魁蚶4個群體形態差異比較分析
2019-09-18 10:29:42王英俊吳瑩瑩麻丹萍宋愛環劉洪軍
水產科學 2019年5期
關鍵詞:差異
劉 童,鄒 琰,王英俊,吳瑩瑩,麻丹萍,宋愛環,劉洪軍
( 山東省海洋生物研究院,青島市海洋生物種質資源挖掘與利用工程實驗室,山東省海水健康養殖工程技術研究中心,山東 青島 266104 )
魁蚶(Scapharcabroughtonii),俗稱血貝、赤貝、大毛蛤,是重要的大型冷水性海洋經濟貝類,營埋棲生活,主要分布于中國、朝鮮、韓國、日本和俄羅斯,在我國以山東和遼寧的資源量最為豐富[1]。魁蚶肉質肥美,具有良好的市場前景和開發利用價值[1]。由于產品深受國內外市場歡迎,致使捕撈強度增加,導致魁蚶的自然資源量急劇下降,我國原有的魁蚶種質資源已遭受嚴重破壞[2]。因此,進行魁蚶種質資源保護評價,開展魁蚶遺傳輔助育種研究已顯得十分必要。
形態性狀是水產經濟動物種質遺傳信息的綜合外在體現,是評估水產動物生長性能和生產性能的重要依據[3-8]。多元統計分析方法是多種水產動物形態性狀研究中的常見方法,主要包括相關性分析、主成分分析、聚類分析和判別分析等,在貝類中已廣泛應用,如鈍綴錦蛤(Tapesdorsatus)[4]、菲律賓蛤仔(Ruditapesphilippinarum)[9]、合浦珠母貝(Pinctadamartensii)[10]、翡翠貽貝(Pernaviridis)[11]、彩虹明櫻蛤(Moerellairidescens)[12]、文蛤(Meretrixmeretrix)[13]、泥蚶(Tegillarcagranosa)[14]等。
在魁蚶養殖生產實踐中,發現中韓兩國魁蚶在外部形態、生長速度、成活率等方面存在差異,韓國魁蚶在生長速度、成活率等方面有一定的優勢[15]。為深入了解魁蚶中國群體與韓國群體遺傳背景的差異,筆者以3個中國群體和1個韓國群體為研究對象,通過多元統計分析方法分析4個地理種群的形態差異,為魁蚶種質資源的合理開發和保護及種質創制利用提供科學參考。
