生長調節劑對紅豆杉內生真菌產紫杉醇的影響
2019-09-17 07:32:25趙赟鑫張歡張顏青
江蘇農業科學 2019年7期
關鍵詞:紫杉醇
趙赟鑫 張歡 張顏青
摘要:根據紫杉醇的結構特點和紅豆杉中紫杉醇的合成機制,選取4種生長調節劑,研究其對紅豆杉內生真菌合成紫杉醇的影響。結果表明,在發酵過程的第10天,補加下列任一生長調節劑,使發酵液中初始濃度分別達到水楊酸20.0 mg/L,茉莉酸甲酯100.0 μmol/L,赤霉素2.0 mg/L,矮壯素2.0 mg/L,均能提高紫杉醇產量。同時,在發酵過程的第10天補加蔗糖,在含量不高于15 g/L時,對紫杉醇的積累和菌絲體生長具有促進作用,但是高于此濃度時,菌絲體生長受抑制,紫杉醇含量降低。
關鍵詞:紅豆杉;紫杉醇;內生真菌;生長調節劑
中圖分類號: Q939.9 ?文獻標志碼: A ?文章編號:1002-1302(2019)07-0120-04
近年來,紅豆杉產紫杉醇的研究進展很快,國內外利用內生真菌產紫杉醇的研究報道逐漸增多,目前內生真菌產紫杉醇的含量普遍偏低,國內外學者也正在不斷探索能夠提高內生真菌產紫杉醇產量的途徑[1-2]。
誘導子是一種能誘導植物細胞中一種或多種反應并形成特征性自身防御反應的分子,是能夠引起植物過敏反應的物質,其在與植物的相互作用中,能夠快速、高度專一和選擇性地活化植物次級代謝途徑中特定酶的基因或改變次級代謝途徑中催化酶的酶活力,誘導植物形成新酶,進而活化特定次生代謝途徑,引起反應速率和次級代謝途徑通量的改變,從而積累特定目的次級的代謝物,因此可以利用誘導子來提高植物次生代謝產物的產量[3]。
紫杉醇是紅豆杉屬植物及其內生真菌產生的次級代謝產物,非生物或生物誘導子可誘導細胞產生抗逆反應,啟動次級代謝途徑,進而合成大量的紫杉醇。通常,誘導子對紅豆杉細胞的生長均有抑制作用,因而添加時間、添加濃度對誘導子作用的最終效果非常關鍵[4]。添加代謝抑制劑,以抑制一些與紫杉醇生物合成無關的次級代謝途徑為目的,通過添加某些代謝旁路抑制劑,將細胞的物質和能量導向紫杉醇生物合成途徑,從而提高紫杉醇的產量。當然,代謝抑制劑也存在一個最佳添加濃度和最佳添加時間[5]。
大量研究表明,在對數生長末期添加前體物質、誘導子及代謝抑制劑的效果最好,因為此時細胞已得到較好的增殖,而且在此階段,植物細胞接受生長調節物質的信號能力最強[6]。
作者采用課題組自行分離的紫杉醇高產菌株綠僵菌(Metarhizium anisopliae) LB-10[7],在對其培養基組成和配比[8]、發酵條件研究[9]的基礎上,根據紫杉醇的結構特點和紅豆杉中紫杉醇的合成機制,選取4種生長調節劑,研究其對紅豆杉內生真菌合成紫杉醇的影響。
1 材料與方法
1.1 材料
1.1.1 菌株 紅豆杉內生真菌Metarhizium anisopliae LB-10是分離自陜西省漢中市留壩縣野生紅豆杉的高產紫杉醇內生真菌。
