干旱與復水對馬鈴薯塊莖膨大期碳氮轉運的影響
2019-09-16 02:19:36喬煥煥李紅兵鄭太波鄧西平
干旱地區農業研究 2019年4期
喬煥煥, 李紅兵, 鄭太波, 鄧西平
1.西北農林科技大學生命科學學院,陜西 楊凌 712100;2.黃土高原土壤侵蝕與旱地農業國家重點實驗室,西北農林科技大學水土保持研究所,陜西 楊凌 712100;3.延安市農科所,陜西 延安 716000)
馬鈴薯(SolanumtuberosumL.)栽培面積廣,是世界上非常重要的糧食作物之一。馬鈴薯是我國第四大主要糧食作物。馬鈴薯通常種植于降雨量不穩定的干旱和半干旱地區,而馬鈴薯是典型的喜濕作物,其根系較淺,對水分虧缺非常敏感,因而其生長與生產力非常容易受到水分脅迫的影響[1]。馬鈴薯生長周期中,塊莖膨大期是塊莖增長最為關鍵的時期,此時的耗水量占全生育期的45%~50%以上,是全生育期中需水量最多的時期,也是對水分最為敏感的時期[2]。研究發現,馬鈴薯塊莖形成期缺水,莖葉生長受到限制,根系的伸展受阻,降低馬鈴薯單株塊莖數[3]。
自然條件下的植物經常處于干濕交替循環變化的環境中[1]。植物對干旱脅迫的抵抗能力和旱后復水的快速恢復能力是植物抗旱能力的表現,其中旱后復水的恢復能力在植物抗旱中的意義也非常重要,因為旱后復水經常使植物的生理生態功能得到一定程度的恢復[4]。近年來關于干旱脅迫及旱后復水處理影響植物碳氮物質含量及其關鍵酶活性的研究受到關注。如干旱脅迫影響煙草光合作用,導致煙葉早衰,復水能緩解干旱脅迫導致的煙葉早衰;干旱脅迫使煙葉碳氮代謝關鍵酶(硝酸還原酶、轉化酶和淀粉酶)活性減弱,而復水在一定程度上能恢復淀粉酶活性[5]。陳朝應[6]等研究發現,干旱脅迫導致烤煙總糖與淀粉含量降低,碳代謝過程受到抑制,但總氮與煙堿含量呈增加趨勢,氮代謝增強,碳氮失衡;復水后,碳氮化合物含量都發生了相應變化,烤煙碳氮失衡狀況有所緩和。另外,有研究表明,干旱脅迫導致玉米碳氮比失衡,但復水后碳氮比并沒有快速恢復到正常水平,而是持續了較長時間,恢復速度較慢[7]。
目前對馬鈴薯的研究多關注干旱脅迫與復水[8-9]、激素[10]、氮素[11]等處理對馬鈴薯不同生育期生長發育及生理代謝等的影響,而干旱脅迫尤其是旱后復水對馬鈴薯塊莖膨大期碳氮轉運的影響鮮有報道。本研究以冀張薯8號馬鈴薯組培苗為材料,通過盆栽控水試驗,研究干旱脅迫和復水對馬鈴薯碳氮轉運過程中碳氮物質含量及其代謝過程中關鍵酶活性的影響,以期揭示馬鈴薯葉片和塊莖中碳氮物質轉運對干旱復水的響應機制,為干旱半干旱地區馬鈴薯高產栽培提供理論依據。
1 材料與方法
1.1 供試材料
供試馬鈴薯為陜北普遍種植的品種冀張薯8號,由延安市農科所薯類研究組提供;供試土壤為陜北黃綿土。
1.2 試驗設計
