2016—2018年四川地區豬流行性腹瀉病毒流行病學調查與ORF3、S1部分基因序列分析
2019-09-23 10:47:30德西措姆楊澤曉姚學萍廖倡宇張鵬飛江地科項明源姜瑞姣
浙江農業學報 2019年9期
關鍵詞:分析
德西措姆,王 印,楊澤曉,姚學萍,羅 燕,廖倡宇,張鵬飛,江地科,項明源,姜瑞姣,宋 勇
(1.昌都市左貢縣畜牧站,西藏 昌都 854499; 2.四川農業大學 動物醫學院,動物疫病與人類健康四川省重點實驗室,四川 成都 611130; 3.天津瑞普生物技術股份有限公司,天津 300300)
豬流行性腹瀉(porcine epidemic diarrhea,PED)是由豬流行性腹瀉病毒(porcine epidemic diarrhea virus,PEDV)引起的一種急性接觸性腸道傳染病[1],可發生于任何年齡的豬,年齡越小,癥狀越重,死亡率越高,嚴重危害我國養豬業[2]。近年來,隨著我國養殖業的快速發展,規模化、集約化豬場數增加,該病的發病率呈上升趨勢,毒株類型呈多樣化發展趨勢[3],危害日趨嚴重。
PEDV為冠狀病毒科冠狀病毒屬成員,形態略呈球形[4],平均直徑80~120 nm,核衣殼呈螺旋對稱,為二十面體對稱[5]。PEDV的結構蛋白主要有4種:纖突蛋白(S)、膜蛋白(M)、囊膜蛋白(E)和核衣殼蛋白(N)[6]。Park等[7]克隆了16個不同PEDV毒株的ORF3,發現致弱病毒較野生型病毒均缺失17個氨基酸,推測缺失的氨基酸與病毒病原性密切相關;Wongthida等[8]研究發現,PEDV的ORF3可編碼1種調節病毒復制鉀離子的通道蛋白,與PEDV發病機制有關,完整的ORF3對病毒離子通道的作用可增強PEDV的產生。
目前臨床上對于PEDV的防控,主要是采用傳統疫苗,隨著PEDV變異菌株的流行,傳統疫苗的防控效果逐漸下降[9]。我國于1976年首次報道PED,并于1980年首次分離到PEDV,此后在國內開始流行。1977年,比利時人從病料中分離出1株PEDV弱毒株,并命名為類冠狀病毒CV777株[10]。2018年4月,隨著Zhou等[11]報道的能引起仔豬腹瀉等相似癥狀的豬只新型冠狀病毒(swine acute diarrhea syndrome coronavirus,SADS-CoV),造成豬腹瀉的病毒已呈種類繁多、株型多樣化的現狀,對于疾病的防控十分不利。
2017年冬、春兩季,四川地區多地發生仔豬大規模腹瀉,對養殖業造成巨大損失。初生仔豬死亡率上升,染病仔豬出現厭食、嘔吐、脫水、消瘦等癥狀,糞便為黃色水樣,并伴有腥臭味。經剖檢發現,病豬小腸絨毛脫落,腸道內含大量乳樣糞便,部分腎臟有出血點。本研究使用RT-PCR法對四川地區2016年6月至2018年6月發生腹瀉的仔豬病料進行流行病學檢測,同時選取4株來源不同、發病狀況典型的PEDV感染樣本,對其ORF3及S1基因高變區段進行序列分析,旨在為探討當前我國PEDV發生發展的規律、疫苗的篩選及制定有效防控措施提供科學依據。
1 材料與方法
1.1 試驗病料
病料來自四川省遂寧、樂山、成都、綿陽等地豬場,共計71份,無菌采集腹瀉仔豬小腸內容物和糞便,經反復凍融3次后,用生理鹽水進行稀釋,4 ℃ 5 000 r·min-1離心5 min,取上清于-70 ℃凍存備用。
1.2 主要試劑
病毒DNA/RNA提取試劑盒,細菌基因組DNA提取試劑盒,凝膠回收試劑盒,大腸桿菌DH5α等,天根生化科技(北京)有限公司;2×TaqPCR Master Mix,DNA分子標準,成都擎科生物科技公司;PrimeScriptTMRT reagent Kit,寶生物技術(北京)有限公司;其他試劑均為國產分析純。
1.3 引物設計與合成
PEDV檢測引物參照NY/T 544—2002《豬流行性腹泄診斷技術》,ORF3、S1基因擴增引物參照美國高毒株PC22A(GenBank No.KY499262.1)設計(表1),由生工生物工程(上海)股份有限公司合成。
表1 引物序列
Table1Primer sequences