1.1.2 培養基 斜面培養基:馬鈴薯葡萄糖瓊脂(PDA)培養基:馬鈴薯200 g,葡萄糖20 g,瓊脂15~20 g,水1 L,pH值自然;種子培養基:馬鈴薯葡萄糖肉湯(PDB)培養基:馬鈴薯200 g,葡萄糖20.0 g,水1.0 L,pH值6.0~8.0;發酵液:葡萄糖50.0 g/L,NH4NO3 6.0 g/L,無水MgSO4 0.3 g/L,KH2PO4 0.5 g/L,維生素B1 5.0×10-2 g/L。
1.1.3 藥品與試劑 紫杉醇標準品(≥98%)、乙酸乙酯、甲醇、葡萄糖、NH4NO3、KH2PO4、MgSO4·7H2O、苯甲酸鈉、苯丙氨酸、乙酸鈉、酪氨酸。
1.1.4 主要儀器 高效液相色譜儀(LC 2000),旋轉蒸發儀(RV-10,IKA),電子天平(TB-214),雙層恒溫干燥培養振蕩器(ZHWY-2102C),人工氣候箱(LRH-250-G-S),數控超聲波清洗儀(KQ-5200-DE)。
1.2 方法
1.2.1 培養方法 種子液培養方法:在新鮮斜面上取 5 mm×5 mm大小的已純化的菌塊,接種到裝有50 mL發酵培養基的250 mL三角瓶中,于25 ℃、180 r/min搖床培養 3 d。
發酵培養:將培養好的種子液混勻,按3%接種量接種到裝有330 mL發酵培養基的500 mL三角瓶中,每次提取所用發酵液的量為1 000 mL,于28 ℃、180 r/min搖床培養10 d。
1.2.2 紫杉醇樣品的提取 通過對培養10 d的發酵液進行抽濾,使其分離為菌液和菌絲體2個部分,菌絲用乙酸乙酯在超聲條件下萃取,菌液用乙酸乙酯通過分液漏斗萃取,分別重復3次,合并收集到的乙酸乙酯相,并用雙層濾紙過濾,濾液在40 ℃下旋轉蒸發至干,樣品用甲醇溶解并且定容至 10.0 mL,檢測。
1.2.3 紫杉醇的HPLC檢測 紫杉醇標準品用甲醇定容至10.0 mL,配制成0.01~0.16 mg/mL的濃度梯度,繪制標準曲線。精確稱取紫杉醇標準品4.0 mg,甲醇定容至10.0 mL,從而得到母液濃度為0.4 mg/mL,將母液依次稀釋成濃度梯度為0.01、0.02、0.04、0.08、0.16 mg/mL,取20.0 μL進樣(N=5),得到回歸方程為y=6.087 8+3 589.391 3x,r=0.999 222。建立標準曲線,如圖1所示,采用外標法測定內生真菌發酵產物的乙酸乙酯抽提物中紫杉醇的含量。
色譜條件:水-甲醇-乙腈(體積比33 ∶ 35 ∶ 32)為流動相,檢測波長為228 nm,流速為1.0 mL/min,進樣量為 20.0 μL,柱溫為室溫,色譜柱:C18(4.6 mm×150 mm)。
紫杉醇含量的計算公式:發酵液中紫杉醇含量(μg/L)=[甲醇中紫杉醇含量(mg/mL)×溶解提取物所用甲醇體積(mL)×106]/提取時所取發酵液體積(mL)。
每次提取的紫杉醇均做3個平行樣,分別經HPLC測定并通過公式計算發酵液中紫杉醇的含量,求其平均值。
