基于SCoT分子標記的48份楊桃種質(zhì)遺傳多樣性分析
2019-09-10 07:22:44歐景莉朱楊帆陳豪軍周俊岸陳燕何江寧琳潘祖建甘衛(wèi)堂
南方農(nóng)業(yè)學(xué)報 2019年8期
歐景莉 朱楊帆 陳豪軍 周俊岸 陳燕 何江 寧琳 潘祖建 甘衛(wèi)堂
摘要:【目的】分析48份楊桃種質(zhì)的遺傳多樣性,為楊桃種質(zhì)資源的鑒定保護及開發(fā)利用提供理論依據(jù)。【方法】從60條SCoT引物中篩選出擴增條帶清晰穩(wěn)定、多態(tài)性豐富的引物,對從國內(nèi)外收集的48份楊桃種質(zhì)材料進行PCR擴增,統(tǒng)計分析電泳圖譜,利用Popgene 1.32計算其遺傳多樣性參數(shù),采用NTSYSpc 2.1計算種質(zhì)間的遺傳相似系數(shù)和遺傳距離,利用UPGMA法進行聚類分析,并根據(jù)遺傳相似系數(shù)進行主成分分析。【結(jié)果】從60條SCoT引物中篩選出的11條引物共擴增出115條條帶,其中多態(tài)性條帶102條,占總條帶數(shù)的88.7%,每條SCoT引物擴增的總條帶數(shù)(TNB)和多態(tài)性條帶數(shù)(NPB)分別為10.45和9.27條,多態(tài)性比率(PPB)為70.00%~100.00%,平均為88.84%;多態(tài)性信息量(PIC)、有效等位基因數(shù)(Ne)、Nei’s基因多樣性指數(shù)(H)和Shannon’s信息指數(shù)(I)的平均值分別為0.77、1.7758、0.4229和0.6061。48份楊桃種質(zhì)間的遺傳相似系數(shù)為0.4957~0.9217,平均為0.6841。聚類分析結(jié)果顯示,在遺傳相似系數(shù)0.6618處可將48份楊桃種質(zhì)劃分為三大類群,第Ⅰ類群均為甜楊桃種質(zhì),第Ⅱ類群以酸楊桃種質(zhì)為主,第Ⅲ類群以甜楊桃種質(zhì)為主。主成分分析結(jié)果與聚類分析結(jié)果基本一致,均與楊桃的果實風(fēng)味和種質(zhì)來源高度相關(guān)。【結(jié)論】楊桃種質(zhì)資源具有較豐富的遺傳多樣性,且篩選獲得的SCoT引物對楊桃種質(zhì)資源有較高的多態(tài)性檢測率,適用于楊桃種質(zhì)資源鑒別及親緣關(guān)系分析。
關(guān)鍵詞: 楊桃;遺傳多樣性;SCoT標記;聚類分析;主成分分析
中圖分類號: S667.902.4? ? ? ? ? ? ? ? ? ? ? ? ? ? ?文獻標志碼: A 文章編號:2095-1191(2019)08-1680-08
Genetic relationship analysis of 48 Averrhoa carambola L. germplasms based on SCoT marker
OU Jing-li, ZHU Yang-fan, CHEN Hao-jun*, ZHOU Jun-an, CHEN Yan, HE Jiang,NING Lin, PAN Zu-jian, GAN Wei-tang
(Guangxi Subtropical Crops Research Institute, Guangxi Academy of Agricultural Sciences, Nanning? 530001, China)
Abstract:【Objective】In order to provide theoretical support and reference for the identification,protection,development and utilization of Averrhoa carambola L. germplasm resources,the present study analyzed the genetic diversity of 48 A. carambola germplasms. 【Method】SCoT primers with rich polymorphism and clear amplified bands were screened out from 60 primers,the 48 A. carambola germplasm resources collected from home and abroad were amplified by PCR. The electrophoretogram was analyzed, and genetic diversity parameters were calculated by Popgene 1.32. Then the genetic similarity coefficient and genetic distance between different germplasms were calculated using NTSYSpc 2.1. The unweighted pair-group method with arithmetic mean(UPGMA) method was used for cluster analysis. Based on the genetic similarity,principal coordinate analysis was also carried out. 【Result】A total of 115 bands were amplified from 11 primers which screened out from 60 SCoT primers,including 102 polymorphic bands, accounting for 88.7% of the total. The total number of bands(TNB) and polymorphic bands(NPB) amplified from each primer were 10.45 and 9.27,respectively. The percentage of polymorphism bands(PPB) was 70.00%-100.00%,with an average of 88.84%.? The mean value of polymorphism information content(PIC), effective number of alleles(Ne),Nei’s gene diversity(H) and Shannon’s information index(I) were 0.77, 1.7758,0.4229 and 0.6061,respectively. The genetic similarity coefficient between different A. ca-rambola germplasms was 0.4957-0.9217,and the average was 0.6841. According to the results of cluster analysis,48 A. carambola germplasms could be divided into three groups at the genetic similarity of 0.6618. All the A. carambola germplasms in groupⅠand Ⅲ belonged to sweet A. carambola,while groupⅡwas mainly acid A. carambola. The results of principal component analysis and cluster analysis were basically consistent, and both were highly related to the fruit flavor and germplasm sources. 【Conclusion】The genetic diversity of A. carambola germplasm resources is quite rich. SCoT marker has high polymorphism detection efficiency on A. carambola germplasm. It can be used for the identification and relationship analysis of A. carambola germplasms.
