新疆伊犁地區乳品中發酵菌種的篩選及產酸性能研究
2019-09-10 09:21:02張亞川蔡靜靜剡文莉王麗軍倪永清
中國乳品工業 2019年7期
關鍵詞:實驗
張亞川,蔡靜靜,剡文莉,王麗軍,倪永清
(石河子大學食品學院食品生物技術實驗室,新疆石河子832000)
0 引言
伊犁州是我國重點牧區之一[1]。當地居民制作的傳統乳制品中蘊藏著豐富的乳酸菌資源。許多學者對新疆傳統乳制品中的乳酸菌進行了研究,董曉婉等[2]從新疆和布克賽爾縣25份傳統酸乳中分離出40株乳酸菌。劉潔潔等[3]從阿勒泰地區傳統乳制品中分離出11株乳酸菌。馬燕等[4]從伊犁、石河子地區分離出104株乳酸菌。
傳統的乳酸菌鑒定方法依賴于表型分析[5]。16S rDNA是細菌分類研究中最常用的分子鐘[6]。隨著分子生物學技術的發展,使用16SrRNA序列分析方法鑒定LAB越來越普遍。
我國不同區域的自然發酵乳品承載了當地豐富的微生物資源,因此應深入研究并從中篩選出優良乳酸菌用于工業化生產[7]。伊犁地區傳統乳制品中微生物數量眾多,對其中優良發酵菌株的開發能夠為酸乳發酵劑的制作提供菌株。
1 實驗
1.1 材料及試劑
(1)實驗樣品。采自新疆伊犁地區的傳統奶制品。
(2)培養基。MRS肉湯培養基,用于乳桿菌的富集;MRS固體培養基(MRS肉湯添加1.7%的瓊脂),用于乳桿菌的分離篩選;M 17肉湯培養基,用于乳球菌的富集;M 17固體培養基(M 17肉湯添加1.7%的瓊脂),用于乳球菌的分離篩選。上述培養基121℃滅菌15 min使用。質量分數為12%脫脂乳培養基,95℃滅菌5 min使用。
(3)試劑。Easy Pure Bacteria Genomic DNA Kit,PCR擴增體系所需試劑以及擴增引物。
1.2 儀器
恒溫生化培養箱,PCR儀,水平電泳儀為,電泳槽,凝膠成像系統等。
2 方法
2.1 乳酸菌的分離鑒定
2.1.1 乳酸菌的分離純化