1.2.4 生物量的測定 取一定體積的發酵液4 800 r/min離心20 min,菌絲體用蒸餾水洗滌2次,收集菌絲體,置于80 ℃烘箱中烘干至恒質量,稱量,計算菌絲體生物量。
1.2.5 添加誘導子的研究方法 以水楊酸為誘導子,采用蒸餾水作為助溶劑,在菌株LB-10對數生長末期即第10天,添加水楊酸水溶液,使發酵液中其初始濃度分別達到5.0、10.0、20.0、40.0、80.0 mg/L,并作空白對照,研究水楊酸對菌株LB-10菌絲體生物量和紫杉醇生物合成的影響。
以茉莉酸甲酯為誘導子,采用乙醇作為助溶劑,在菌株LB-10對數生長末期即第10天,添加茉莉酸甲酯醇溶液,使發酵液中其初始濃度分別達到50.0、75.0、100.0、125.0、150.0 μmol/L,并作空白對照,研究茉莉酸甲酯對菌株LB-10菌絲體生物量和紫杉醇生物合成的影響。
1.2.6 添加抑制劑的研究方法 以赤霉素為抑制劑,采用蒸餾水為助溶劑,在菌株LB-10對數生長末期即第10天,添加赤霉素水溶液,使發酵液中其初始濃度分別達到0.5、1.0、2.0、4.0、8.0 mg/L,并作空白對照,研究赤霉素對菌株LB-10菌絲體生物量和紫杉醇生物合成的影響。
以矮壯素為抑制劑,采用蒸餾水為助溶劑,在菌株LB-10對數生長末期即第10天,添加矮壯素水溶液,使發酵液中其初始濃度分別達到0.5、1.0、2.0、4.0、8.0 mg/L,并作空白對照,研究矮壯素對菌株LB-10菌絲體生物量和紫杉醇生物合成的影響。
1.2.7 培養基中補加蔗糖的研究方法 在菌株LB-10對數生長末期即第10天,添加蔗糖水溶液,使發酵液中其初始濃度分別達到5.0、10.0、15.0、20.0、25.0 g/L,并作空白對照,研究補加蔗糖對菌株LB-10菌絲體生物量和紫杉醇生物合成的影響。
2 結果與分析
2.1 誘導子的影響
2.1.1 水楊酸的影響 水楊酸是酚類化合物,為白色結晶性粉末,易溶于水,微溶于乙醇和乙醚。主要作為植物抗病反應的重要信號分子,能夠激活多種與抗病相關的植物防御機制。涉及并參與系統獲得抗性反應和植物過敏反應,在植物抗病反應中起重要的作用。水楊酸可分為內源水楊酸和外源水楊酸,前者能夠誘導與抗病蛋白有關基因的表達,而后者則主要誘導防御反應的產生[10]。試驗結果如圖2所示。
由圖2可知,在0~20 mg/L濃度范圍,水楊酸對紫杉醇合成有明顯的促進作用,在20 mg/L時紫杉醇含量達到 986.1 μg/L,繼續增加濃度后紫杉醇含量開始下降。而且水楊酸含量在高于5.0 mg/L后,對菌絲體生長具有明顯的抑制作用。由于水楊酸誘導細胞產生過敏反應,能夠提高與次生代謝物合成相關酶的活性[11],從而促進細胞積累紫杉醇,所以水楊酸對紫杉醇合成具有顯著性影響,但過高濃度的水楊酸會嚴重影響細胞的生長,反而使紫杉醇含量下降,因而須選擇合適的水楊酸添加劑量。