Key words: Averrhoa carambola L.; genetic diversity; SCoT marker; clustering analysis; principal component analysis
0 引言
【研究意義】楊桃(Averrhoa carambola L.)為熱帶亞熱帶水果,在我國已有1500多年的栽培歷史,廣西、廣東、福建、海南、臺灣和云南等省(區(qū))均有較大面積栽培,其可加工成罐頭、蜜餞、果酒、果醬和蜜煉果膏等,滿足了人們對果品多樣化、營養(yǎng)化及保健化的需求(劉韜等,2011;劉鍇棟等,2013)。楊桃種質(zhì)資源較豐富,其栽培品種多由實生變異而來,故品種(系)間的形態(tài)特征極其相似,易出現(xiàn)同物異名或同名異物的現(xiàn)象,難以科學(xué)準確鑒別。此外,由于歷史原因,一些地方品種的來源和親緣關(guān)系難以確認,給楊桃種質(zhì)資源的科學(xué)保護及有效利用帶來了極大困難。因此,開展楊桃種質(zhì)資源的遺傳多樣性及親緣關(guān)系分析,對楊桃的種質(zhì)鑒定及有效開發(fā)利用具有重要意義。【前人研究進展】目標起始密碼子多態(tài)性分子標記(SCoT)是一種基于翻譯起始位點的目的基因標記(Collard and Mackill,2009),其操作簡單、多態(tài)性高、重復(fù)性好、引物通用性強,能有效產(chǎn)生與性狀連鎖的位點,可檢測出不同基因型供試材料間的微小差異,準確反映種質(zhì)間親緣關(guān)系(蔡元保等,2014)。目前,SCoT分子標記已廣泛應(yīng)用于龍眼(陳虎等,2010,2012)、芒果(Luo et al.,2010;羅聰,2012)、枇杷(韓國輝等,2011a;龍治堅,2013)、柑橘(韓國輝等,2011b;靳佳佳,2018)、葡萄(Guo et al.,2012;王發(fā)明等,2017)、番木瓜(楊祥燕等,2013)、澳洲堅果(蔡元保等,2013)、桃(陳紅等,2014)、梨(和世玉,2014;楊君祎,2015)、番石榴(何江,2017)和柿子(關(guān)長飛等,2019)等果樹的種質(zhì)鑒定、基因差異表達、遺傳多樣性和親緣關(guān)系分析及指紋圖譜構(gòu)建、功能基因挖掘等。隨著近年來分子標記的廣泛應(yīng)用,已有多位學(xué)者利用RAPD分子標記對楊桃種質(zhì)的遺傳多樣性進行研究。戴子云(2010)研究發(fā)現(xiàn),17份廣東楊桃種質(zhì)的地理起源與RAPD表型無相關(guān)性;劉韜等(2011)對17份福建楊桃品種進行遺傳多樣性分析,結(jié)果發(fā)現(xiàn)其具有豐富的遺傳多樣性;劉鍇棟等(2013)研究發(fā)現(xiàn),RAPD分子標記與形態(tài)標記所反映出的10份廣東地區(qū)楊桃品種間的遺傳多樣性具有較高一致性和可信度,因此,將二者結(jié)合起來可較好地揭示楊桃種質(zhì)的遺傳多樣性。【本研究切入點】雖然前人已采用RAPD分子標記對我國部分地區(qū)楊桃種質(zhì)的遺傳多樣性進行分析,但供試材料較少,種質(zhì)來源不夠廣泛,無法為楊桃種質(zhì)資源的有效利用提供系統(tǒng)全面的參考。且目前尚無利用SCoT分子標記對楊桃種質(zhì)資源進行遺傳多樣性分析的研究報道。【擬解決的關(guān)鍵問題】采用SCoT分子標記對從國內(nèi)外收集的48份楊桃種質(zhì)資源進行遺傳多樣性分析,為楊桃種質(zhì)資源的鑒定保護及開發(fā)利用提供理論參考。