樣品用無菌生理鹽水稀釋至10-4,10-5,10-6;吸取100 L稀釋后的樣品均勻的涂布在MRS、M 17培養基上[8]。觀察菌落顏色、大小、凸起程度和光澤度,挑選其中具有乳酸菌典型特征的單菌落進行富集。取適量菌懸液進行鏡檢,若鏡檢中菌株細胞形態、排列方式不一致,則需繼續劃線純化至一致。純化后的菌株37℃條件下培養18 h后進行革蘭氏染色。將革蘭氏陽性菌株富集后以2%的接種量接種到12%的滅菌脫脂乳中37℃培養,具備凝乳特性的菌株保藏至-80℃備用。
2.1.2 生理生化實驗
參照“東秀珠”等《常見細菌鑒系統定手冊》[9]。
2.1.3 乳酸菌16SrRNA序列分析
(1)菌株DNA的提取。菌體DNA的提取采用Easy Pure Bacteria Genomic DNA Kit試劑盒,使用試劑盒所提供的藥品按照說明書步驟提取DNA。
(2)16S rRNA序列擴增。擴增引物采用乳酸菌通用引物,上游引物27F:5′-AGA GTT TGA TCC TGG CTC AG-3′,下游引物1492R:5′-TAC CTT GTT ACG ACT T-3′。
PCR擴增體系(25 L體系):預混液2×Taq MasterMix 12.5 L,引物27F 0.5 L,引物1492R 0.5 L,DNA模板2 L,補充ddH2O至25 L。PCR反應條件:94℃預變性5 min;95℃變性1 min,57℃退火1 min,72℃延伸1 min,35個循環;72℃終延伸7 min;4℃保存備用。
(3)PCR擴增產物檢測。PCR擴增產物使用瓊脂糖凝膠電泳檢測,100 V電泳40 min,凝膠成像儀下采集圖像。
(4)系統發育分析。菌株16S rRNA序列擴增成功后,委托上海生工生物工程技術服務有限公司進行測序,將測序結果在NCBI數據庫進行BLAST在線分析。將測序菌株以及模式菌株的16SrRNA序列經Clustal X多重比對后,運用MEGA 6.0軟件對各菌株的系統發育地位進行分析。
2.2 發酵菌種的篩選
2.2.1 凝乳時間的確定
將37℃條件下培養18 h的乳酸菌在轉速為4 000 r/min下離心10 min后,棄去上清液并用滅菌生理鹽水洗滌,反復洗滌3次后用生理鹽水將離心菌體重懸。以2%的接種量將菌種接種于滅菌脫脂乳中,置于37℃恒溫培養至其凝乳,保存至4℃冰箱作為發酵劑備用[10](以下實驗所用酸奶發酵劑均按此方法制備)。將單菌株發酵劑以2%的添加量接種于滅菌脫脂乳中,置于37℃下恒溫培養,每隔1 h檢查凝乳狀況,待樣品凝乳后記錄凝乳時間,并將酸乳置于4℃冷藏柜保藏待用。