于2017年在西北農林科技大學水土保持研究所干旱大廳氣候室內進行盆栽試驗。土壤含有機質3 g·kg-1、全氮0.2 g·kg-1、全磷0.59 g·kg-1、全鉀18.7 g·kg-1、速效磷4 mg·kg-1、速效鉀175.8 mg·kg-1。盆缽上徑32 cm,底徑22 cm,高27 cm,每盆裝土10 kg ,每盆土中分別施尿素6.42 g(純氮0.3 g·kg-1),硫酸鉀5.55 g(K2O 0.3 g·kg-1),過磷酸鈣5.47 g(P2O50.2 g·kg-1),土壤經自然風干、過篩后裝盆。供試組培苗于3月23日在玻璃瓶中繼代培養,置于人工培養室,每天光周期為光/暗:14 h/10 h,光照強度 750 μmol·m-2·s-1,晝夜溫度為20℃/18℃;4月12日開始于紙杯中煉苗,置于人工氣候室培養,培養條件同上;5月9日挑選長勢一致的健壯苗移栽于盆缽中,每盆一株。移栽一周后,每盆覆土1 kg,現花蕾時,為防止馬鈴薯匍匐莖露出地面而影響馬鈴薯正常生長發育,每盆再覆土2 kg。
試驗設干旱、干旱-復水和正常供水3個處理,干旱與干旱-復水各設置10個重復,正常供水30個重復,共50盆。土壤水分設兩個水平:正常供水的土壤含水量為土壤最大田間持水量(凈含水量為22%)的80%;干旱脅迫的土壤含水量為土壤最大田間持水量的40%。待植株開花一周時(6月24日,馬鈴薯塊莖膨大期)開始試驗處理(開花數量達70%),盆栽稱重法控制土壤含水量,每天18∶00左右補水。
試驗于7月1日開始采樣,取樣時間為上午10∶00 左右。取樣時取每株第三或第四功能葉,立即于液氮冷凍后,-80℃冰箱中保存;將塊莖挖出,清洗干凈,切成小塊,同樣于液氮中冷凍后,存于-80℃冰箱中,用于測定可溶性糖、淀粉、可溶性蛋白等物質含量及酶活性,取樣時間見表1。
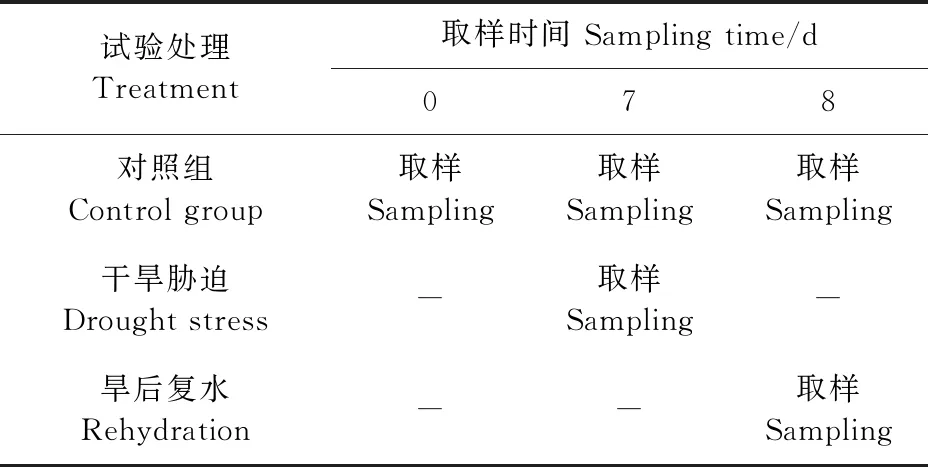
表1 試驗設計與取樣時間
1.3 碳素代謝相關指標
淀粉與可溶性糖的測定采用蒽酮比色法[12];蔗糖合成酶和蔗糖磷酸合成酶活性酶液的提取參考王文靜[13]的方法,測定過程參照於新建[14]和Wardlaw等[15]的方法。
1.4 氮素代謝相關指標
游離氨基酸測定采用茚三酮顯色法;可溶性蛋白測定采用考馬斯亮藍法;全氮測定采用凱氏定氮法,參照《土壤農化分析》[16]測定;硝酸還原酶采用磺胺比色法,參照高俊鳳[12]測定方法。
谷氨酰胺合成酶測定:稱取植物材料0.1 g鮮樣于研缽中,加0.3 mL提取緩沖液 (0.05 mol·L-1Tris-HCl, pH8.0,2 mmol·L-1DTT,2 mmol·L-1Mg2+,0.4 mol·L-1蔗糖),置冰浴上研磨勻漿,轉移于離心管中,4℃下15 000 g離心20 min,上清液即為粗酶液。吸取0.125 mL反應混合液B(反應液A的成分再加入80 mmol·L-1鹽酸羥胺,pH 7.4),加入0.05 mL粗酶液和0.05 mL ATP溶液,混勻,37℃下水浴保溫30 min,加入0.07 mL顯色劑(0.2 mol·L-1TCA,0.37 mol·L-1FeCl3,0.6 mol·L-1HCl),搖勻并放置5 min后,于5 000 g離心10 min,取上清液測定540 nm處的吸光值,以加入0.125 ml反應混合液A(0.1 mol·L-1Tris-HCl緩沖液,pH7.4,80 mmol·L-1Mg2+,20 mmol·L-1谷氨酸鈉鹽,20 mmol·L-1半胱氨酸,2 mmol·L-1EDTA)的為對照。
1.5 數據處理