1 材料與方法
1.1 材料
分別在中國長島、榮成、即墨和韓國釜山海區采捕野生魁蚶群體,每個群體隨機選取30枚進行形態性狀和活體質量指標測定。
1.2 形態指標測定
用游標卡尺(精確到0.01 mm)常規測量魁蚶殼長(x1)、殼高(x2)、殼寬(x3)、楯面長(x4)、楯面寬(x5)、前端到腹緣距離(x6)、后端到腹緣距離(x7)、小月面長(x8)、韌帶長(x9)和放射肋寬(x10)共10個表型參數,吸干貝殼表面水分在陰暗環境下放置約30 min,用電子天平(精確到0.01 g)稱量體質量(x11)。
1.3 分析方法
采用SPSS 24.0和Excel軟件對各項表型參數進行統計分析及相關分析,為消除魁蚶規格不同對形態指標的影響,以各性狀指標的比值作為魁蚶形態的分析指標,分別為殼寬/殼長、殼高/殼長、楯面長/殼長、楯面寬/殼長、前端到腹緣/殼長、后端到腹緣/殼長、小月面長/殼長、韌帶長/殼長、放射肋寬/殼長和體質量/殼長等10個比例性狀。
用SPSS 24.0軟件對上述比例性狀進行主成分分析、聚類分析和判別分析[16]。主成分分析運用Analyze→Dimension Reduction→Factor Analysis方法對魁蚶4個群體的形態特征數據進行主成分分析,得到各主成分的特征值和貢獻率;聚類分析采用歐氏距離的最短距離法對各群體10個比例性狀的平均值進行系統聚類,繪出反映形態相似性的樹形圖;判別分析采用逐步判別法進行,選擇對4個群體形態差異貢獻較大的參數建立4個群體的判別函數。
2 結 果
2.1 表型參數統計量
4個群體魁蚶形態性狀存在一定差異,其中榮成群體的殼寬/殼長最小(0.643),說明其殼型隆起程度最低,而長島群體的殼寬/殼長最大(0.668),說明其殼型隆起程度最高;從殼高/殼長可以看出,榮成群體(0.799)與其他3個群體相比殼型較圓;體質量/殼長的比較結果表明,釜山群體(0.711)個體體質量最小,長島群體(1.411)最大,幾乎是釜山群體的2倍(表1)。