2.1.2 茉莉酸甲酯的影響 茉莉酸類作為與損傷相關的植物激素和信號分子,在自然界廣泛存在,其主要代表物為茉莉酸和茉莉酸甲酯,茉莉酸甲酯的外源應用能夠激發防御植物基因的表達,誘導植物的化學防御,產生與機械損傷和昆蟲取食相似的效果[12]。由圖3可知,茉莉酸甲酯濃度在0~100 μmol/L 范圍內時,隨著濃度的升高,紫杉醇含量也逐漸增高,在添加 100 μmol/L 時,紫杉醇含量達到996.2 μg/L,但繼續提高濃度,生物合成紫杉醇含量下降,而且在試驗范圍內,茉莉酸甲酯對菌絲體生長有明顯的抑制作用。在茉莉酸甲酯的誘導作用下,菌絲體生長和紫杉醇的合成呈負相關關系。由于茉莉酸甲酯可激活特定防御基因的轉錄和翻譯[13],最終促進次級代謝產物的合成,所以茉莉酸甲酯對紫杉醇合成是一種非常有效的誘導子。但由于茉莉酸甲酯會影響菌絲體的生長,反而使紫杉醇含量下降,因而須選擇合適的茉莉酸甲酯添加劑量。
2.2 抑制劑的影響
2.2.1 赤霉素的影響 赤霉素是雙萜類化合物,化學結構比較復雜,易溶于水,其最突出的生理效應是能顯著地促進莖的伸長、能夠引起某些植物形成單性果實和誘導長日植物在短日條件下開花,對某些生理現象有時有抑制作用[14]。由圖4可知,在0~2.0 mg/L濃度范圍內,赤霉素對紫杉醇合成有一定的促進作用,在2.0 mg/L時紫杉醇含量達到 1 015.2 μg/L,繼續增加濃度后紫杉醇含量逐漸下降,而且赤霉素在低濃度下對菌絲體生長具有一定的促進作用,但當濃度高于0.5 mg/L時,對生物量的積累具有明顯的抑制作用。由于赤霉素類化合物與紫杉醇具有一個同樣的赤霉烷二萜母核,所以甲羥戊酸途徑是其合成途徑,并且也是由牻牛兒基焦磷酸(GGPP)環合而成[15]。在甲羥戊酸途徑中的許多酶活性是受其代謝產物的反饋抑制,而赤霉素也有相似的作用,那么添加的赤霉素可能會抑制由GGPP合成赤霉素類化合物,為紫杉醇的合成提供更多的基礎物質,從而提高紫杉醇的生物合成量;由于赤霉素濃度的提高,對菌絲體生長具有明顯的抑制作用,導致紫杉醇含量下降。
2.2.2 矮壯素的影響 矮壯素即氯化氯膽堿,是一種能夠與赤霉素發生拮抗作用的生長調節物質,屬于甾體合成代謝抑制劑[16],其生理作用是控制植株的營養生長和生殖生長,使植株的間節縮短、矮壯并抗倒伏。由圖5可知,在0~2.0 mg/L 濃度范圍內,矮壯素對紫杉醇合成有一定的促進作用,在2.0 mg/L時紫杉醇含量最高,達到1 032.5 μg/L,繼續增加濃度后紫杉醇含量逐漸下降。而且還發現矮壯素對菌株LB-10的菌絲體生長有明顯的抑制作用。由于矮壯素可以抑制赤霉素合成途徑中的環化步驟,此作用與上述假設的赤霉素反饋抑制作用基本相同。同時,矮壯素能抑制固醇的合成,而固醇抑制甲羥戊酸途徑中限速酶3-羥基-3-甲基戊二酸單酰輔酶A還原酶(HMG-CoA)的活性[17],所以添加矮壯素能夠間接地提高HMG-CoA的活性,從而促進甲羥戊酸途徑的代謝,間接地促進紫杉烷類化合物和紫杉醇的合成。隨著矮壯素濃度的增加,會對菌絲體生長產生明顯地抑制,從而限制了紫杉醇含量的增加。
2.3 培養基補加蔗糖的研究
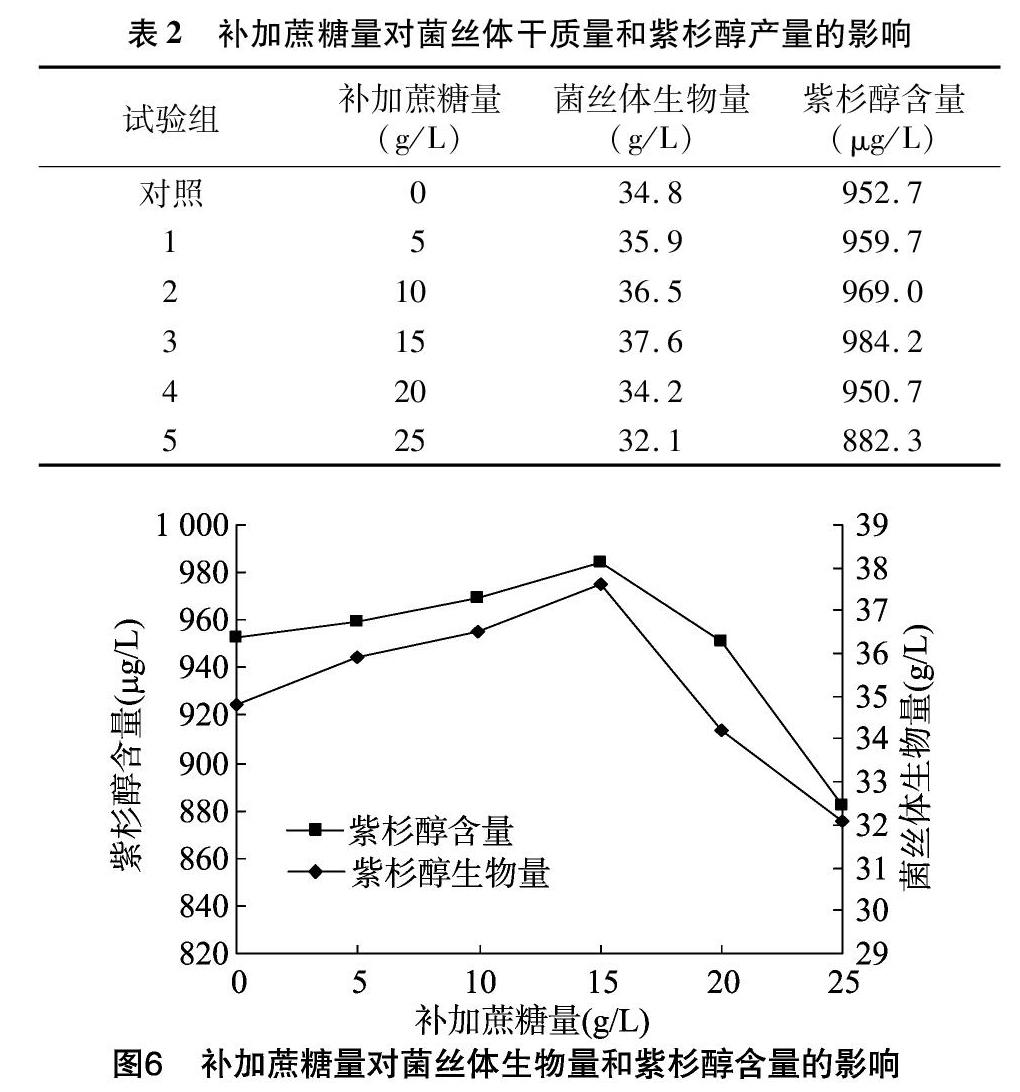