所有指標測定均設3次生物學重復,計算平均值和標準誤差。數據處理采用Excel 2013 軟件整理,SPSS 16.0進行數據統計分析,應用Duncan多重對比法分析數據,P<0.05的統計檢驗是差異顯著的,運用SigmaPlot 12.0軟件制圖。
2 結果與分析
2.1 干旱脅迫與旱后復水對馬鈴薯塊莖膨大期碳水化合物積累與分配的影響
2.1.1 干旱脅迫與復水對馬鈴薯葉片中碳水化合物積累與分配的影響 由圖1A可知,干旱脅迫后馬鈴薯葉片中可溶性糖含量顯著高于對照,復水后葉片可溶性糖含量仍保持較高水平,未恢復至正常水平。蔗糖是主要的可溶性糖,水分脅迫對源葉中蔗糖含量的影響非常顯著。圖1B表明,無論是干旱脅迫還是解除干旱脅迫,蔗糖含量均顯著高于對照水平,但復水后蔗糖含量較干旱條件下有所下降。蔗糖磷酸合成酶(SPS)存在于植物細胞質,主要參與蔗糖合成。由圖1C可知,干旱脅迫與復水條件下,馬鈴薯葉片中SPS活性均顯著高于對照,復水后SPS活性仍高于對照,這與蔗糖的變化相一致。
蔗糖合成酶(SS)既可催化蔗糖合成又可催化蔗糖分解,是一種可逆酶,但通常認為SS主要起分解蔗糖的作用[18]。由圖1D可知,干旱脅迫后SS活性較對照顯著升高,復水后與對照差異不顯著。葉片淀粉含量在干旱脅迫下顯著升高(圖1E),在干旱脅迫解除后,淀粉含量卻顯著降低。
2.1.2 干旱脅迫與復水對馬鈴薯塊莖中碳水化合物積累與分配的影響 塊莖中可溶性糖含量不僅反映源端光合同化物的供應能力,也可反映庫端對光合同化物的轉化利用能力。圖2A表明,馬鈴薯受到干旱脅迫時,塊莖中可溶性糖含量高于對照水平,恢復正常土壤水分狀況后,馬鈴薯塊莖中可溶性糖含量仍保持較高水平。

注:0 d:處理前;D7:干旱脅迫7 d;DW1:干旱脅迫7 d后復水1 d;不同字母表示不同處理間差異顯著(P<0.05)。下同。 Note: 0d: before the treatment; D7: drought stress for 7 days; DW1: rehydration for 1 day after drought stress of 7 days; Different letters indicate significant difference between different treatments (P<0.05). The same below.圖1 干旱脅迫及復水對馬鈴薯葉片中蔗糖磷酸合成酶(SPS)和蔗糖合成酶(SS)活性、可溶性糖、蔗糖及淀粉含量的影響Fig.1 Effects of drought stress and rehydration on sucrose phosphate synthase (SPS) activity, sucrose synthase (SS) activity, soluble sugar, sucrose, and starch content of leaves in potato
干旱脅迫下塊莖中SS與SPS活性均顯著增強(圖2B和圖2C),但SS活性高于SPS,蔗糖的降解速度大于合成速度,因而SS活性增強加快了蔗糖的降解,蔗糖含量顯著降低(圖2D)。淀粉含量是碳水化合物積累的重要指標之一,干旱脅迫后淀粉含量呈顯著下降趨勢(圖2E)。復水后,SS與SPS活性迅速下降,但蔗糖含量顯著增多,淀粉含量也相應增加。