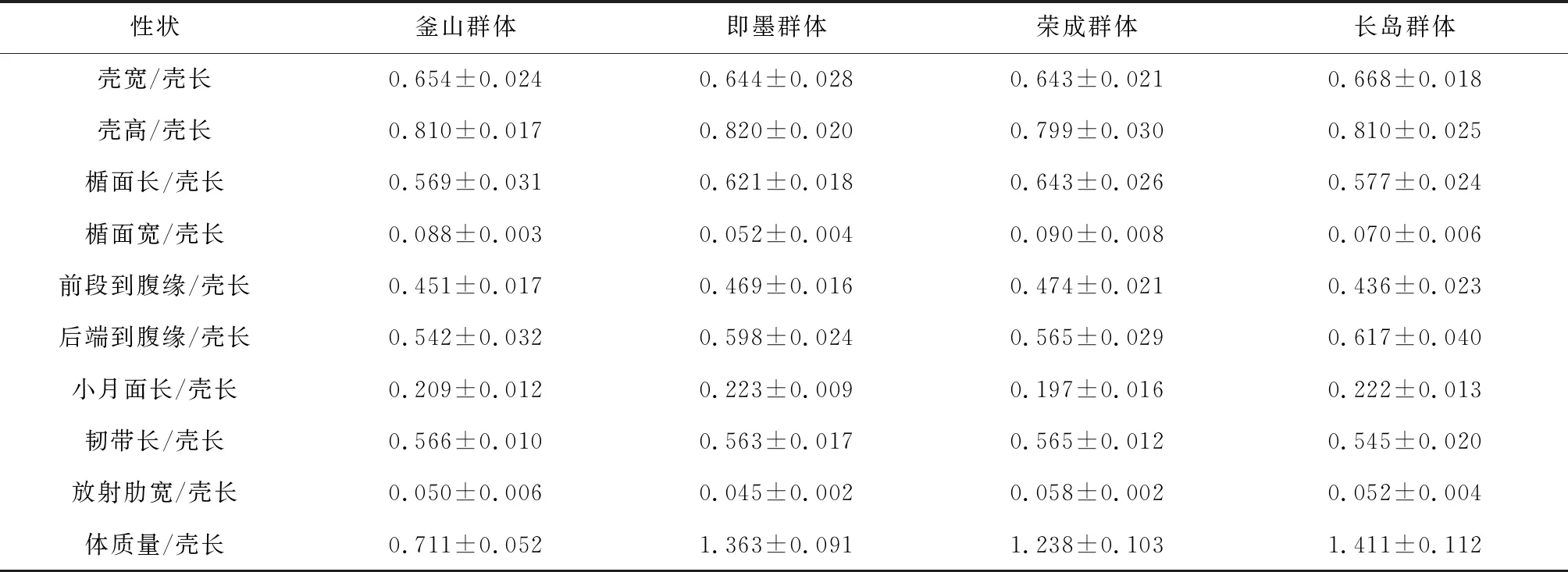
表1 4個地理群體魁蚶主要形態指標的統計值(n=30)
2.2 各性狀間的相關系數
不同魁蚶群體各性狀之間相關系數變化存在差異,其中3個中國群體除小月面長外,其他性狀間的表型相關性均達到顯著或極顯著水平;而韓國釜山群體各性狀之間相關系數變化較大,有些性狀之間未達到顯著相關水平,其中體質量與殼長、體質量與殼高極顯著相關,體質量與韌帶長顯著相關(表2)。
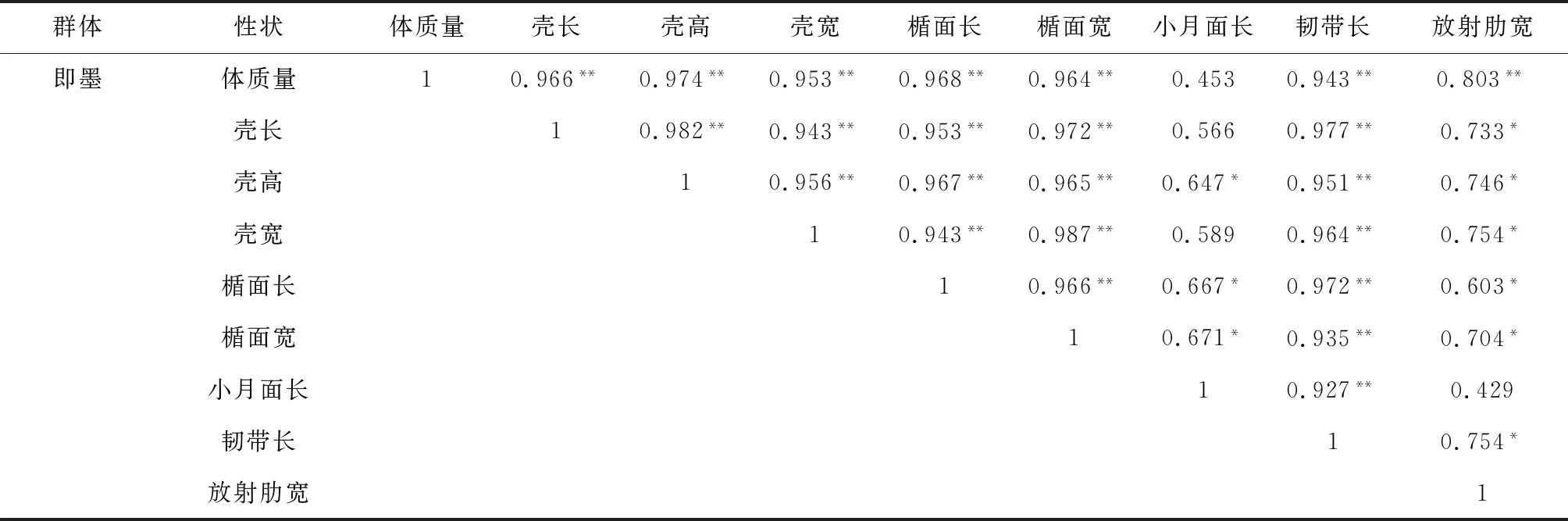
表2 各測量性狀之間的相關關系
續表2
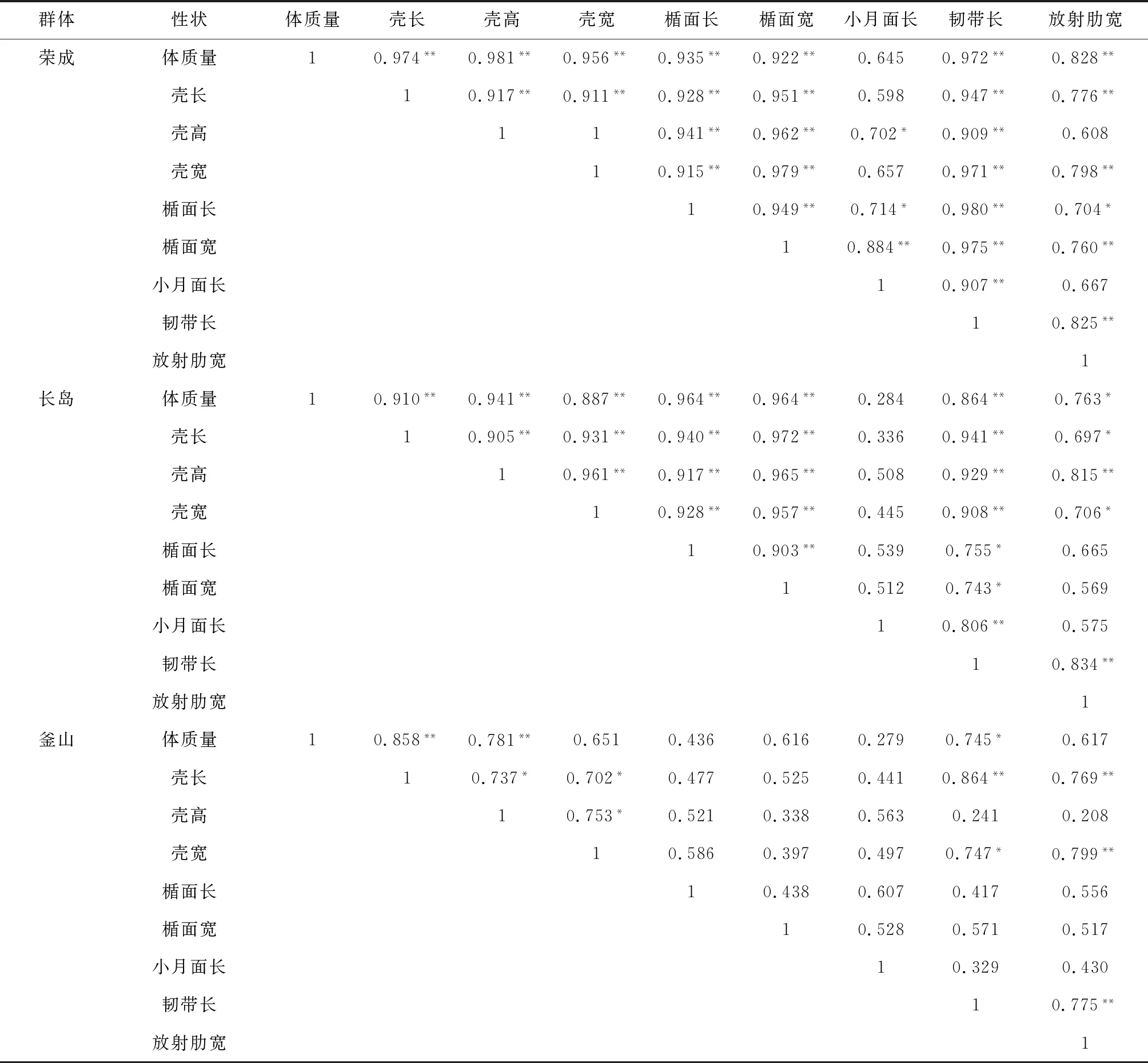
群體性狀體質量殼長殼高殼寬楯面長楯面寬小月面長韌帶長放射肋寬榮成體質量10.974**0.981**0.956**0.935**0.922**0.6450.972**0.828**殼長10.917**0.911**0.928**0.951**0.5980.947**0.776**殼高110.941**0.962**0.702*0.909**0.608殼寬10.915**0.979**0.6570.971**0.798**楯面長10.949**0.714*0.980**0.704*楯面寬10.884**0.975**0.760**小月面長10.907**0.667韌帶長10.825**放射肋寬1長島體質量10.910**0.941**0.887**0.964**0.964**0.2840.864**0.763*殼長10.905**0.931**0.940**0.972**0.3360.941**0.697*殼高10.961**0.917**0.965