1 材料與方法
1. 1 試驗材料
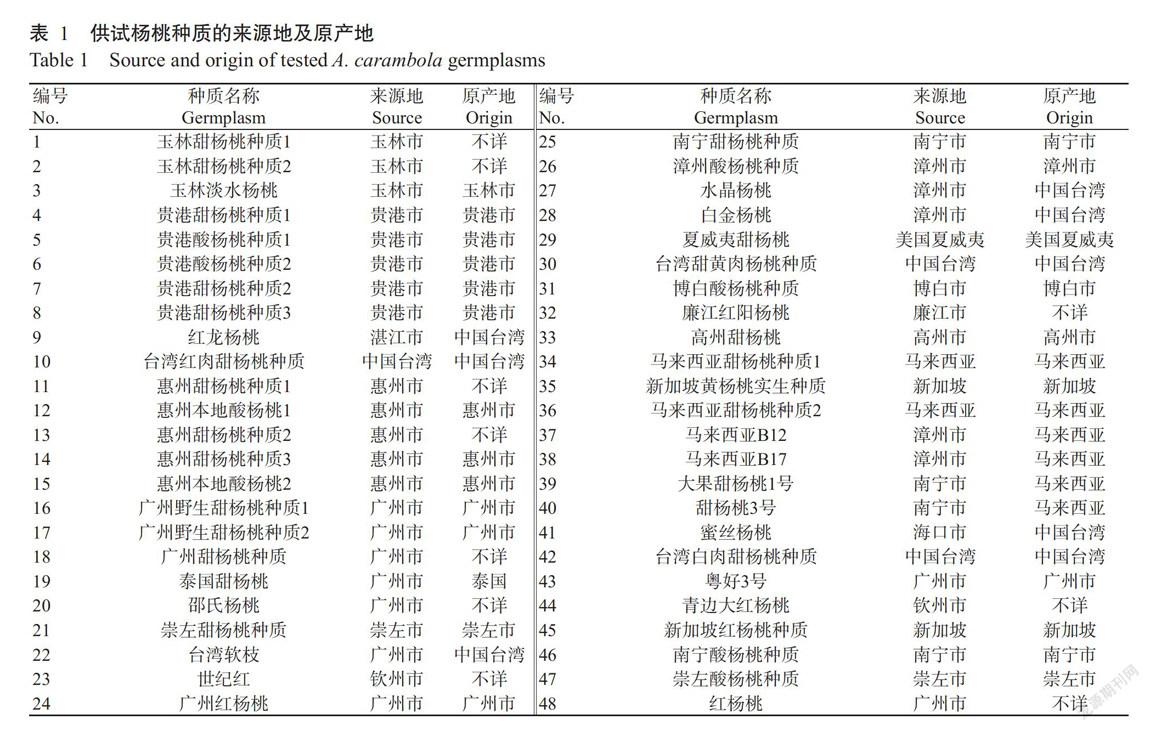
供試的48份楊桃種質(zhì)材料均采自廣西亞熱帶作物研究所楊桃種質(zhì)資源圃,其來源地和原產(chǎn)地見表1。dNTPs、Taq DNA聚合酶和DNA Marker購自天根生化科技(北京)有限公司,其他化學(xué)試劑均購自南寧市茵興科技產(chǎn)品有限公司。主要儀器設(shè)備:PCR擴增儀(ABI Veriti,美國)、冷凍離心機(SIGMA 3-30K,德國)、核酸蛋白測定儀(Bio-Rad Smart Spec Plus,美國)和凝膠成像系統(tǒng)(Bio-Rad UV-3,美國)等。
1. 2 試驗方法
1. 2. 1 DNA提取 選取供試種質(zhì)的健康無病害干凈嫩葉,用液氮速凍后于-80 ℃保存?zhèn)溆谩2捎酶牧嫉腃TAB法(陳虎等,2009)提取嫩葉DNA,并用核酸蛋白測定儀檢測其濃度和純度,最后統(tǒng)一稀釋至30 mg/L,-20 ℃保存?zhèn)溆谩?/p>
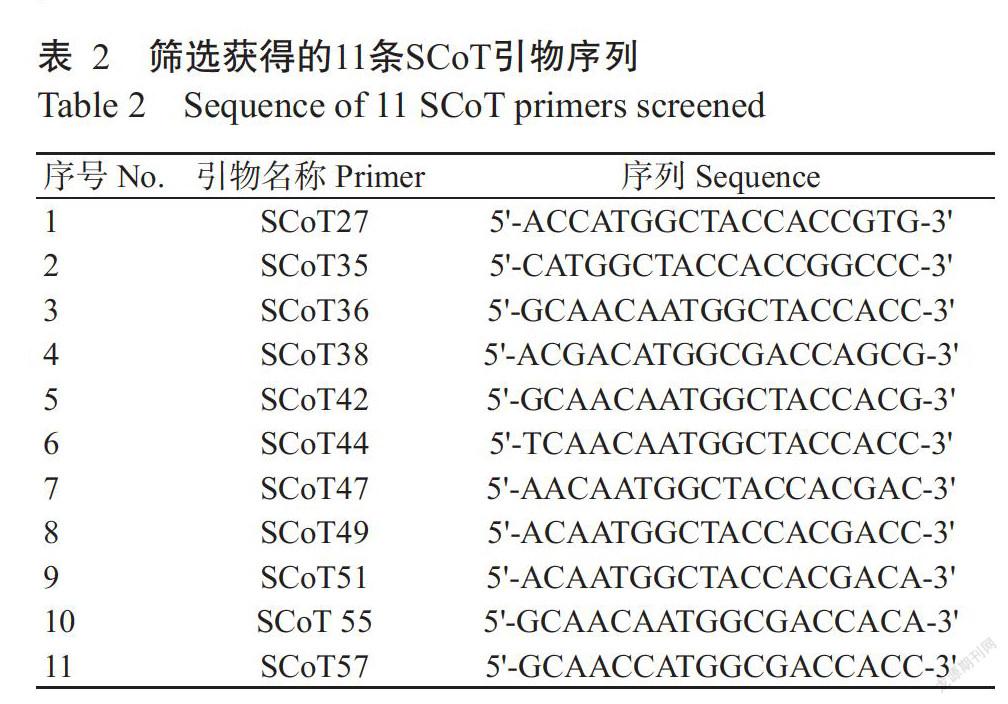
1. 2. 2 SCoT引物篩選及PCR擴增 參考Collard和Mackill(2009)的設(shè)計原則設(shè)計60條SCoT引物(SCoT1~SCoT60),由生工生物工程(上海)股份有限公司合成。采用歐景莉等(2015)優(yōu)化的SCoT-PCR反應(yīng)體系,以馬來西亞B17、廉江紅陽楊桃、貴港酸楊桃種質(zhì)1和夏威夷甜楊桃等8份遺傳背景差異明顯的楊桃種質(zhì)嫩葉DNA為模版,從60條SCoT引物中篩選出11條擴增條帶清晰穩(wěn)定、多態(tài)性好的引物(表2),對48份供試楊桃種質(zhì)進行PCR擴增。SCoT-PCR反應(yīng)體系20.0 μL:2.5 mmol/L Mg2+,0.3 mmol/L dNTPs,30 mg/L DNA模板,1.00 μmol/L引物,0.4 U Taq DNA聚合酶。擴增程序:94 ℃預(yù)變性5 min;94 ℃ 1 min;50 ℃ 1 min;72 ℃ 90 s,進行35個循環(huán);72 ℃延伸10 min。PCR產(chǎn)物用1.8%瓊脂糖凝膠電泳進行檢測,并在Bio-Rad UV-3型凝膠成像系統(tǒng)上觀察并拍照。
1. 3 統(tǒng)計分析
觀察電泳圖譜,按同一位置有無條帶進行賦值,有條帶記為“1”,無條帶記為“0”,最終形成(0,1)矩陣。利用Popgene 1.32計算遺傳多樣性參數(shù),包括各引物的擴增總條帶數(shù)(Total number of bands,TNB)、多態(tài)性條帶數(shù)(Number of polymorphism bands,NPB)、多態(tài)性條帶百分比(Percentage of polymorphism bands,PPB)、有效等位基因數(shù)(Effective number of alleles,Ne)、多態(tài)性信息量(Polymorphism information content,PIC)、Nei’s基因多樣性指數(shù)(Nei’s gene diversity,H)和Shannon’s信息指數(shù)(Shannon’s information index,I)。采用NTSYSpc 2.1計算不同種質(zhì)間的遺傳相似系數(shù)和遺傳距離,利用UPGMA法進行聚類分析,并根據(jù)遺傳相似系數(shù)進行主成分分析(PCA)。