微生物培養基中經常使用蔗糖作為碳源,它的主要作用就是用于合成微生物細胞物質以及提供能量,蔗糖在微生物體內經常被降解成果糖和葡萄糖,其中果糖對細胞培養后期合成紫杉醇有促進作用,而葡萄糖主要是在細胞的前期生長中利于獲得大量的細胞。在培養到第10天時,營養物質基本已消耗殆盡,考慮在此時添加不同濃度的蔗糖,使其發酵液中的蔗糖濃度范圍在0~25 g/L之間。由表2、圖6可知,在發酵液中蔗糖濃度為15 g/L時,菌體能快速生長和繁殖,紫杉醇產量和生物量最高,其中紫杉醇含量達到984.2 μg/L,而第5試驗組的發酵液與對照組相比,其發酵液的黏度較大,呈糊狀,紫杉醇含量也比較低。間隔補加蔗糖,隨著蔗糖濃度的提高,其滲透壓相應地提高,導致膜蛋白降解,從而增加了菌體的通透性,使得代謝產物更多地向胞外釋放,促進代謝產物的生物合成速率,進而提高其產量,但在后期,由于過高的蔗糖濃度和大量的菌絲體量導致發酵液黏度明顯增加,使菌絲體呼吸困難,從而抑制了紫杉醇的生物合成量。因此補加的蔗糖量一定要根據菌株的特性和發酵條件適量添加。
3 結論與討論
代謝調控是通過定向改良細胞的代謝特性,對特定的生化反應進行修飾,從而實現合成新產物或提高目的產物的產率以及減少副產物的產率。在正常的生理條件下,微生物總是通過協調系統最經濟地吸收和利用營養物質,用于合成細胞結構,進行生長和繁殖,一般不積累中間代謝產物。代謝人工調控就是打破微生物的代謝控制體系,使代謝朝著人們希望的方向進行。因此,在了解紫杉醇生物合成途徑和催化各步反應酶的特性后,有目的地控制培養條件,通過添加重要的前體物質、誘導子及抑制劑,對關鍵酶的活性進行調節,抑制其他無用的旁路途徑,是完全可以提高紫杉醇產量的。
本研究根據紅豆杉紫杉醇的合成機制和紫杉醇的結構特點,選取水楊酸、茉莉酸甲酯、赤霉素和矮壯素分別作為生長調節劑,研究其對Metarhizium anisopliae LB-10合成紫杉醇的影響。結果表明,在發酵過程中第10天補加下列任一生長調節劑,使發酵液中初始濃度分別達到水楊酸20.0 mg/L、茉莉酸甲酯100.0 μmol/L、赤霉素2.0 mg/L、矮壯素2.0 mg/L,均能提高紫杉醇產量。
通過研究在液體發酵過程中補加蔗糖對菌株LB-10的紫杉醇含量和菌絲體生物量的影響。結果顯示,蔗糖含量在不高于15 g/L時,對紫杉醇積累和菌絲體生長具有促進作用,但是高于此濃度時,菌絲體生長受抑制,進而降低了紫杉醇含量。
紫杉醇具有細胞毒害作用[18],通常不會大量地合成,只有在特殊環境脅迫條件下,初生代謝受到抑制,次生代謝產物才會大量合成。在試驗過程中,筆者所在課題組發現添加某些生長調節劑,在紫杉醇產量較高時,生物量卻較低。這就造成紫杉醇生物合成和菌絲體生長之間的矛盾,所以找到二者最佳平衡點是接下來須要解決的問題,也是實現工業化生產的必要條件。
參考文獻:
[1]張 歡,趙赟鑫,高 文,等. 紫杉醇生產現狀的分析與對策[J]. 中國現代中藥,2016,18(1):126-130.