**0.5080.929**0.815**殼寬10.928**0.957**0.4450.908**0.706*楯面長10.903**0.5390.755*0.665楯面寬10.5120.743*0.569小月面長10.806**0.575韌帶長10.834**放射肋寬1釜山體質量10.858**0.781**0.6510.4360.6160.2790.745*0.617殼長10.737*0.702*0.4770.5250.4410.864**0.769**殼高10.753*0.5210.3380.5630.2410.208殼寬10.5860.3970.4970.747*0.799**楯面長10.4380.6070.4170.556楯面寬10.5280.5710.517小月面長10.3290.430韌帶長10.775**放射肋寬1
2.3 主成分分析
分別對4個魁蚶群體所有樣本的9個比例性狀進行主成分分析,各獲得前3個主成分第1主成分、第2主成分和第3主成分,第1主成分貢獻率為27.989%,第2主成分貢獻率為20.742%,第3主成分貢獻率為13.612%,累積貢獻率為62.342%。3個主成分可解釋不同群體之間的形態差異的62.343%,在第1主成分中,后端到前緣的影響最大,其貢獻率為79.6%;第2主成分中殼高和殼寬影響最大,其貢獻率分別為78.4%和75.3%。
2.4 聚類分析
4個魁蚶群體所有樣本的10個比例性狀參數的平均值聚類分析結果見圖1。結果表明,4個魁蚶群體聚為2支,其中榮成種群與即墨種群距離最短,形態最為接近,長島群體、即墨群體、榮成群體的趨異程度逐漸增加,而韓國釜山群體與前三者的趨異程度最大。
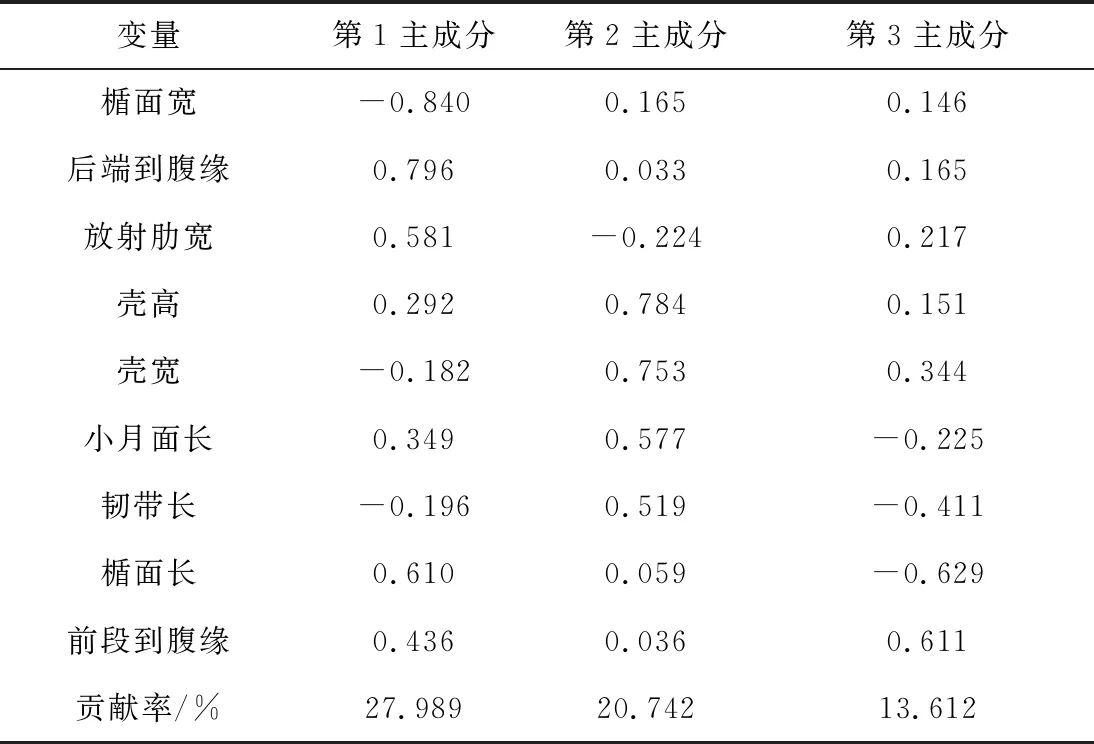
表3 不同魁蚶群體形態特征因子負荷矩陣及主成分貢獻率
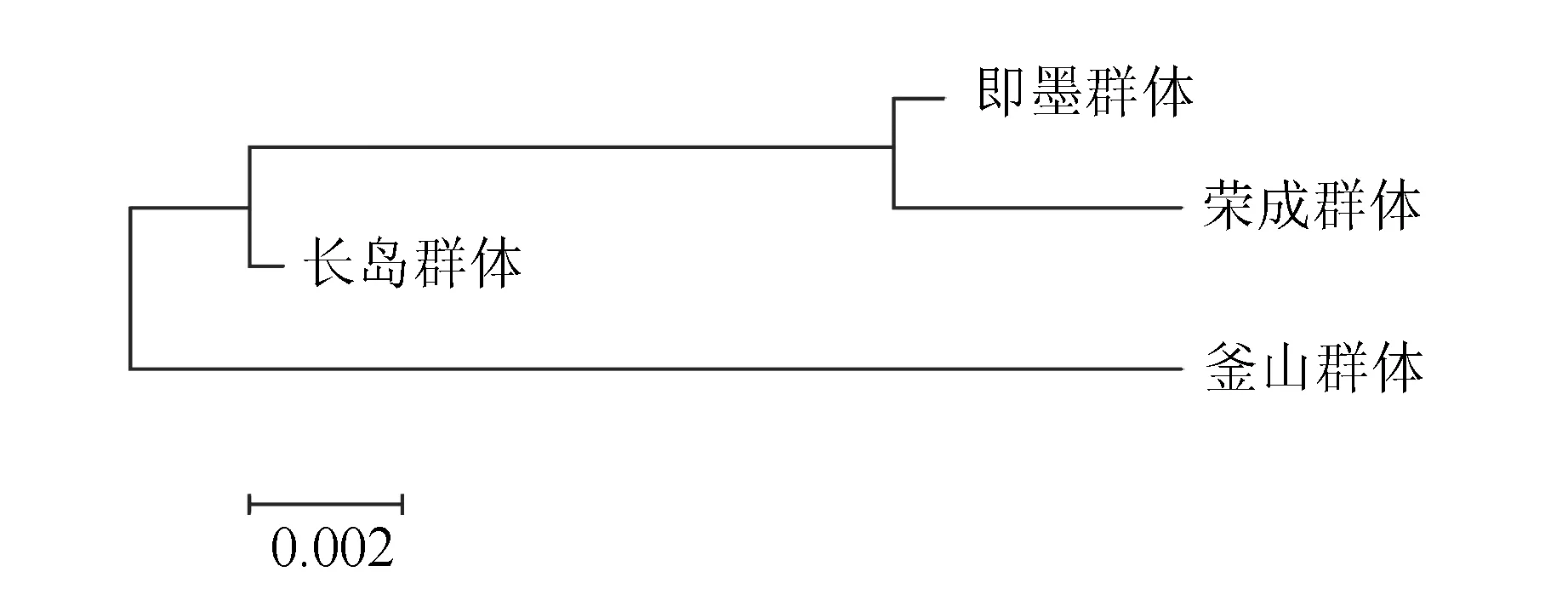