2 結(jié)果與分析
2. 1 遺傳多樣性分析結(jié)果
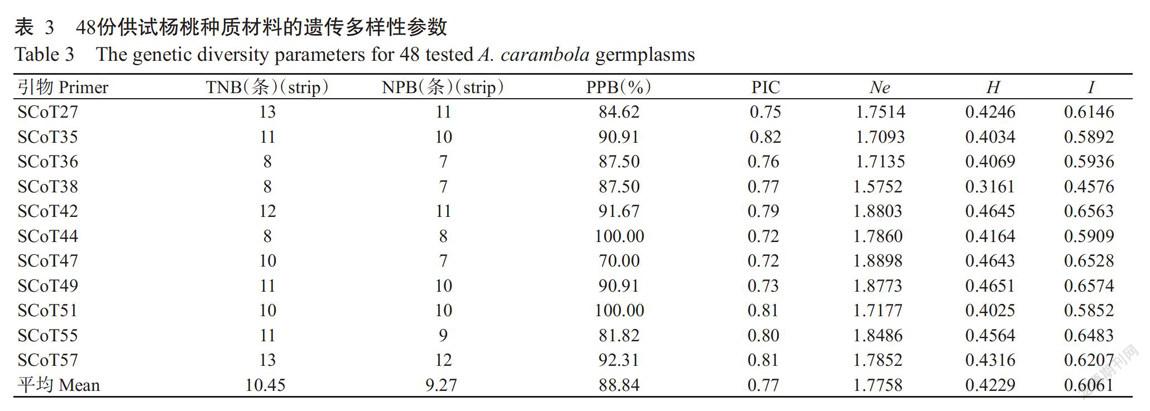
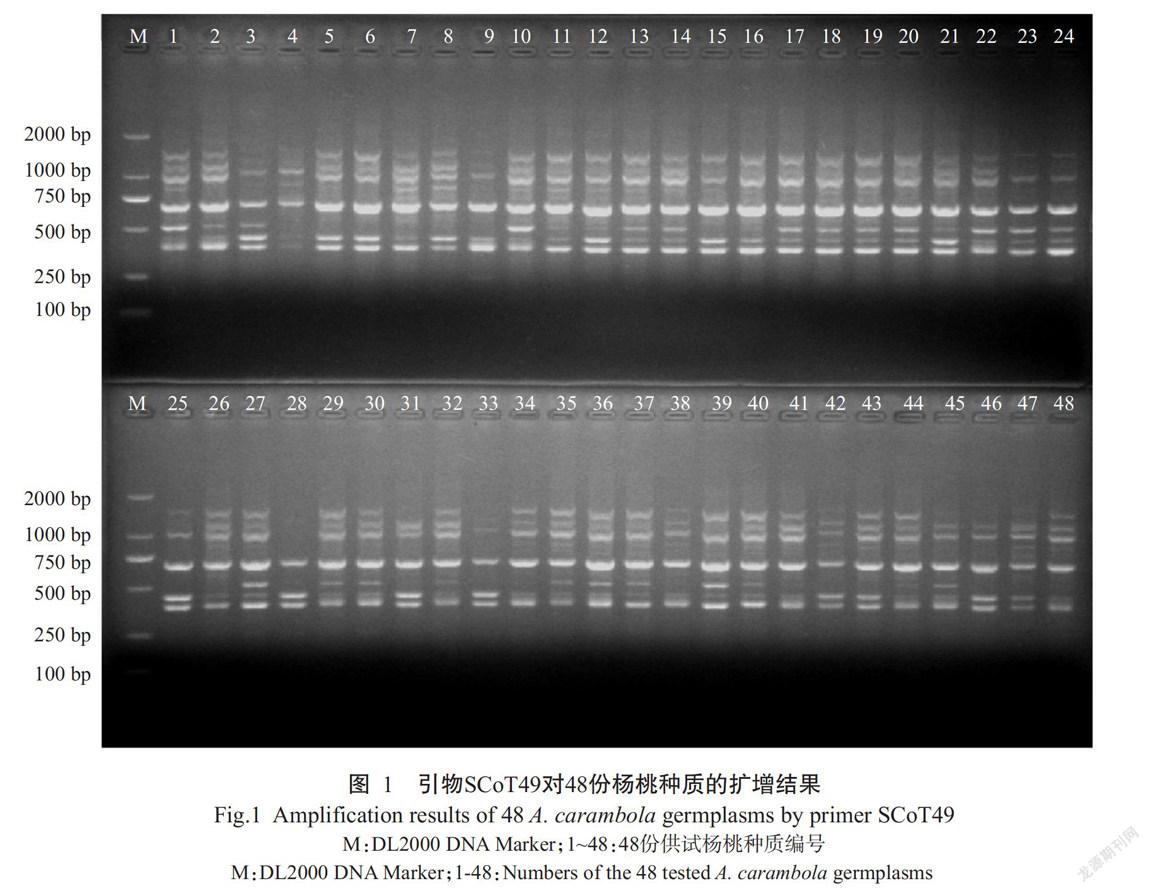
從60條SCoT引物中篩選出11條擴增條帶清晰、多態(tài)性好的引物,利用其對48份楊桃種質(zhì)資源進行擴增,共得到115條條帶,其中多態(tài)性條帶數(shù)為102條,占總條帶數(shù)的88.7%,多態(tài)性較好(表3)。其中,引物SCoT49的擴增結(jié)果如圖1所示。每條引物的TNB為8~13條,NPB為7~12條;PPB為70.00%~100.00%,平均為88.84%,其中,引物SCoT44和SCoT51的PPB均為100.00%;PIC為0.72~0.82,平均為0.77;Ne為1.5752~1.8898,平均為1.7758;H為0.3161~0.4651,平均為0.4229;I為0.4576~0.6574,平均為0.6061,表明楊桃種質(zhì)資源具有較豐富的遺傳多樣性,且篩選獲得的11條SCoT引物對楊桃種質(zhì)資源有較高的多態(tài)性檢測率,適用于楊桃種質(zhì)資源的鑒別及親緣關(guān)系分析。
2. 2 聚類分析結(jié)果
利用NTSYSpc 2.1計算48份楊桃種質(zhì)材料間的遺傳相似系數(shù),再利用UPGM法進行聚類分析,結(jié)果如圖2所示。48份楊桃種質(zhì)的遺傳相似系數(shù)為0.4957~0.9217,平均為0.6841,其中,廣州野生甜楊桃種質(zhì)2(17)(括號內(nèi)為種質(zhì)編號,下同)和廣州甜楊桃種質(zhì)(18)間的遺傳相似系數(shù)最高,為0.9217,結(jié)合二者的采集地、樣本樹齡及農(nóng)藝性狀、植物學(xué)性狀等數(shù)據(jù)信息,廣州野生甜楊桃種質(zhì)2很有可能是廣州甜楊桃種質(zhì)的實生后代;玉林甜楊桃種質(zhì)1(1)與紅龍楊桃(9)間的遺傳相似系數(shù)最小,為0.4957,說明二者的遺傳差異最大,推測二者的原產(chǎn)地相隔較遠,在進化過程中產(chǎn)生明顯的基因差異。
在遺傳相似系數(shù)為0.6618處,可將48份楊桃種質(zhì)材料劃分為三大類群。第Ⅰ類群包含27份甜楊桃種質(zhì),該類群在遺傳相似系數(shù)0.7020處又可分為A和B兩個亞群,其中,A亞群共14份種質(zhì),除馬來西亞B17(38)外,原產(chǎn)地為馬來西亞的種質(zhì)(34、36、37、39和40)均聚在此亞群,A亞群還包含了2份玉林甜楊桃種質(zhì)(1、2)、2份臺灣種質(zhì)(10、41)及來源于惠州(11)、廉江(32)、廣州(48)、欽州(44)和新加坡(35)的種質(zhì)各1份;B亞群包含13份種質(zhì),除了來源于崇左的甜楊桃種質(zhì)(21)、漳州的水晶楊桃(27)、欽州的世紀紅(23)外,其余10份均來源于廣東的廣州和惠州。雖然紅楊桃(48)和廣州紅楊桃(24)均采源于廣州,且名稱極為相似,但分別屬于不同亞群,說明二者非同一種質(zhì)。