不同進口白霉奶酪中優勢霉菌分離鑒定及其凝乳酶酶學特性研究
2019-09-10 09:21:04沈陽農業大學食品學院沈陽110866沈陽市農業綜合行政執法支隊沈陽110031
中國乳品工業 2019年7期
(1.沈陽農業大學食品學院,沈陽110866;2.沈陽市農業綜合行政執法支隊,沈陽110031)
0 引言
奶酪富含多種對人體健康有益的營養物質[1,2],鈣含量突出[3-4],是全球重要的乳制品之一[5]。但我國奶酪產品主要依賴進口,普遍存在口味單一和品質同質化問題[6]。已有眾多研究選用不同霉菌制作奶酪,以豐富奶酪口味[7-10]。白霉奶酪典型特征為奶酪表面有一層白色霉菌,營養價值極高[11-13]。白地霉在食品、釀造、飼料等行業已有長期研究[14],其在牛奶、奶酪等乳制品中自然存在[15],對奶酪質構、風味、香氣等貢獻明顯[11,16-17]。近年來,有關奶酪風味[18]、作用機理[19]、分子鑒定[20]等研究較為突出,微生物源凝乳酶酶學特性研究日益廣泛[21-24]。基于此,本研究從進口白霉奶酪中篩選優勢霉菌進行鑒定,并研究該凝乳酶的酶學性質,以期為白地霉在霉菌奶酪生產中的應用提供基礎資料。
1 材料與方法
1.1 材料與設備
1.1.1 實驗材料
白霉奶酪,生產商Nordex Food;Danish Brie cheese白霉奶酪,生產商Arla Foods amba Troldhede Dairy Y;Danish Camembert cheese白霉奶酪,生產商北京申囤貿易有限公司。
1.1.2 培養基
麥芽汁瓊脂培養基:麥芽膏20 g,蛋白胨1.0 g,葡萄糖20 g,瓊脂20 g,蒸餾水1 000 m L。麥芽汁液體培養基:10~15°P新鮮麥芽汁1 000 mL,自然pH值,1×105Pa滅菌20 min。5°P麥汁培養基:5度麥芽汁,2%瓊脂,1×105Pa滅菌20 min。馬鈴薯葡萄糖瓊脂(PDA)培養基:馬鈴薯200 g,葡萄糖20 g,瓊脂15-20 g,蒸餾水1 000 mL。制法:將馬鈴薯去皮切塊,加1 000 mL蒸餾水,煮沸10-20 min。用紗布過濾,補加蒸餾水至1 000 mL。加入葡萄糖和瓊脂,加熱溶化,自然p H值,1×105Pa滅菌20 min。
1.2 儀器與設備
Nikon TS100型倒置熒光顯微鏡,AYU-12O型分析天平,HCB-1300V型垂直層流潔凈工作臺,電子天平,LDZF-30KB型立式壓力蒸汽滅菌器,上海一恒THZ-98A型恒溫振蕩器,上海精宏SHP-250型生化培養箱,DK-8D型電熱恒溫水槽,3-30K型低溫高速離心機。
1.3 實驗方法
1.3.1 優勢霉菌篩選分離