[2]趙赟鑫,高 文,張 歡,等. 提高紅豆杉內生真菌及紫杉醇含量的途經分析[J]. 陜西農業科學,2015,61(10):13-17.
[3]施中東,未作君,元英進. 南方紅豆杉細胞培養合成紫杉醇誘導子濃度的優化[J]. 天然產物研究與開發,2000,12(4):36-40.
[4]Ketehum R E B,Gibson D M,Croteau R B,et al. The kineties of taxoid accumulation in cell suspension cultures of taxus following elieitation with methyl jasmonate[J]. Bioteehnol Bioeng,1999,62(1):97-105.
[5]梅興國,吳奇君,江振然,等. 東北紅豆杉細胞培養生產紫杉醇的調控研究Ⅱ——交互作用對東北紅豆杉細胞培養生產紫杉醇的影響分析[J]. 生命科學研究,2002,6(2):152-155.
[6]周忠強,梅興國,吳奇君. 前體、誘導子及抑制劑對細胞培養生產紫杉醇的調節作用[J]. 天然產物研究與開發,2002,14(2):19-21.
[7]耿 直,劉開輝,趙赟鑫,等. 一株產紫杉醇中國紅豆杉內生真菌的分離和鑒定[J]. 微生物學通報,2010,37(2):199-203.
[8]趙赟鑫,張 歡,鄧百萬,等. 紅豆杉內生真菌產紫杉醇的培養基優化[J]. 江蘇農業科學,2014,42(11):389-392.
[9]張 歡,趙赟鑫,李昭瑩,等. 紅豆杉內生真菌產紫杉醇的發酵條件優化研究[J]. 江蘇農業科學,2018,46(21):306-309.
[10]鄭愛珍,王啟明. 水楊酸與植物逆境脅迫[J]. 安徽農業科學,2006,34(5):844-845.
[11]梅國興,張舟寧,蘇湘鄂,等. 水楊酸對紅豆杉細胞的誘導作用[J]. 生物技術,2000,10(6):90-92.
[12]薛仁鎬,金圣愛. 茉莉酸甲酯:一種重要的植物信號轉導分子[J]. 生物技術通訊,2006,17(6):985-988.
[13]余龍江,朱 敏,周 瑩,等. 茉莉酸甲酯對紫杉醇生物合成的誘導作用[J]. 天然產物研究與開發,1998,11(5):1-7.
[14]黃先忠,蔣才富,廖立力,等. 赤霉素作用機理的分子基礎與調控模式研究[J]. 植物學通報,2006,23(5):499-510.
[15]盧利方,馮仁軍,張銀東. 赤霉素信號轉導的分子生物學研究進展[J]. 生命科學研究,2005,9(4):66-71.
[16]周忠強,梅國興,吳奇君. 前體、誘導子及抑制劑對細胞培養生產紫杉醇的調節作用[J]. 天然產物研究與開發,2001,14(2):19-21.
[17]Hamill J D,Parr A J,Rhodes M J C,et al. New routes to plant secondary products[J]. Bio Technology,1987,5(8):800-804.
[18]李宗霆. 植物激素及其免疫檢測技術[M]. 南京:江蘇科學技術出版社,1996.竇允清,王振華,張金珠,等. 水肥耦合對滴灌加工番茄生理生長及產量的影響[J]. 江蘇農業科學,2019,47(7):124-129.
猜你喜歡
世界科學技術-中醫藥現代化(2022年2期)2022-05-25 13:17:14
昆明醫科大學學報(2021年12期)2021-12-30 07:00:10
中國臨床醫學影像雜志(2021年6期)2021-08-14 02:21:56
中成藥(2018年12期)2018-12-29 12:25:22
實用口腔醫學雜志(2017年6期)2017-09-19 02:51:06
中外醫療(2016年15期)2016-12-01 04:25:50
哈爾濱醫藥(2015年2期)2015-12-01 03:57:41
原子與分子物理學報(2015年3期)2015-11-24 12:49:34
醫學研究雜志(2015年5期)2015-06-10 06:43:26
安徽醫藥(2014年4期)2014-03-20 13:12:25