第Ⅱ類群包含14份楊桃種質(zhì),其中包括8份酸楊桃種質(zhì),還包括3份來源于貴港的甜楊桃種質(zhì)(4、7、8)及來源于臺灣(42)、廣州(43)和南寧(25)的甜楊桃種質(zhì)各1份。可見,該類群種質(zhì)的果實風(fēng)味多為酸、酸甜或甜酸。第Ⅲ類群包含7份楊桃種質(zhì),其中,3份原產(chǎn)地為臺灣(9、28和30),其他4份原產(chǎn)地分別為玉林(3)、高州(33)、馬來西亞(38)和新加坡(45)。該類群中,除玉林淡水楊桃(3)的果實甜味和酸味均很淡,介于甜楊桃和酸楊桃之間外,其他種質(zhì)均為甜楊桃。玉林淡水楊桃(3)與類群內(nèi)其他種質(zhì)的遺傳相似系數(shù)均較小,從遺傳角度上反映出該類群種質(zhì)在風(fēng)味特征上的差異。
2. 3 主成分分析結(jié)果
根據(jù)遺傳相似系數(shù)進行主成分分析,并繪制二維散點平面分布圖,如圖3所示。前3個主成分貢獻率依次為13.02%、9.11%和7.45%,累計貢獻率為29.58%,能代表原始數(shù)據(jù)的主要信息。在二維散點平面分布圖中,種質(zhì)間的距離越近,表示親緣關(guān)系越近;反之則越遠。根據(jù)種質(zhì)間的距離可將48份楊桃種質(zhì)分成A、B、C和D 4個組,其中,A組和B組分別相當(dāng)于聚類分析結(jié)果中第Ⅰ類群的A亞群和B亞群,C組和D組分別相當(dāng)于第Ⅱ類群和第Ⅲ類群,表明主成分分析結(jié)果與聚類分析結(jié)果高度相似。A組和B組存在交錯現(xiàn)象,與聚類分析結(jié)果中A和B兩個亞群同屬Ⅰ類群相互印證。但在主成分分析結(jié)果中,馬來西亞甜楊桃種質(zhì)1(34)歸為C組,可能其起源與C組的種質(zhì)更接近,因相關(guān)科研資料欠缺,歷史年代久遠,現(xiàn)已無法考證。從整體來看,供試楊桃種質(zhì)在圖3中分布廣泛,表明其遺傳背景較廣,遺傳多樣性較豐富。綜合對比圖2和圖3可知,系統(tǒng)聚類分析結(jié)果和主成分分析結(jié)果基本一致,雖然主成分分析的分類結(jié)果不如聚類分析結(jié)果可將各組邊界明確劃分,但能更直觀地反映出種質(zhì)間的親緣關(guān)系,將兩者結(jié)合可更全面地了解各種質(zhì)間的親緣關(guān)系。
3 討論
SCoT分子標記是一種根據(jù)起始密碼子ATG側(cè)翼序列的保守性和一致性設(shè)計單向引物,擴增產(chǎn)生偏向候選功能基因區(qū)的顯性多態(tài)性標記(Collard and Mackill,2009),且其擴增的多態(tài)性條帶絕大部分是基因組內(nèi)部的功能基因或與之緊密相鄰,能有效與目標性狀連鎖,可通過篩選某個分子標記來實現(xiàn)性狀篩選(熊發(fā)前等,2009)。本研究選用11條SCoT引物對48份楊桃種質(zhì)進行PCR擴增,共得到115條條帶,其中多態(tài)性條帶數(shù)為102條,占總條帶數(shù)的88.7%。戴子云(2010)利用13條PAPD引物從17份廣東楊桃種質(zhì)擴增獲得的多態(tài)條帶百分比為59%;劉韜等(2011)利用13條RAPD引物從17份福建楊桃種質(zhì)擴增獲得的多態(tài)性條帶百分比為97.19%;劉鍇棟等(2013)利用10條RAPD引物從10份廣東楊桃種質(zhì)擴增獲得的多態(tài)性條帶百分比為93.02%。