(1)采樣。在垂直層流潔凈工作臺打開奶酪外包裝,用滅菌切割刀將整塊奶酪不同部分切割成小片,平鋪放置于加有抗生素的PDA培養基中,28°C培養24 h,定期觀察菌種培養過程并記錄,選擇發霉量大,生長旺盛的培養基平板進行后續實驗。
(2)增殖培養。無菌操作條件下,接種環挑取上述菌種物置于麥芽汁液體培養基,28°C恒溫箱中培養24 h。
(3)平板分離。無菌操作將增殖培養后的菌種稀釋為10-1濃度菌液,再依次稀釋為10-2,10-3,10-4,10-5,10-6濃度梯度菌液。分別吸取0.1 mL各濃度梯度菌液至麥芽汁瓊脂平板中涂布,置于28°C恒溫培養箱中培養定期觀察菌落形態特征并記錄。
(4)初篩。將培養好的平皿選取單個分散菌落,在無菌條件下,進行第一次、第二次平板分離,接種針挑取第二次平板分離的菌株,進行麥芽汁瓊脂培養基劃線培養,每株菌株劃線三個平板,記錄并觀察菌株生長情況及菌落形態特征,選取生長能力較強菌株。
(5)復篩。將初篩后得到的菌株接種于添加抗生素、無外源污染物的麥芽汁瓊脂培養基,于28°C培養5 d后,觀察菌株生長能力,保留生長能力較強菌株。
1.3.2 優勢霉菌菌種鑒定與保藏
(1)常規鑒定。對篩選得到的生長能力較強、菌落旺盛菌株進行形態學和理化特性分析。形態學包括顯微形態和培養特征,顯微鏡觀察根據篩分菌種的菌絲有隔或無隔;菌絲體透明或暗黑,有色或無色;是否產生有性孢子,孢子的類型;無性孢子的類型等對霉菌進行初步鑒定。初步鑒定菌株通過碳源同化試驗、氮源同化試驗、水解蛋白、分解楊梅苷測定及分解脂肪試驗進行理化特性分析鑒定,并參照《真菌鑒定手冊》進行鑒定[25]。
(2)分子生物學鑒定。a.菌種培養:取4°C低溫冷藏菌種,連續2次在PDA培養基活化,轉接到5°P麥芽汁固體平板上,30°C過夜培養,挑取一環菌種,加入25 mL、5°P麥芽汁培養基,于28°C、150 r/min條件下培養48 h。b.基因組提取:篩分菌株基因組DNA提取CTAB法,使用0.8%瓊脂糖凝膠電泳檢驗DNA純度和完整性。c.26SrDNA D1/D2區序列擴增:以菌株基因組DNA為模板,用引物5′-GGTGACGCAG-3′進行PCR擴增。反應體系:10×PCR Buffer5μL,MgCl2(25 mmol/L)4μL,d NTPs(2.5 mol/L)4μL,隨機引物(5μmol/L)2μL,模板DNA2μL,Taq酶(5 U/μL)0.6μL,用ddH2O補至50μL;采用分段PCR擴增程序:94°C 5 min,(94°C 3 min,35°C 1.5 min,72°C 2 min)×5個循環,(94°C 30 s,35°C 1 min,72°C 1.5 min)×30個循環,72°C 5 min。擴增產物用1.5%瓊脂糖凝膠電泳檢測,并拍照記錄結果。d.序列測定與比對分析:將圖譜結果與模式菌株DNA進行同源性比較,獲得菌株相似度。