圖1 4種魁蚶的聚類分析
2.5 判別分析
通過逐步判別分析,篩選出了9個比例性狀的特征值建立了4個魁蚶群體的判別函數,判別公式如下:
釜山=235.243x1+504.017x2+510.319x3+352.362x4-171.897x5+298.589x6+559.341x7+885.369x8+1604.906x9-914.094
長島=214.747x1+510.804x2+573.793x3-110.000x4-156.232x5+316.199x6+602.823x7+909.157x8-914.094x9-975.344
即墨=289.123x1+479.964x2+123.578x3+432.678x4-786.245x5+276.471x6+576.881x7+885.369x8+1309.445x9-897.246
榮成=253.747x1+578.234x2+567.903x3-117.543x4-243.562x5+782.246x6+562.783x7+108.465x8-1064.032x9- 934.122
對各個地區采集的樣本按判別函數進行預測分類,判別結果表明,釜山、長島、即墨、榮成魁蚶判別分析正確率為90.0%、96.7%、86.9%和87.7%。
3 討 論
3.1 多元分析方法在貝類形態差異分析中的應用
多元統計分析方法是統計學中常見的綜合分析方法,是研究多變量總體統計問題的理論和方法,包括相關性分析、主成分分析、聚類分析、判別分析等多種實用可行的方法[17-22]。目前,多元統計方法在貝類等水產動物的分類及鑒定中已得到廣泛應用,如牛泓博等[9]利用主成分分析、聚類分析等多元分析方法對遼寧沿海菲律賓蛤仔8個野生群體進行形態學比較分析,發現旅順群體和錦州群體形態最為接近,而與黑石礁群體差異較大,營口群體的趨異程度最大,長興島群體、金州群體和海洋島群體的趨異程度居中。