2.2 干旱脅迫與旱后復水對馬鈴薯塊莖膨大期氮素同化物積累與分配的影響
2.2.1 干旱脅迫與復水對馬鈴薯葉片中氮素同化物積累與分配的影響 圖3A表明,干旱脅迫后馬鈴薯葉片硝酸還原酶(NR)活性顯著降低,復水后NR活性與對照差異不顯著。圖3B顯示,干旱脅迫下葉片谷氨酰胺合成酶(GS)活性與對照差異不顯著,復水后GS活性顯著高于對照水平。圖3C和圖3D顯示,干旱脅迫條件下,葉片中游離氨基酸與可溶性蛋白含量均顯著低于對照水平,全氮含量也呈相同趨勢(圖3E),復水后游離氨基酸、可溶性蛋白以及全氮含量均仍保持較低水平,顯著低于對照。
2.2.2 干旱脅迫與復水對馬鈴薯塊莖中氮素同化物積累與分配的影響 圖4A、圖4B表明,塊莖中游離氨基酸與可溶性蛋白含量在干旱脅迫下均顯著高于對照;由圖4C可知,干旱脅迫下塊莖中全氮含量也顯著高于對照,復水后全氮含量與對照差異不顯著,而可溶性蛋白質與游離氨基酸含量仍顯著高于對照。表明干旱脅迫促進馬鈴薯塊莖中氮素同化物的合成與積累,復水后對氮素同化物的合成與分配影響較小。

圖2 干旱及復水對馬鈴薯塊莖中蔗糖磷酸合成酶(SPS)和蔗糖合成酶(SS)活性、可溶性糖、蔗糖及淀粉含量的影響Fig.2 Effects of drought stress and rehydration on sucrose phosphate synthase (SPS) activity, sucrose synthase (SS) activity, soluble sugar, sucrose, and starch content of tuber in potato tuber
2.3 干旱脅迫與復水對馬鈴薯葉片和塊莖中碳氮平衡的影響
以可溶性糖及淀粉含量之和與全氮含量的比值表示碳氮比。由圖5A中可知,干旱脅迫下馬鈴薯葉片中碳氮比增加,復水后仍保持較高水平;塊莖中碳氮比在干旱脅迫后顯著下降,復水后可迅速恢復正常水平,并顯著高于對照(圖5B)。表明馬鈴薯在干旱脅迫與復水過程中影響了碳水化合物與氮素同化物的合成與分配,使碳氮比發生改變。
3 討 論
3.1 干旱脅迫及復水對馬鈴薯碳水化合物轉運的影響
植物生長發育與碳水化合物的轉運緊密相關,光合作用產生的碳水化合物為植物新陳代謝提供能量來源,同時碳水化合物參與植物源組織和庫組織間的調控與分配[17-18]。研究認為非結構性碳水化合物是植物碳水化合物在生產過剩時積累的暫時儲存物質[19],包含蔗糖、果糖及淀粉等[20]。而干旱脅迫將干擾碳水化合物的合成、積累與轉運過程。
可溶性糖在一定程度上可反映植物受逆境脅迫的程度。本研究發現,水分脅迫下馬鈴薯葉片中可溶性糖含量顯著增加,同時塊莖中可溶性糖也大量積累,表明此時馬鈴薯受干旱脅迫的程度較為嚴重,而且干旱脅迫促進了可溶性糖從葉片向塊莖中轉運,參與細胞滲透調節過程,防止細胞大量脫水。另外,干旱脅迫后馬鈴薯葉片SPS活性高于SS,蔗糖合成速率大于降解速率,蔗糖大量積累,但塊莖中蔗糖含量顯著降低。表明干旱脅迫后,馬鈴薯葉片中合成的蔗糖主要積累于葉片中,同時,干旱脅迫下馬鈴薯葉片中淀粉發生積累,塊莖中淀粉的合
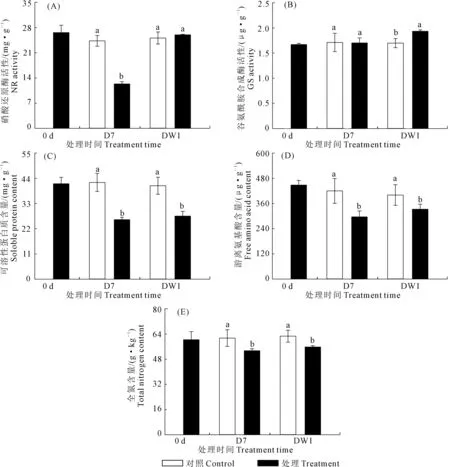