(3)鑒定菌種保藏。菌種短期保藏采用斜面保藏法,分離得到純菌株移接到麥芽汁斜面,放置于28°C恒溫箱中培養24 h后置于冰箱4°C存放。菌種長期保藏采用甘油超低溫保藏,分離得到純菌株搖瓶培養后獲得菌懸液,取菌懸液與60%甘油各500μL于保菌管中混合均勻,置于-80°C超低溫冰箱存放。
1.3.3 優勢霉菌凝乳酶活性及酶學特性測定
(1)保藏菌種活化。將冷凍保藏優勢霉菌接種于PDA斜面培養基進行活化培養,28°C培養3~4 d,待培養基中長滿菌絲后備用。
(2)孢子懸浮液制備。
用10 mL無菌生理鹽水沖刷生長良好的試管斜面種子,將沖刷液移入裝有生理鹽水和玻璃珠的三角瓶中,在搖床上振蕩打散,并用帶有脫脂棉的漏斗進行過濾。
(3)凝乳酶分離與提取
將1 mL優勢霉菌孢子懸浮液接種于發酵培養基,28°C培養3~4 d,培養完成后,加入100 mL蒸餾水,在20°C、180 r/min振蕩浸提1 h,浸提液用4層紗布過濾,濾液在10 000 r/min條件下離心30 min。在4°C向所得上清液中加入預冷的無水乙醇,使無水乙醇體積分數分別為20%、30%、40%、50%、60%、70%和80%,靜置24 h后離心30 min,用5 mmol/L磷酸鹽緩沖液溶解沉淀,分別測定凝乳酶活性,選取凝乳酶活力最高時無水乙醇的體積分數進行凝乳酶提取。
(4)凝乳酶活性及酶學特性測定
參照Arima[26]方法,用0.01 mol/L的CaCl2溶液將脫脂乳溶解,配制成10%的脫脂乳溶液,室溫下靜置30 min后,各量取5 mL分裝到小試管中,35°C恒溫水浴5 min,在35°C下取待測酶液0.5 mL加入到5 mL裝有10%脫脂乳溶液的試管中,立即搖勻,開始計時,出現絮狀沉淀停止計時,并計算凝乳酶活性。
凝乳酶最適反應溫度:配置濃度為10%的脫脂乳溶液,取同量的酶液,分別在30°C、35°C、40°C、45°C、50°C、55°C、60°C、65°C、70°C保溫10 min,測定凝乳酶活性,確定凝乳酶最適反應溫度。
凝乳酶最適反應pH:用濃度為0.1 ml/L的氫氧化鈉溶液和0.1 ml/L鹽酸溶液將酶液的pH分別調整為4.0、4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0、8.5、9.0、10.0,在最適反應溫度下保持10 min,測定凝乳酶活性,確定凝乳酶最適反應pH。