引物 Primers引物序列Primer sequences(5′-3′)長度 Length/bp位置 Position標準引物Standard primerP1: CCTAGACTTCAACCTTACGA77424 750~25 523P2: CAGGAAAAAGAGTACGAAAAORF3基因 OFR3 geneP1: TAGTGTTCTGCTGCATTTCC93124 690~25 600P2: TAAACTGCGCTATTACACAACCS1部分基因 S1 parital geneP1: TTGTGATGATCCTGTTAGC98822 022~22 987P2: AATACTCATACTAAAGTTGGTGG
1.4 樣品核酸提取
取病料上清液,參照病毒DNA/RNA提取試劑盒操作說明書提取病毒和細菌核酸,用適量RNase free水溶解,-20 ℃保存備用。
1.5 病毒核酸檢測及ORF3基因、S1部分基因擴增及測序
取提取的DNA作PCR模板,反應體系為:2×TaqPCR Master Mix 20 μL,ddH2O 10 μL,上下游引物各2 μL,模板4 μL。反應條件為:95 ℃ 5 min;94 ℃ 1 min,56 ℃ 30 s,72 ℃ 2 min,34個循環;72 ℃ 10 min。將PCR產物純化后,送至華大基因進行測序。
1.6 統計分析
采用圓形分布法[12]進行集中趨勢分析,GraphPad Prism 5進行直觀記錄,計算發病高峰時點及高峰時期。圓形分布法的核心內容是將發病時間轉換成角度來計算角均數,用角均數來表示疾病發病的高峰時點,并推算出相應的高峰時期。運用Excel對陽性病例及樣本地區病史情況進行卡方檢驗,P<0.05為差異顯著。
1.7 同源性分析及遺傳進化樹分析
測序結果利用DNAstar同源性分析、Mega 6.0軟件進行分析。進化樹采用Neighbor-joining方法1 000次舉例進行繪制。
2 結果與分析
2.1 流行病學調查
2016年6月至2018年6月,共采集腹瀉臨床病例71例,實驗室確診PEDV感染病例35例。PEDV感染病例在腹瀉總樣本中,占比較大,達39%(表2)。PEDV發病狀況呈明顯的季節聚集性,PEDV高發期為冬、春兩季,夏、秋兩季發病狀況較為好轉。
如表3所示,通過對病史資料統計,發現發生PEDV的地區新疫源地所占比例較大,達61%。存在兩年病史的豬場PEDV發病率低于1年病史豬場和新疫源地。老疫源地或有過病史存在的地區,發病數呈現下降趨勢,表明PEDV防控取得良好的效果,但整體形式依舊嚴峻。
參考文獻[12],將2016—2018年四川部分地區PEDV月發病數轉換為角度,結果見表4。根據圓形分布法基本公式,對PEDV發病高峰期進行預測,計算出PEDV發病高峰日為1月28日和2月11日,發病高峰時間為12月~3月,與實際調查結果一致。
2.2 PEDV ORF3基因及S1基因片段擴增
將選取的4株PEDV分別命名為scds1、scds2、scds3、scds4。擴增出的ORF3基因目的條帶大小為931 bp,S1高變區段基因目的條帶大小為988 bp(圖1)。
表2PEDV流行病學調查結果
Table2PEDV epidemiological survey results
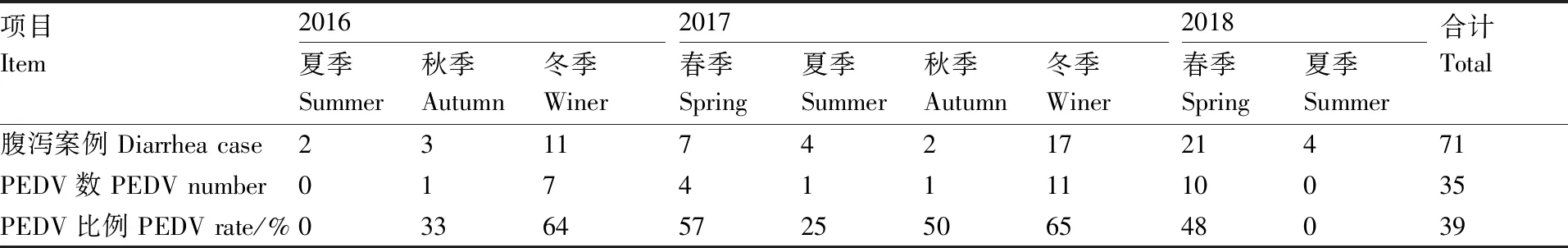
項目Item2016夏季Summer秋季Autumn冬季Winer2017春季Spring夏季Summer秋季Autumn冬季Winer2018春季Spring夏季Summer合計Total腹瀉案例 Diarrhea case23117421721471PEDV數 PEDV number0174111110035PEDV比例 PEDV rate/%033645725506548039
表3PEDV陽性地區或豬場病史調查結果
Table3Results of PEDV-positive areas or pig farm history

項目Item兩年內無病史No history of illnesswithin two years1年病史1 yearhistory2年病史2 yearhistory陽性數17114Positive number陰性數111724Negative number合計 Total282828陽性率613914Positive rate/%
表4 四川部分地區2016—2018年PEDV月發病數角度計算表
Table4Calculation of the incidence of PEDV in the two years from 2016 to 2018 in some parts of Sichuan