圖3 干旱脅迫及復水對馬鈴薯葉片中硝酸還原酶(NR)和谷氨酰胺合成酶(GS)活性、可溶性蛋白質、 游離氨基酸以及全氮含量的影響Fig.3 Effects of drought stress and rehydration on nitrate reductase and glutamine synthesis (GS) activity, soluble protein, free amino acid and total nitrogen content of leaves in potato tuber swelling stage

圖4 干旱脅迫及復水對馬鈴薯塊莖中可溶性蛋白、游離氨基酸以及全氮含量的影響Fig.4 Effects of drought stress and rehydration on soluble protein content, free amino acid content, and total nitrogen content in tuber of potato tuber

圖5 干旱脅迫及復水對馬鈴薯葉片與塊莖中碳氮比的影響Fig.5 Effects of drought stress and rehydration on the ratio of C/N of leaves and tuber of potato
成卻受到制約,說明干旱脅迫顯著阻礙了馬鈴薯葉片中碳水化合物向塊莖轉運,影響了碳水化合物轉運過程相關酶的活性,這與Zrenner R等[21]對菠菜的研究結果一致。復水后,葉片與塊莖中可溶性糖含量仍保持較高水平,這可能與復水時間較短有關,可溶性糖的合成與轉運尚未恢復正常水平。而葉片SPS活性顯著增強,促進了葉片中蔗糖的合成與轉運,此時塊莖中蔗糖含量也迅速增加,充分說明旱后復水恢復了碳水化合物從葉片向塊莖的轉運。在蔗糖充分供應的前提下,塊莖中SS活性顯著增強,淀粉的合成得以恢復,淀粉快速積累。由此表明,干旱脅迫后復水能夠在一定程度上恢復馬鈴薯碳水化合物從葉片向塊莖的轉運,修復干旱脅迫對馬鈴薯植株造成的損傷,保障了馬鈴薯植株碳水化合物轉運過程正常運行。
3.2 干旱脅迫及復水對馬鈴薯氮素同化物轉運的影響
氮素在植物生命活動中發揮重要作用[22]。植物主要通過根系吸收土壤氮素中的無機氮。而葉片是植物氮同化的主要部位,植物同化氮素主要是經由硝酸還原酶(NR)與谷氨酰胺合成酶(GS)等關鍵酶進行。隨后將同化后的物質轉運到植株的各個部位,參與生長代謝過程[23-25]。植物處于逆境脅迫時氮素轉運同樣會發生較大的改變[26]。
高等植物從土壤吸收的氮在葉肉細胞中同化后,用于葉片蛋白質的合成或以氨基酸的形式通過韌皮部輸送到庫組織[31]。研究發現,土壤干旱使葉片內氨基酸合成受阻,進而影響葉片中蛋白質的合成。此外,干旱脅迫下蛋白質水解是植株適應環境的一個重要的代謝變化,釋放的氨基酸向塊莖轉運,合成新的蛋白質,同時某些特定的蛋白也可能被激活[32-33],因而塊莖中氮素同化物含量均顯著增加。說明干旱脅迫促進馬鈴薯葉片氮素向塊莖重新分配,以適應干旱脅迫環境。
3.3 干旱脅迫及復水對馬鈴薯碳氮平衡的影響
