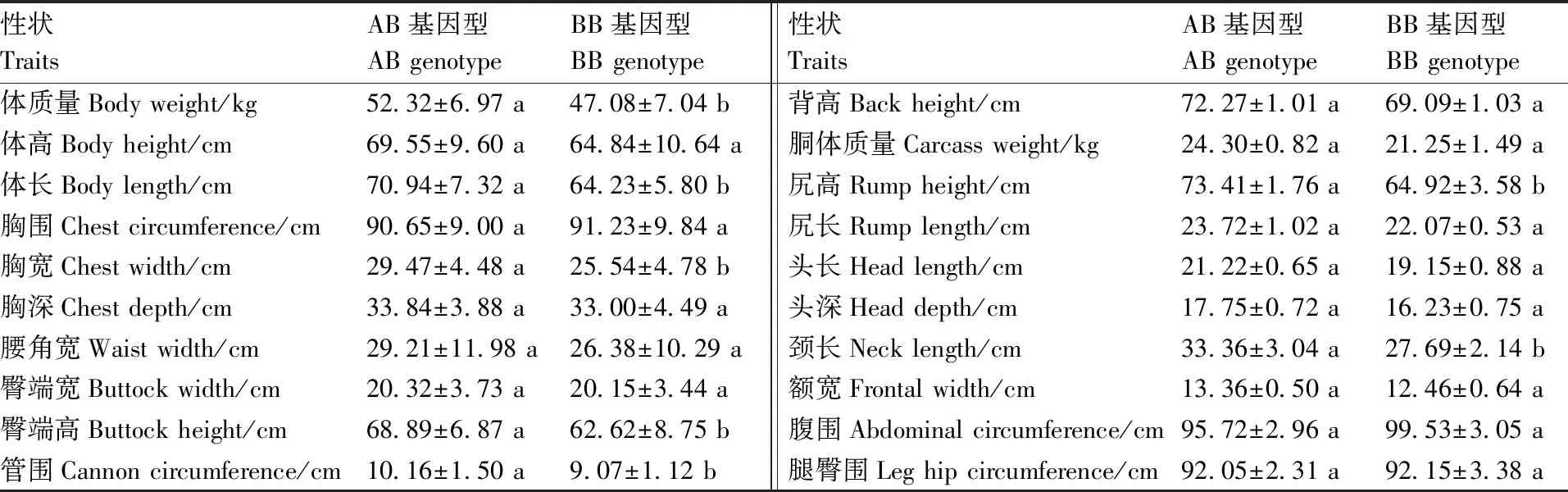
性狀TraitsAB基因型AB genotypeBB基因型BB genotype性狀TraitsAB基因型AB genotypeBB基因型BB genotype體質量Body weight/kg52.32±6.97 a47.08±7.04 b背高Back height/cm 72.27±1.01 a69.09±1.03 a體高Body height/cm 69.55±9.60 a64.84±10.64 a胴體質量Carcass weight/kg 24.30±0.82 a21.25±1.49 a體長Body length/cm 70.94±7.32 a64.23±5.80 b尻高Rump height/cm 73.41±1.76 a64.92±3.58 b胸圍Chest circumference/cm90.65±9.00 a91.23±9.84 a尻長Rump length/cm 23.72±1.02 a22.07±0.53 a胸寬Chest width/cm 29.47±4.48 a25.54±4.78 b頭長Head length/cm 21.22±0.65 a19.15±0.88 a胸深Chest depth/cm 33.84±3.88 a33.00±4.49 a頭深Head depth/cm 17.75±0.72 a16.23±0.75 a腰角寬Waist width/cm 29.21±11.98 a26.38±10.29 a頸長Neck length/cm 33.36±3.04 a27.69±2.14 b臀端寬Buttock width/cm 20.32±3.73 a20.15±3.44 a額寬Frontal width/cm 13.36±0.50 a12.46±0.64 a臀端高Buttock height/cm 68.89±6.87 a62.62±8.75 b腹圍Abdominal circumference/cm 95.72±2.96 a99.53±3.05 a管圍Cannon circumference/cm10.16±1.50 a9.07±1.12 b腿臀圍Leg hip circumference/cm 92.05±2.31 a92.15±3.38 a
同行不同列數據后沒有相同小寫字母表示差異顯著(P<0.05)。
The values in the same row followed by different lower-case letters showed the significant difference (P<0.05).
胡沈榮等[17]分析了5個山羊品種GH基因,除了波爾山羊外,其他山羊品種均有多態性,且GH基因外顯子5位點的基因頻率分布與品種特性有關,外顯子2與產羔多態性有顯著關聯(P<0.05)。張春香等[18]在南江黃羊生長激素第3外顯子發現G/A突變與該品種6月齡和12月齡體質量和體高有影響。閔令江等[19]通過PCR擴增了山羊5′調控區序列,共得到5處突變位點,不同突變位點均與山羊體質量、體尺性狀關聯性較強。張建軍等[20]以隴東山羊為研究對象,在GH基因外顯子4區域發生堿基突變,與6月齡質量,周歲質量,胸圍等指標顯著相關。張浩等[21]在甘肅現代肉羊品種中GH外顯子1上發現301 bp處的T/A突變造成AA、AB基因型,數據關聯顯示與肉羊體質量關聯顯著,且表現為雜合型明顯高于純合型。馬麗娜[16]對GH基因在灘羊群體中多態性研究表明,GH基因不同基因型(AB和BB)與初生質量、二毛活質量、三月體質量和六月體質量差異均極顯著(P<0.01)。多重比較結果顯示,AB基因型個體各年齡階段體重的最小二乘均值均極顯著高于BB基因型(P<0.01)。王鵬[22]發現湖羊GH基因的外顯子3位點的GH基因型和外顯子4位點的LM基因型的初生質量、斷奶質量和六月齡質量平均值均相對于另一個基因型。李文文等[23]通過對7個綿羊品種GH基因檢測發現5′端調控區、第4外顯子3′端區域均存在多態性,其中山谷型藏羊不同基因型個體間的體質量、體長、體高、胸圍具有顯著差異(P<0.05)。施安[11]研究表明,灘羊的GH基因內含子2的AB基因型初生質量、斷奶質量、6月齡體質量顯著高于BB基因型(P<0.05)。本研究表明,綿羊GH基因內含子Ⅱ的PvuⅡ位點AB基因型的體重顯著高于BB基因型(P<0.05),這與施安[11]和馬麗娜[16]研究結果相符,此外本研究還表明,GH基因內含子Ⅱ的PvuⅡ位點對體長、胸寬、臀端高、管圍、尻高、頸長等有顯著影響(P<0.05),這與閔令江等[19]和李文文等[23]的研究結果相似。可見,GH基因內含子Ⅱ的PvuⅡ位點可以作為綿羊生長性狀的候選標記進行標記輔助選擇。
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