凝乳酶穩定性:取同量的酶液,在最適反應溫度和pH下反應3、5、7、10、15、20和30 min,測定凝乳酶活性,考察凝乳酶穩定性。
金屬離子對凝乳酶活性影響:配置不同濃度的金屬離子溶液,按照一定比例加入到脫脂乳溶液中,使得脫脂乳中金屬離子濃度為0.001 mol/L,所選擇的金屬離子包括Fe2+、Cu2+、Mg2+、Co2+、Zn2+、Al3+、Ca2+、Fe3+、Mn2+金屬離子,在最適溫度、p H條件下催化反應,測定凝乳酶活性,考察金屬離子對凝乳酶影響。
1.4 數據處理
用Excel軟件整理數據和繪圖,使用SPSS 22.0軟件對數據進行方差分析和相關性分析,方差分析選取Duncan檢驗,在p<0.05檢驗水平上對數據進行統計學分析。
2 結果
2.1 菌種篩選分離及鑒定
2.1.1 菌種篩選分離
篩分培養期間觀察得到4個奶酪平板具有較強的生霉能力,分別在奶酪表面長有白色絨毛狀兼粉狀菌絲,菌絲緊貼奶酪表面,平坦且分布均勻,菌絲叢細密度相接近。培養開始第二天有菌絲出現,在第三天到第四天菌絲生長旺盛,形成大面積菌落,菌絲叢大多呈圓形,顏色主要為白色,部分略帶微黃色。篩分得到的菌株其菌落形態特征如圖1。
優勢菌株的菌落形態如圖1所示,菌株在麥芽汁瓊脂培養基中28°C培養3 d,菌落正面觀看呈圓形,較薄,正面顏色白色,平坦,邊緣不整齊,中心略微凸起,呈放射狀,略有微絨狀,菌絲質地粉狀,背面觀看呈圓形,黃白色,邊緣不整齊,邊緣色淺,呈放射狀。
從篩選獲得45個菌落選取1個單菌落進行優勢菌株制片進行細胞形態觀察,結果如圖2。

圖1 菌落形態
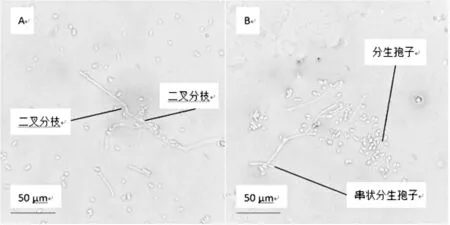
圖2 菌株顯微形態
由圖2可以觀察到,分離所得菌株細胞形態,菌體產生真菌絲,菌絲匍匐,有橫隔,菌絲以二叉分枝形式生長,菌絲無色。孢子梗頂端生有長串狀的分生孢子,分生孢子5.4μm×4μm,呈圓筒狀或長筒形,近乎長方形,兩端多圓形,少數平切,無色透明,單孢,有時連接成孢子鏈。
2.1.2 菌種理化鑒定
菌株對于試驗選用7種不同唯一碳源培養基表現出不同的利用結果。碳源同化試驗測定結果如表1所示,菌株能夠在D-核糖、蔗糖、麥芽糖、乳糖、核糖醇作為唯一碳源培養基中正常生長,但在不同碳源化合物周圍的霉菌菌落數目存在差異,說明菌株對D-核糖、蔗糖、麥芽糖、乳糖、核糖醇的利用程度存在一定差異,對碳源種類有一定要求。而在纖維二糖、木糖醇作為唯一碳源培養基中不能生長,說明菌種對碳源的利用具有選擇性。

表1 碳源同化結果
氮源同化試驗測定結果如表2所示,菌株在蛋白胨、尿素、硫酸銨作為唯一氮源培養基中正常生長,但在蛋白胨作為培養基中唯一氮源時,菌體生長迅速,菌絲濃密,菌落特征明顯。而在硝酸鉀作為唯一氮源培養基中不能生長,菌株對有機氮源的利用效果優于無機氮源。