劉韜等(2011)、劉鍇棟等(2013)擴增獲得的多態(tài)性條帶百分比較本研究稍高,究其原因可能是本研究供試材料較多,但僅用11條SCoT引物進行多態(tài)性分析。PIC作為評價物種遺傳多樣性參數(shù)的重要指標,當(dāng)PIC大于0.5時,具有高度多態(tài)性(Vanhala et al.,1998)。本研究篩選出的SCoT引物平均PIC為0.77,遠高于0.5,說明這些引物為高度多態(tài)性信息引物,具有較高的檢測效率,可用于楊桃種質(zhì)鑒定、遺傳多樣性分析及遺傳圖譜構(gòu)建,且這些多態(tài)性位點很可能是功能基因或與目標性狀相關(guān)聯(lián),為基因定位和基因克隆提供理論依據(jù)。
分析了解楊桃種質(zhì)資源間的親緣關(guān)系及變異信息,為楊桃種質(zhì)資源鑒定及育種時親本選擇提供理論依據(jù)。根據(jù)本研究聚類分析和主成分分析結(jié)果,48份楊桃種質(zhì)可分為三大類群,所有酸楊桃種質(zhì)均屬于第Ⅱ類群,而甜楊桃種質(zhì)則主要集中在第Ⅰ和Ⅲ類群,僅少數(shù)甜楊桃種質(zhì)在第Ⅱ類群。第Ⅰ類群又可在遺傳相似系數(shù)0.7020處分為A和B兩個亞群,A亞群以原產(chǎn)地為馬來西亞及來源于玉林和新加坡等地的種質(zhì)為主,B亞群則主要為廣東的種質(zhì)。而來自廣西貴港和南寧的種質(zhì)集中在Ⅱ類群,來源于我國臺灣的種質(zhì)則主要集中在第Ⅲ類群。可見,本研究的聚類分析結(jié)果與楊桃果實風(fēng)味和種質(zhì)來源高度相關(guān),與劉鍇棟等(2013)的研究結(jié)論基本一致。因此,本研究篩選獲得的11條SCoT引物擴增產(chǎn)物中極可能存在與果實風(fēng)味(酸甜)相關(guān)的基因位點。本研究中,臺灣軟枝和泰國大果甜楊桃歸于同一類,而馬來西亞B17歸于另一類群,與劉韜等(2011)的研究結(jié)果相一致。雖然本研究的聚類分析結(jié)果與楊桃種質(zhì)的果實風(fēng)味和品種來源相關(guān)性較高,但并未嚴格按照果實風(fēng)味或來源地進行劃分,可能是因為部分種質(zhì)來源地并非原產(chǎn)地,說明楊桃種質(zhì)遺傳多樣性較豐富,遺傳背景較復(fù)雜,要明確其親緣關(guān)系需要進行更深入的調(diào)查和研究。因此,要鑒定不同的楊桃種質(zhì),區(qū)分同名異種或同種異名的楊桃品種,除了采用分子標記技術(shù)外,還應(yīng)綜合形態(tài)特征、品質(zhì)性狀等進行分析更有效。
4 結(jié)論
楊桃種質(zhì)資源具有較豐富的遺傳多樣性,且篩選獲得的SCoT引物對楊桃種質(zhì)資源有較高的多態(tài)性檢測率,適用于楊桃種質(zhì)資源鑒別及親緣關(guān)系分析。
參考文獻:
蔡元保,楊祥燕,陳豪軍,曾黎明,龐新華,孫光明. 2014. SRAP結(jié)合SCoT標記分析番木瓜種質(zhì)的遺傳多樣性[J]. 植物遺傳資源學(xué)報,15(2):292-298. [Cai Y B,Yang X Y,Chen H J,Zeng L M,Pang X H,Sun G M. 2014. Genetic diversity analysis of papaya resources by SRAP and SCoT combination[J]. Journal of Plant Genetic Resources,15(2):292-298.]