2.2.2 酸度測定
取5.0 g冷藏的發酵乳樣品于100 mL錐形瓶中,加入40 mL煮沸蒸餾水混合均勻,并加入5滴0.5%的酚酞指示劑,用0.1 mol/L的標準氫氧化鈉溶液滴定至粉紅色,1 min內不變色。記錄氫氧化鈉的消耗量及酸乳的質量[11],其計算公式為
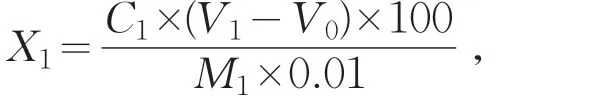
式中:X1為試樣的酸度;C1為氫氧化鈉濃度;V1為樣品消耗氫氧化鈉體積;V0為空白消耗氫氧化鈉體積;100為100 g試樣;M1為試樣的質量;0.1為酸度理論定義氫氧化鈉的濃度。
2.2.3 感官評定
邀請10名食品專業的研究生對冷藏24 h的發酵乳從色澤、風味、組織狀態方面進行感官特性評價,各項總分為色澤10分、風味40分、組織狀態50分。取出冷藏酸乳觀察其組織狀態和色澤,再聞其氣味,用純凈水漱口后,品嘗樣品的滋味,并記錄對應項得分。
2.3 菌株產酸性能
2.3.1 生長性能
采用比濁法測定菌株的生長曲線。活化后的菌株以2%的接種量接種于裝有150 mL液體MRS培養基的200 mL錐形瓶中,37℃條件下培養30 h,每隔2 h測其OD600值。繪制OD600值與培養時間的曲線[12]。
2.3.2 菌株產酸能力
單菌株發酵劑以2%的接種量接種于脫脂乳中,37℃恒溫培養,每隔2 h取出樣品測其滴定酸度,直至滴定酸度趨于穩定。繪制酸乳發酵過程中酸度隨時間變化的曲線。酸度測定方法以及發酵劑制備方法見上述。
2.3.3 菌株后酸化能力
單菌株發酵劑以2%的接種量接種于滅菌脫脂乳中,發酵至終點后將發酵乳取出置于4℃保藏,分別于第1,5,10,15,20 d取出測其滴定酸度。酸度測定方法以及發酵劑制備方法見上述。
3 結果與討論
3.1 乳酸菌的分離鑒定
3.1.1 乳酸菌的分離純化
從采集的酸牛乳、酸駝乳以及奶酪中初步分離出具有明顯乳酸菌特征的菌株71株,其中具有產酸凝乳特性的乳酸菌8株,8株乳酸菌的顯微鏡檢圖片以及菌體形態特征如圖1和表1所示。