全球環境的改變導致陸地生態系統的結構和功能發生變化,而植物的碳氮營養及其分配對環境變化的調節作用是其最為重要的過程之一[34-35]。其中碳氮平衡對作物生長代謝的調控發揮重要作用[36]。而碳氮比可反映植株碳氮轉運協調程度[37]。但水分虧缺會顯著影響植株體內碳氮轉運過程,進而影響到氮素同化物和碳水化合物的組分平衡。
Chen等[38]對高粱進行干旱脅迫處理后,發現高粱葉片中碳氮比增加。本研究經干旱脅迫處理后,馬鈴薯植株碳水化合物與氮素同化物的合成與轉運均受到顯著影響,表現為可溶性糖在葉片中大量合成,而氮素同化物的積累減少,塊莖中淀粉的合成受阻,全氮含量卻增加,導致馬鈴薯葉片與塊莖碳氮失衡,葉片碳氮比增加,這與上述研究結果一致。葉片碳氮比增加的原因:一方面在于干旱脅迫下碳水化合物由于轉運不暢而大量積累,而且碳素轉運過程需要氮素轉運途徑提供相應的氨基酸,用以合成光合作用有關的蛋白質;另一方面在于干旱脅迫降低了葉片中氮素的積累與同化,導致氮素同化物積累不足。同時,馬鈴薯塊莖中碳氮比下降原因在于:塊莖中氮素的同化增強,需要碳水化合物轉運過程提供能量,導致能量消耗過大,同時消耗大量光合產物,使塊莖中碳水化合物供應不足,碳氮失衡,可能是淀粉貯存含量下降的原因之一。
復水后馬鈴薯葉片中碳氮比仍保持較高水平,但復水對馬鈴薯塊莖碳氮比影響非常顯著。這是因為復水恢復了碳水化合物的合成與轉運,塊莖中的淀粉大量積累,進而使干旱脅迫導致的低水平的碳氮比迅速升高。另外,由上述可知,復水恢復了氮素的吸收與同化能力,且全氮含量已恢復正常水平,表明旱后復水在某種程度上恢復了馬鈴薯碳水化合物與氮素同化物的合成、分配及轉運過程,修復了干旱脅迫對植物造成的損傷。但是,可能由于復水處理時間較短,氮素同化物在復水后的變化并不顯著,所以,后期試驗可以適當延長復水后取樣時間,以期對碳氮轉運在復水后修復過程中的作用有更加深入地了解。
4 結 論
研究發現,干旱脅迫下馬鈴薯葉片蔗糖與淀粉進行積累,塊莖中淀粉含量降低,淀粉合成受到制約,復水后,葉片SPS活性顯著增強,葉片蔗糖的合成與轉運過程得以恢復,促進塊莖蔗糖含量增加,在蔗糖充分供應的前提下,塊莖淀粉迅速積累。由此表明,干旱脅迫顯著阻礙了馬鈴薯葉片中碳水化合物向塊莖轉運,復水后能夠在一定程度上恢復馬鈴薯碳水化合物從葉片向塊莖的轉運,修復干旱脅迫對馬鈴薯植株造成的損傷,保障了馬鈴薯植株碳水化合物轉運過程正常運行。
另外,干旱脅迫下馬鈴薯葉片NR活性降低,使葉片中氮素還原過程受阻,葉片游離氨基酸、可溶性蛋白質與全氮含量均顯著降低,而塊莖中游離氨基酸與可溶性蛋白質含量增加,說明干旱脅迫促進馬鈴薯葉片氮素向塊莖重新分配。復水后葉片中NR與GS活性顯著增強,而葉片游離氨基酸、可溶性蛋白質以及全氮含量仍保持較低水平,塊莖游離氨基酸、可溶性蛋白含量較高。表明旱后復水雖然恢復了氮素的吸收與同化能力,但氮素同化物的合成與轉運尚未恢復正常水平,這可能是復水時間較短所致。
由此表明,干旱脅迫下馬鈴薯葉片可溶性糖積累,氮素同化物減少積累,而塊莖中呈相反現象,因而馬鈴薯葉片與塊莖中碳氮失衡,葉片碳氮比增加,塊莖碳氮比降低。復水后,上述情況均得到一定程度的緩解,表現為塊莖碳氮比增加。因此,旱后復水可在一定程度上修復干旱脅迫造成的損傷。