月份 Monthfαfsinαfcosα1915.29 2.373 3428.681 4312444.38 2.797 6562.858 8673773.48 6.711 0441.990 45044103.56 3.888 500-0.937 85053133.64 2.171 071-2.070 37060163.730071193.81 -0.238 700-0.971 09080224.380092254.47 -1.926 980-0.535 490100284.5500110314.6300125344.71 -1.318 5204.823 017合計Total35—14.457 40013.838 960
2.3 PEDV分離株同源性分析
將分離的4株PEDV菌株(scds1、scds2、scds3、scds4)與15株已有菌株(表5)的ORF3基因進行同源性分析,發現4株PEDV菌株與OH851、PC22A、GER/L00719/2014、CH/GDZH02/1401、CH/GDZHDM/1401在同一分支(圖2)。通過對S1基因高變區進行同源性進化樹的構建,將目前流行的毒株分成2個大群,4個亞型。scds1、scds2、scds3、scds4均屬于GII群,與CH/GDZH02/1401、CH/GDZHDM/1401、PC22A、AJ1102等參考株同屬一分支,為當前國內流行的PEDV變異毒株群。
通過對scds1、scds2、scds3、scds4菌株OFR3基因核苷酸序列進行比對,結果表明,以上4株與CH/GDZH02/1401、CH/GDZHDM/1401、PC22A等變異毒株同源性高達99%,其中scds1、scds2的ORF3基因的第158位、第259位發生突變分別為C→T、T→C。未發生弱毒株的堿基缺失現象,臨床癥狀亦符合強毒株特征。S1基因高變區段堿基更替較大,與CH/GDZH02/1401等變異毒株同源性為92.8%~99.3%;與CV777、DR13等G1亞型的經典株親緣關系較遠,同源性為82.7%~87.3%。
3 討論
豬流行性腹瀉是一種對仔豬危害較大的傳染病,多發于秋、冬、春3季。通過對四川地區近兩年的PEDV流行病學資料的整理分析,發現該地區發病呈明顯的季節聚集性,高峰期從12月持續至第2年的3月,與冠狀病毒高發時間大致吻合。研究發現,經長期免疫防控的豬場PEDV感染率明顯降低,可能是因長期的免疫接種和良好的生物安全措施致使帶毒豬轉為隱性感染狀態,病毒感染的發病閾值提高,感染風險下降。而未有病史的豬場,由于缺乏對該病的防治手段,存在該病暴發流行的風險。

M,DL 2 000 marker;1,陰性對照;2,scds1;3,scds2;4,scds3;5,scds4。A,ORF3基因;B,S1基因。M, DL 2 000 marker; 1, Negative control; 2, scds1; 3, scds2; 4, scds3; 5, scds4. A, ORF3 gene; B, S1 gene.圖1 PEDV ORF3、S1基因擴增結果Fig.1 PEDV ORF3, S1 gene amplification results
表5 毒株說明
Table5Description of the strain

編號No.株名Strains登錄號GenBank No.分離年份Collectedyear分離地Collectedarea1AJ1102JX1884542012中國China2GER/L00719/2014LM6450582014德國Germany3PC22AKY4992622015美國USA4MNKF4687522013美國USA5CH-GDZH02-1401KR1533252014中國China6CH-GDZHDM-1401KR1533262014中國China7NPL-PEDv/2013/p10.1KM0523652014美國USA8CHN/SH/2016-4/2016MG8370122016中國China9IA1KF4687532013美國USA10OH851KJ3999782014美國USA11DR13JQ0231612012韓國Korea12Br1/87LT9065822017德國Germany13CV777AF3535112001比利時Belgium14LZCEF1859922007中國China15SM/98gbGU9377972011韓國Korea

圖2 PEDV ORF3基因進化樹分析Fig.2 Phylogenetic tree analysis based on PEDV ORF3 gene

圖3 PEDV S1基因進化樹分析Fig.3 Phylogenetic tree analysis based on PEDV S1 gene
PEDVORF3基因可編碼鈣離子通道蛋白,調節病毒產量與毒力,ORF3蛋白可抑制IFN-β mRNA轉錄,具有拮抗干擾素產生的功能[13]。同時,ORF3基因是鑒別PEDV強弱毒株的重要基因,本研究中,4株PEDV與國內外的變異高毒毒株的ORF3基因具有高度相似。近年來也有研究表明,PEDV可以通過PK15傳代[14],仔豬感染PEDV后,常見腎小球病變[15-16]。腎臟作為鉀離子代謝的重要器官,富集較多的鉀離子,推測PEDV增殖可能與鉀離子濃度存在一定的相關性。
PEDV的S1基因核苷酸更替上具有明顯的優勢,通過對S1基因進行基因分型,可將PEDV劃分為2個大群、4種亞種。本研究中,4株PEDV的S1基因高變區段與2014年廣東、美國等地出現的PEDV高毒毒株歸為同一分支,與CV777親緣關系較遠。當豬只感染PEDV時,由于毒株產生較大變異,易導致疫苗免疫效果不理想。通過對ORF3基因進行序列分析,未出現弱毒株部分核苷酸片段缺失情況,臨床發病情況上也表現為強毒株特征,可通過對ORF3缺失情況與S1基因分型結果相結合,有針對性地進行疫苗毒株的初步篩選,對當前流行的PEDV進行防控。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