蔡元保,楊祥燕,陳顯國,曾黎明,郭凌飛,林玉虹,崔明勇. 2013. 澳洲堅果SCoT反應(yīng)體系的建立及應(yīng)用[J]. 熱帶亞熱帶植物學(xué)報,21(3):253-258. [Cai Y B,Yang X Y,Chen X G,Zeng L M,Guo L F,Lin Y H ,Cui M Y. 2013. Establishment and application of SCoT amplification system for macadamia[J]. Journal of Tropical and Subtropical Botany,21(3):253-258.]
陳紅,楊鑫,安華明. 2014. 貴州桃種質(zhì)資源遺傳多樣性的SCoT分析[J]. 西北植物學(xué)報,34(8):1559-1564. [Chen H,Yang X,An H M. 2014. Genetic diversity of peach acce-ssions in Guizhou analysed by SCoT markers[J]. Acta Botanica Boreali-Occidentalia Sinica,34(8):1559-1564.]
陳虎,何新華,黃桂香,李峰,姜建初,朱建華. 2012. 不同龍眼資源遺傳多樣性的SCoT和ISSR比較分析[J]. 廣西植物,32(4):536-541. [Chen H,He X H,Huang G X,Li F,Jiang J C,Zhu J H. 2012. Comparion and analysis of SCoT and ISSR markers for genetic diversity of longan[J]. Guihaia,32(4):536-541.]
陳虎,何新華,羅聰,高美萍,朱建華. 2009. 龍眼SCoT-PCR反應(yīng)體系的優(yōu)化[J]. 基因組學(xué)與應(yīng)用生物學(xué),28(5):970-974. [Chen H,He X H,Luo C,Gao M P,Zhu J H. 2009. The optimization of SCoT-PCR system of longan(Dimocarpus longan)[J]. Genomics and Applied Biology,28(5):970-974.]
陳虎,何新華,羅聰,朱建華,李峰. 2010. 龍眼24個品種的SCoT遺傳多樣性分析[J]. 園藝學(xué)報,37(10):1651-1654. [Chen H,He X H,Luo C,Zhu J H,Li F. 2010. Analysis on the genetic diversity of 24 longan(Dimocarpus longan) accessions by SCoT markers[J]. Acta Horticulturae Sinica,37(10):1651-1654.]
戴子云. 2010. 19個品種楊桃(Averrhoa carambola)對Cd的富集能力與修復(fù)潛力比較[D]. 廣州:中山大學(xué). [Dai Z Y. 2010. Cadmium accumulation and potential for phytoextraction of cadmium in 19 cultivars of Averrhoa carambola[D]. Guangzhou:Sun Yat-sen University.]
關(guān)長飛,胡超瓊,王孟珂,阮小鳳,王仁梓,楊勇. 2019. 陜西省柿種質(zhì)資源遺傳多樣性分析[J]. 植物遺傳資源學(xué)報. doi:10.13430/j.cnki.jpgr.20181213001. [Guan C F,Hu C Q,Wang M K,Ruan X F,Wang R Z,Yang Y. 2019. Genetic diversity analysis of persimmon germplasm resour-ces in Shaanxi Province[J]. Journal of Plant Genetic Resources. doi:10.13430/j.cnki.jpgr.20181213001.]
韓國輝,汪衛(wèi)星,向素瓊,邊禹,郭啟高,何橋,李曉林,梁國魯. 2011a. 多倍體枇杷SCoT分析體系的建立與優(yōu)化[J]. 果樹學(xué)報,28(3):433-437. [Han G H,Wang W X,Xiang S Q,Bian Y,Guo Q G,He Q,Li X L,Liang G L. 2011a. Establishment and optimization of SCoT system in polyploidy loquats[J]. Journal of Fruit Science,28(3):433-437.]
韓國輝,向素瓊,汪衛(wèi)星,賈志剛,洪棋斌,梁國魯. 2011b. 柑橘SCoT分子標記技術(shù)體系的建立及其在遺傳分析中的應(yīng)用[J]. 園藝學(xué)報,38(7):1243-1250. [Han G H,Xiang S Q,Wang W X,Jia Z G,Hong Q B,Liang G L. 2011b. Establishment and application of SCoT molecular marker system for Citrus[J]. Acta Horiiculturae Sinica,38(7):1243-1250.]
何江. 2017. 40份番石榴種質(zhì)資源親緣關(guān)系的形態(tài)學(xué)性狀和SCoT研究[D]. 南寧:廣西大學(xué). [He J. 2017. Morphological characters and SCoT analysis of genetic relationships among 40 accessions of guava germplasm[D]. Nanning:Guangxi University.]