楊陽[23]對嵊泗養殖海域3種貽貝(Mytilus)貝體性狀的比較研究發現,“雜交貽貝”的外部形態特征介于紫貽貝(M.galloprovincialis)與厚殼貽貝(M.coruscus)之間,更似紫貽貝;采用逐步判別法,以Fisher分類函數方程組可清晰區分3種貽貝,綜合判別準確率為96.4%。王慶恒等[11]運用主成分分析、判別分析和聚類分析等多元統計分析方法,比較了廣西北海、廣東湛江和汕尾的3個翡翠貽貝野生種群的形態性狀差異,其中主成分分析累計貢獻率為88.704%,3個種群判別函數綜合判別率為79.7%;聚類分析顯示,北海種群與汕尾種群形態較為接近,湛江種群趨異程度最大。對不同地理群體魁蚶的形態差異分析也取得了一定進展,梁超等[24]運用多變量形態度量學分析方法對韓國統營、山東黃島、山東蓬萊、江蘇前三島4個魁蚶種群間的形態變異進行了研究,聚類分析結果表明,黃島和前三島兩個種群形態最為接近,蓬萊種群與其他3個種群距離最遠;主成分分析構建的3個主成分貢獻率分別為33.366%、20.407%和15.422%,累計方差貢獻率為69.195%;建立了魁蚶4個種群的判別函數,其判別準確率分別為韓國通營96.4%、山東蓬萊100%、山東黃島84.3%和江蘇前三島95.6%。利用多元分析方法可揭示不同群體貝類的形態差異,可為貝類地理群體識別、種質資源保護與恢復提供重要依據。
3.2 4個魁蚶群體形態差異的比較分析
本研究發現,3個中國魁蚶群體除小月面外,其他性狀與體質量的相關性均達到顯著或極顯著水平,而韓國釜山群體則只有殼長、殼高和韌帶長與體質量呈顯著或極顯著相關,該結果與之前的研究結果類似[24-25]。推測其原因可能與魁蚶的生活習性和棲息環境的不同有關,魁蚶營埋棲性生活,遷移流動性較小,其外部形態除受基因等遺傳因素影響外,也受其生活環境的溫度、鹽度、底質等理化因素的影響。