表2 氮源同化結果
水解蛋白質、分解楊梅苷、脂肪試驗測定結果如表3所示,菌株不能分解楊梅苷,能分解脂肪和蛋白質。霉菌菌株中包含多種酶系,其中蛋白酶和脂肪酶表現出對蛋白質和脂肪較強的分解能力,其揮發性產物對奶酪風味形成具有重要作用。

表3 其他生理實驗結果匯總表
2.1.3 菌種分子生物學鑒定
菌株進行基因分子水平鑒定,將其26S rDNA D1/D 2區序列同NCBI庫中序列進行比對,其結果顯示測定菌株與Geotrichum sp.(白地霉屬)同源性高達99%以上,與其他物種序列相比得分最高,具有明顯物種差異。本序列測定使用該菌株的脂肪酸脫飽和酶基因作為特征序列,其GenBank編號為MH 198047。
2.2 凝乳酶酶學特性
2.2.1 溫度對凝乳酶活性影響
溫度主要影響凝乳酶對酪蛋白作用的第二步即影響酪蛋白膠束的聚集作用。在反應溫度分別為30°C、35°C、40°C、45°C、50°C、55°C、60°C、65°C和70°C條件下測定了其活性。從圖3結果中可以看出白地霉凝乳酶的活性隨著溫度的升高逐漸提高,在反應溫度為45°C時其活性達到(0.495±0.067)IMCU。而后隨溫度的升高凝乳酶活性有下降的趨勢,在45°C~65°C時,凝乳酶活性緩慢下降,在70°C凝乳酶活性比45°C降低了45.4%。反應溫度在30°C~40°C和55°C~70°C之間,溫度對凝乳酶活性影響不存在顯著性差異。在45°C~50°C范圍內凝乳酶活性未見明顯損失,說明白地霉凝乳酶在45°C~50°C內具有較強的穩定性和凝乳效果。
白地霉凝乳酶最適反應溫度為45°C,此結果與Rajesh Kumari Narwal等[27]所報告的Bacillus subtilis MTCC 10422凝乳酶最適溫度一致。Lmdaki等[28]研究結果表明,提取自Pediococcus acidilactici SH菌株的凝乳酶最高活性在50°C時不受抑制,而在60°C時沒有活性。我們的結果表明凝乳酶在70°C活性依然存在,這可以解釋Ahmed and Helmy[29]發現最大的地衣芽孢桿菌5A5凝乳酶最高活性在75°C的可能性,體現了白地霉凝乳酶作用溫度范圍的廣泛。溫度作為影響酶活性的重要條件,在溫度過低時酶活性受到抑制,溫度過高時凝乳酶三維空間結構易被破壞而發生變性,導致在低溫和高溫條件下凝乳酶活性相對較低。白地霉凝乳酶最適溫度為45°C,說明在此溫度下凝乳酶與底物結合最佳,酶的空間構象未被改變,凝乳酶表現出相對較高的活性。
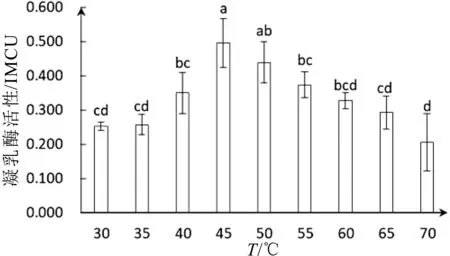