圖1 乳酸菌顯微特征

表1 菌落形態以及菌株形態表征
3.1.2 生理生化實驗
8株乳酸菌的生理生化結果如表2所示。由表2可以看出,8株乳酸菌接觸酶實驗、明膠液化實驗、硫化氫實驗、硝酸鹽還原實驗、運動性實驗的結果均為陰性。糖醇發酵實驗中,所有菌株均能利用葡萄糖和乳糖發酵產酸;果糖實驗中,只有菌株BLK7-10和BLK7-12呈 陰 性;菌 株 BLK7-10,BLK7-12,XKN 1-5,BST 4-1可以利用蔗糖發酵產酸;菌株BLK7-10,XKN 1-5,BST4-1可以利用麥芽糖發酵產酸;阿拉伯糖發酵實驗中,只有菌株BLK7-10呈陽性。菌 株BLK7-10,BST 4-1,BST 2-3,BSTS6-1,BSTS6-3,BSTS6-4與乳桿菌的特征相似,初步判斷上述6株菌為乳桿菌。菌株XKN 1-5和BLK7-12與乳球菌的特征相似,初步判斷菌株XKN 1-5和BLK7-12為乳球菌。
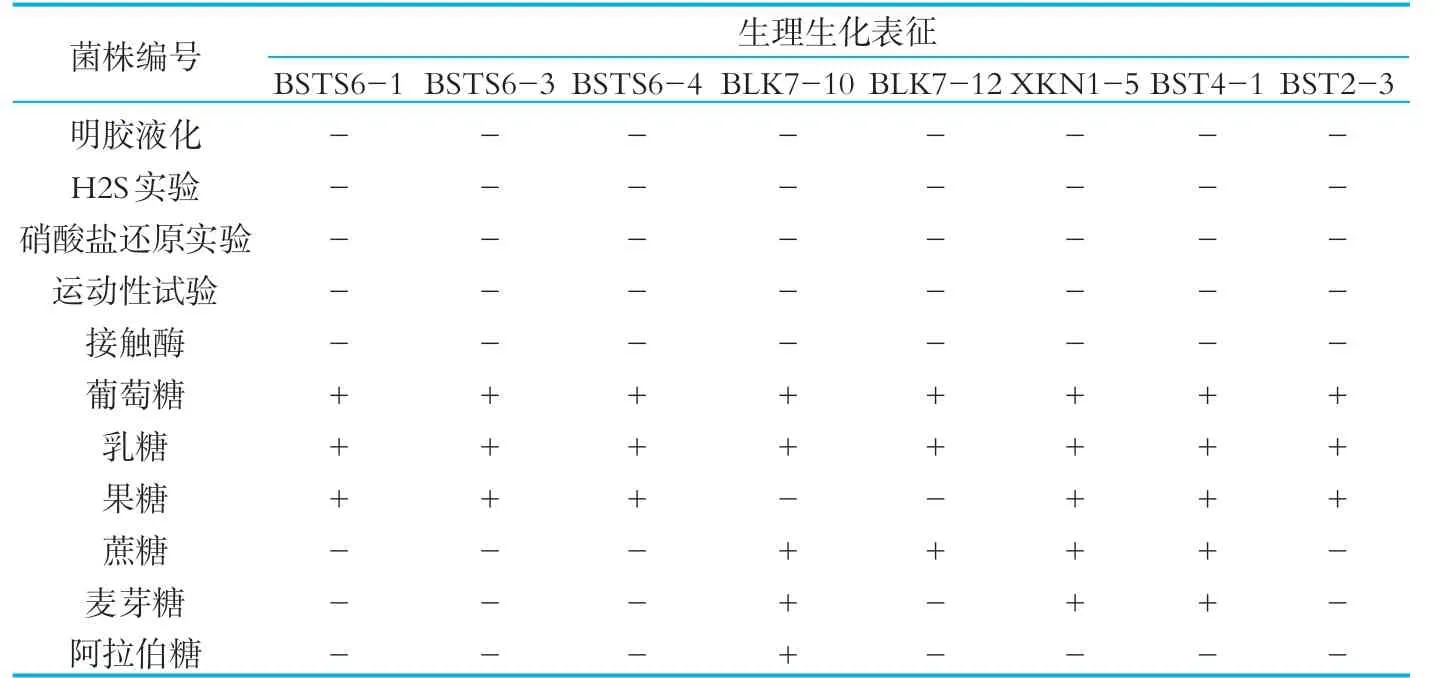
表2 生理生化鑒定
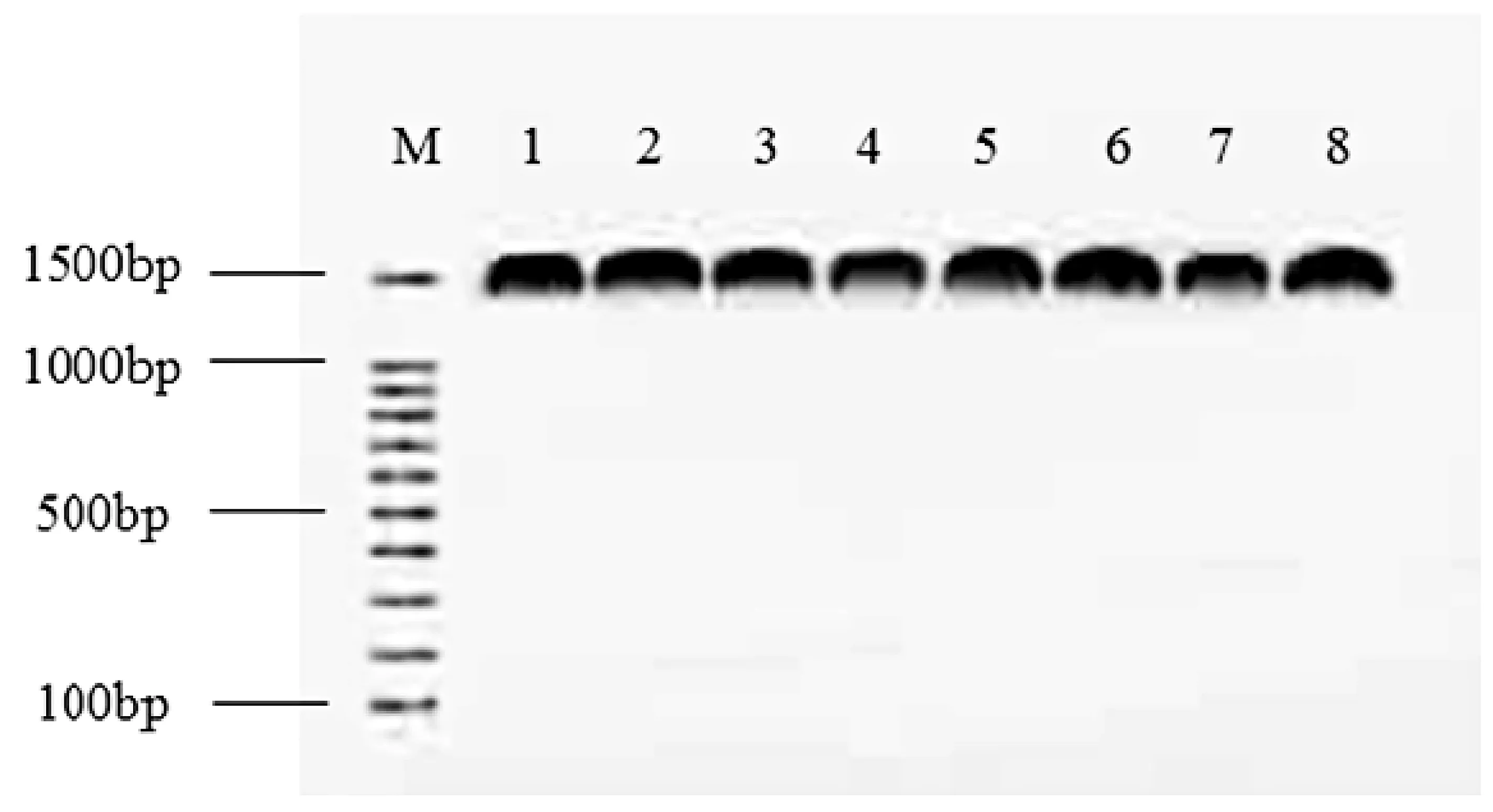
圖2 PCR產物凝泳膠電圖
3.1.3 16SrRNA序列分析
所篩選菌株的PCR擴增產物如圖2所示。由圖2可以看出,所有菌株的PCR擴增產物均在1 500 pb左右出現特異性擴增帶,與預期大小相符,說明擴增成功。將8株菌的PCR產物送至上海生工生物科技有限公司完成相關測序工作。
3.1.4 系統發育分析
將8株乳酸菌的部分16SrRNA基因序列將測序結果在NCBI數據庫進行BLAST在線分析,比對結果如表3所示。選取同源性最高的序列構建系統發育樹,系統發育樹如圖3所示,由圖3可以看出,7株乳酸菌屬于Lactobacillus,Leuconostoc,Enterococcus三個屬;其中菌株BSTS6-1,BSTS6-3,BSTS6-4與Lactobacillus delbrueckii subsp.bulgaricus 57-1,Lactobacillus delbrueckii subsp.bulgaricus 