和世玉. 2014. 庫爾勒香梨SCoT反應(yīng)體系優(yōu)化及優(yōu)良營養(yǎng)系鑒定[D]. 新疆:石河子大學(xué). [He S Y. 2014. Optimization of start codon targeted polymorphism(SCoT) system and identification of Korla Fragrant Pear excellent clones[D]. Xinjiang:Shihezi University.]
靳佳佳. 2018. 不同金柑品種親緣性及葉片氣孔和果皮精油差異性[D]. 南寧:廣西大學(xué). [Jin J J. 2018. Affinity and differences of blade stomatal and pericarp essential oil in defferent kumquat varieties[D]. Nanning:Guangxi University.]
劉鍇棟,袁長春,黎海利,陳春華,譚雪瓊. 2013. 楊桃遺傳多樣性的形態(tài)特征與RAPD標記的相關(guān)性分析[J]. 果樹學(xué)報,30(1):69-74. [Liu K D,Yuan C C,Li H L,Chen C H,Tan X Q. 2013. Correlation analysis of genetic diversity of carambola cultivars using morphological and RAPD markers[J]. Journal of Fruit Science,30(1):69-74.]
劉韜,車建美,黃素芳,廖汝玉,劉波. 2011. 福建省主要楊桃品種遺傳多樣性分析[J]. 果樹學(xué)報,28(3):448-452. [Liu T,Che J M,Huang S F,Liao R Y,Liu B. 2011. RAPD analysis of genetic diversity of carambola in Fu-jian Province[J]. Journal of Fruit Science,28(3):448-452.]
龍治堅. 2013. 枇杷屬植物的遺傳多樣性分析和指紋圖譜初步構(gòu)建[D]. 重慶:西南大學(xué). [Long Z J. 2013. Genetic diversity analysis and fingerprint construction of Eriobotrya on SSR and SCoT markers[D]. Chongqing:Southwest University.]
羅聰. 2012. 芒果SCoT分子標記與逆境和重要開花時間相關(guān)基因研究[D]. 南寧:廣西大學(xué). [Luo C. 2012. Study on SCoT markers and analysis on genes of stress-related and important flowering time in mango[D]. Nanning:Guangxi University.]
歐景莉,覃劍峰,余炳寧,黃小江,陸祖雙,朱鵬錦,陳豪軍. 2015. 楊桃SCoT標記PCR反應(yīng)體系建立與驗證[J]. 中國南方果樹,44(3):16-21. [Ou J J,Qin J F,Yu B N,Huang X J,Lu Z S,Zhu P J,Chen H J. 2015. Establishment and varification of the PCR protocol for SCoT in Averrhoa carambola L.[J]. South China Fruits,44(3):16-21.]
王發(fā)明,李潔維,葉開玉,龔弘娟,莫權(quán)輝,蔣橋生,劉平平. 2017. 41份葡萄種質(zhì)遺傳多樣性的ISSR和SCoT對比分析[J]. 廣西植物,37(1):1-8. [Wang F M,Li J W,Ye K Y,Gong H J,Mo Q H,Jiang Q S,Liu P P. 2017. Comparative analysis on the genetic diversity of 41 vitis germplasm resources by ISSR and SCoT molecular mar-kers[J]. Guihaia,37(1):1-8.]
熊發(fā)前,唐榮華,陳忠良,潘玲華,莊偉建. 2009. 目標起始密碼子多態(tài)性(SCoT):一種基于翻譯起始位點的目的基因標記新技術(shù)[J]. 分子植物育種,7(3):635-638. [Xiong F Q,Tang R H,Chen Z L,Pan L H,Zhuang W H. 2009. SCOT:A novel gene targeted marker technique based on the translation start codon[J]. Molecular Plant Breeding,7(3):635-638.]
楊君祎. 2015. 碭山酥梨授粉品種親緣關(guān)系的SCoT分析及對石細胞含量的影響[D]. 合肥:安徽農(nóng)業(yè)大學(xué). [Yang J Y. 2015. The effects on the stone cell content of Dangshansu pear based on genetic relationship of pollination cultivars by SCoT analysis[D]. Hefei:Anhui Agriculture University.]
楊祥燕,蔡元保,黃秋偉,陳濤,覃劍峰,彭靖茹. 2013. 番木瓜主栽品種SCoT指紋圖譜構(gòu)建及遺傳變異分析[J]. 西北植物學(xué)報,33(9):1756-1761. [Yang X Y,Cai Y B,Huang Q W,Chen T,Qan J F,Peng J R. 2013. SCoT fingerprints and genetic variations of papaya(Carica papaya L.) major cultivars[J]. Acta Botanica Boreali-Occidentalia Sinica,33(9):1756-1761.]
Collard B C Y,Mackill D J. 2009. Start codon targeted(SCoT) polymorphism:A simple,novel DNA marker technique for generating gene-targeted markers in plants[J]. Plant Molecular Biology Reporter,27(1):86-93.
Guo D,Zhang J,Liu C. 2012. Genetic diversity in some grape varieties revealed by SCoT analyses[J]. Molecular Biology Reports,39(5):5307-5313.
Luo C,He X,Chen H,Ou S,Gao M. 2010. Analysis of diversity and relationships among mango cultivars using start codon targeted(SCoT) markers[J]. Biochemical Syste-matics and Ecology,38(6):1176-1184.
Vanhala T,Tuiskula-Haavisto M,Elo K,Vilkki J,Maki-Tanila A. 1998. Evaluation of genetic variability and genetic distances between eight chicken lines using microsatellite markers[J]. Poultry Science,77(6):783-790.
(責(zé)任編輯 陳 燕)