圖3 溫度對凝乳酶活性影響
2.2.2 p H值對凝乳酶活性影響
在最適反應溫度45°C條件下,pH值對凝乳酶活性影響如圖4所示。從圖中可以看出,白地霉凝乳酶在pH 6.0時活性最高,達到(0.613±0.043)IMCU。而后隨著p H值的增加白地霉凝乳酶活性呈下降趨勢,當p H值為8.5時凝乳酶活性僅為p H=6.0時的27.0%。pH在4.5-5.0、6.5-7.5和8.5-10.0之間,凝乳酶活性沒有顯著性差異,在pH為9.0和10.0時凝乳酶活性比p H=6.0時分別降低71.5%和67.2%。從圖中還可以看出,白地霉凝乳酶在酸性條件活性高于堿性條件,這與凝乳酶是酸性蛋白酶相吻合。在pH 4.0-6.0之間凝乳酶活性上升平緩,說明白地霉凝乳酶對酸耐受性較好,在酸性條件具有相對較高的穩定性;而在p H 7.0-8.5范圍內,凝乳酶活性有下降的趨勢,波動較明顯。
白地霉凝乳酶最適反應pH值為6.0,此結果與Poza M等[30]所報告的Myxococcus xanthus strain 422凝乳酶最適p H 6.0一致。p H值對酶活性的影響主要為破壞酶的空間結構,影響酶活性部位催化基團的解離狀態和影響底物解離狀態。在過酸或過堿條件下凝乳酶的空間構象將被破壞,影響酶分子構象的穩定性,導致酶活性降低。而在p H 6.0時,凝乳酶分子上的極性基團受到低酸性環境的影響,處于較為穩定的解離狀態,此時凝乳酶與底物結合表現出較高的活性。
2.2.3 反應時間對凝乳酶活性影響
凝乳酶穩定性對奶酪生產加工具有現實意義,在奶酪的加工成熟過程中凝乳酶起著極其重要的作用,凝乳酶的穩定性將會影響奶酪的質構和風味。在反應溫度為45°C、pH=6.0的條件下,將白地霉的凝乳酶分別保溫3、5、7、10、15、20和30 min后,測定了其活性。結果如圖5所示,凝乳酶活性隨作用時間增加緩慢降低。在3 min時具有最高活性(0.613±0.043)IMCU,5~7 min凝乳酶活性略有降低,分別為(0.549±0.048)IMCU、(0.536±0.031)IMCU,其活性分別保留了89.6%和87.4%。而在20-30 min其活性顯著下降,分別為(0.347±0.041)IMCU、(0.251±0.057)IMCU,其活性分別保留了56.6%和40.9%。白地霉凝乳酶在實際應用中,應保持反應時間在3~7 min內,才能夠發揮凝乳酶最佳凝乳作用,保持相對較高的穩定性[31]。因此,該凝乳酶具備較好的凝乳穩定性。

圖4 pH值對凝乳酶活性影響