主成分分析、聚類分析和判別分析的結果一致表明,榮成、即墨、長島魁蚶群體的形態變異差異較小,與釜山群體的形態變異差異較大。其中主成分分析中第3主成分中前段到腹緣距離影響最大,其貢獻率為61.1%,第1主成分上,釜山群體和榮成群體的差異較小,而在第2主成分上,兩者差異較大。由于對第2主成分影響較大的主要是殼高性狀,說明長島群體和即墨群體魁蚶在這一性狀上存在較明顯差異。在第1主成分上,長島群體和榮成群體沒有明顯分開,說明兩者在幾個性狀上差異較小,而在第2主成分上,釜山群體與長島群體明顯分開,說明二者在這一性狀上存在明顯差異,長島魁蚶群體的殼頂明顯前移。聚類分析結果表明,榮成群體與即墨群體魁蚶遺傳距離最近,即墨群體、長島群體、榮成群體的趨異程度逐漸增加,而釜山群體與三者的趨異程度最大。本研究結果與之前的研究結果存在一定差別,梁超等[24]的研究結果表明,蓬萊群體與韓國通營、黃島和前三島3個群體的距離最遠,造成這種差別的原因可能為不同群體魁蚶的形態上的差異不僅僅與地理距離相關,還受其他多種因素的影響,如遺傳因素等[25]。
通過逐步判別分析,篩選出了9個性狀的特征值建立了4個魁蚶群體的判別函數,釜山、長島、即墨、榮成魁蚶群體判別分析正確率為90.0%、96.7%、86.9%和87.7%,這種差異的產生可能是由不同海區的生態環境存在著一定差異所致。山東和韓國雖然同屬溫帶水系,但是底質方面,山東沙底質較多,而韓國泥底質較多,魁蚶的生長規律不僅與水溫有關,而且其生理形狀與底質的理化性質也具有相關性。筆者認為導致魁蚶形態不同的原因也可能是對不同環境適應的結果,環境中底質是決定形態變異的主要因素之一,這種表型特征的地理變異也是長期演變的結果。
綜上,韓國魁蚶群體與中國魁蚶群體在外形上存在差異,遺傳距離明顯大于中國國內群體,利用韓國群體與中國群體魁蚶進行雜交可能產生較大的雜種優勢。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