白地霉凝乳酶在3~7 min相對穩定,酶活性變化不顯著,凝乳酶在短時間內受到溫度和pH影響不明顯,凝乳酶對環境具有一定耐受性,酶分子的空間結構、解離狀態、催化活性都較為穩定,能夠保持較高的活性。但長時間處于恒定溫度和p H環境下,凝乳酶活性也會有所下降,穩定性發生波動。7~10 min為酶活性顯著下降階段,此時溫度和pH對凝乳酶活性影響累積作用達到臨界點,酶活性發生突變,10~20 min凝乳酶作用環境不足以使酶分子解離狀態發生較大變化,活性下降較為平緩,直至30 min環境累積作用進一步加強,凝乳酶解離狀態大大增加,酶活性波動較大,下降明顯。
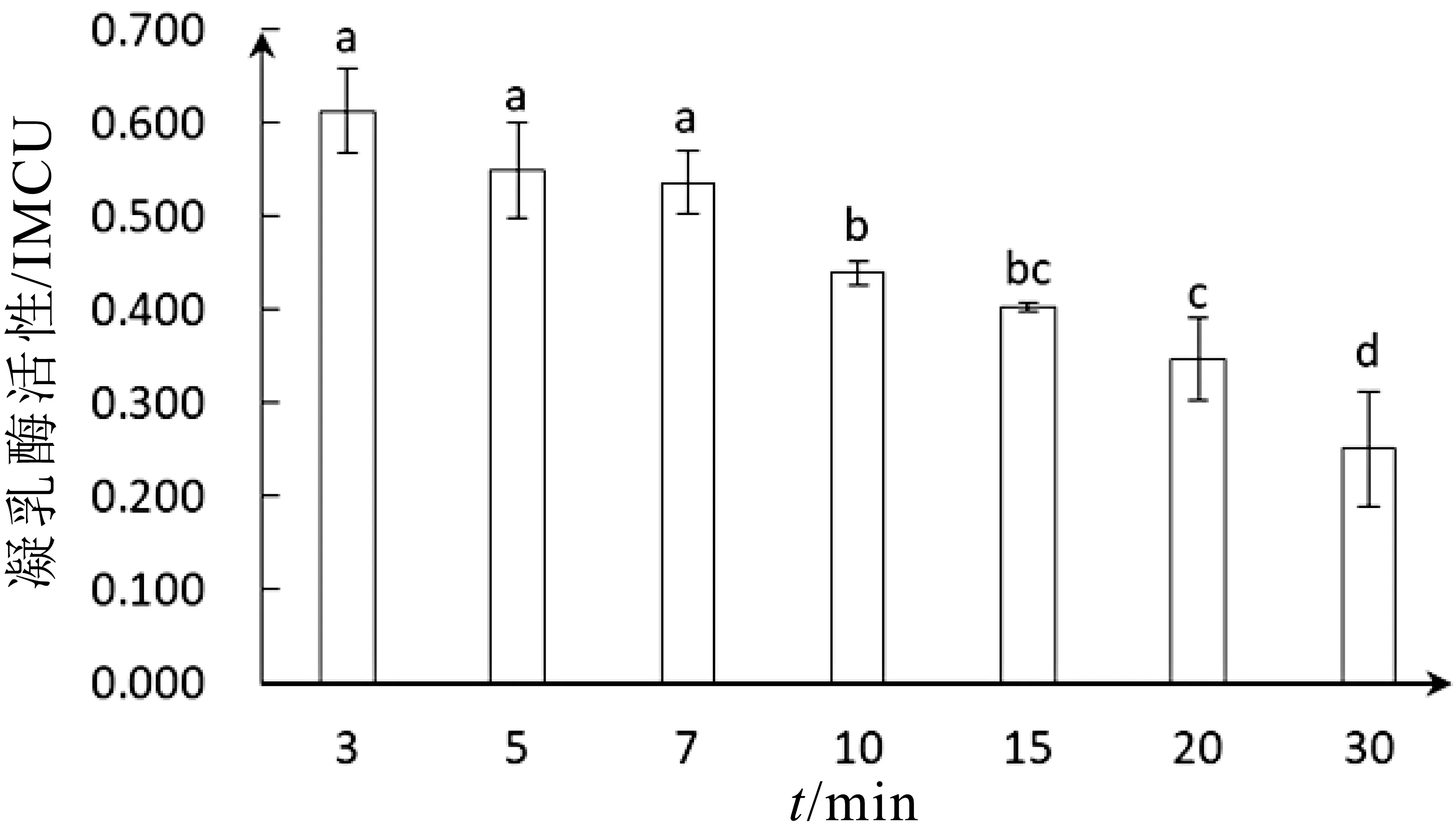
圖5 反應時間對凝乳酶活性影響
2.2.4 金屬離子對凝乳酶活性影響
在反應溫度為45°C、pH=6.0、作用時間3 min條件下,不同金屬離子對凝乳酶活性影響如圖6所示。由結果可知,不同金屬離子對白地霉凝乳酶活性具有較大影響。Fe2+、Ca2+、Fe3+、Mn2+能夠降低凝乳酶活性,Fe3+對凝乳酶活性有極大抑制作用,其活性降低至(0.175±0.070)IMCU僅為原來的19.9%。Mn2+對凝乳酶活性抑制作用較小,其活性降低至(0.594±0.061)IMCU。Cu2+、Mg2+、Zn2+、Al2+能夠提升凝乳酶活性,其活性分別達到(0.812±0.055)IMCU、(0.880±0.092)IMCU、(0.639±0.034)IMCU和(0.664±0.024)IMCU,與對照組相比分別提高32.5%、43.6%、4.2%和8.3%。由此可知,Mg2+和Cu2+對提高白地霉凝乳酶活性有顯著影響。
白地霉凝乳酶在不同金屬離子存在時,酶活性變化較大。凝乳酶和底物因均帶負電荷而相互排斥,不同金屬離子因所帶正電荷數量不同將會對它們之間的排斥作用產生不同程度的影響,引起酶活性變化。不同金屬離子可以作為輔基,通過與酶分子結合影響酶活性,Mg2+和Cu2+與酶的結合更加牢固,凝乳酶表現出較高的活性,而Fe3+常作為血紅蛋白的輔基離子,不易與凝乳酶結合,體現出較低的酶活性。同時,有報道表明凝乳酶對酪蛋白的Phe105-106鍵呈高度專一性,Fe3+將會影響к酪蛋白的構象[32],導致凝乳酶活性降低。
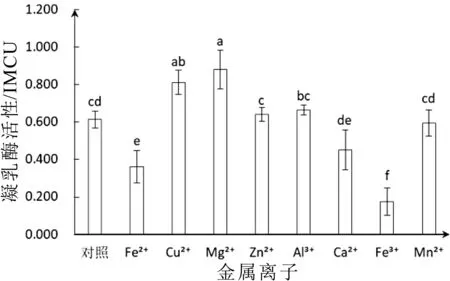
圖6 金屬離子對凝乳酶活性影響
3 結論
本研究從進口奶酪中分離篩選得到優勢霉菌,并鑒定該優勢霉菌為白地霉。考查了白地霉凝乳酶在不同溫度、不同pH值、作用時間以及不同金屬離子條件下的酶活性,確定其最佳反應溫度為45°C,最適pH 6.0,最適反應時間3 min,最優結合金屬離子為Mg2+和Cu2+。本研究為提高白霉奶酪品質和生產應用提供數據基礎。