9-4,Lactobacillus delbrueckii subsp.bulgaricus YLj15在一個分支上,且相似度達到99%,可確定3株菌為德氏乳桿菌保加利亞亞種;菌株BLK7-10與Lactobacillus kefiranofaciens KF16在一個分支上;菌株BLK7-12與Enterococcus faecalis G1在同一分支上,且相似度高達99%,判定菌株BLK7-12為糞腸球菌;菌株XKN 1-5與Leuconostoc lactis MYSN 98在同一個分支上,相似度達99%,判定菌株XKN 1-5為乳酸明串球菌;菌株BST4-1、BST2-3分別于Lactobacillus plantarum AS-6、Lactobacillus fermentum LDTM在同一分支上,相似度均達100%,判定兩株菌分別為植物乳桿菌和發酵乳桿菌。
3.2 發酵劑菌種的篩選
(1)發酵時間。在酸乳生產過程中,如果凝乳時間太短,不利于酸乳香味的生成,也會對酸乳的組織狀態產生影響。凝乳時間過長,雖然對狀態和香味有一定作用,但不利于節約生產能源和時間,也易造成發酵過程中的雜菌污染[13]。由表4可以看出,大部分乳酸菌的凝乳時間在17 h以下,菌株BST4-1發酵時間為32 h,發酵時間過長,不滿足發酵需求。
(2)滴定酸度。由表4可以看出,8株乳酸菌的滴定酸度均在60~90°T,GB19302-2010規定酸乳酸度應大于70°T[14],但是由于后期要進行菌株復合發酵,故酸度滿足生產要求。
(3)感官評價。由8株乳酸菌發酵制成的酸乳色澤較為良好,均呈現乳白色或微黃色,其中菌株BSTS6-4的色澤最好;菌株BSTS6-1,BSTS6-3,BSTS6-4,XKN 1-5均具有酸牛奶特有的風味且無苦澀味;菌株BLK7-12,BLK7-10,BST2-3雖然有酸牛奶特有的風味但由略微的澀味;菌株BSTS6-1,BSTS6-3,BSTS6-4發酵制得的酸乳質地細膩、表面光滑、無裂縫;菌株BLK7-10,BST 2-3,XKN 1-5發酵制得的酸乳有輕微的乳清析出。菌株BLK7-12制得的酸乳有乳清析出且分層,凝乳狀態差,不滿足需求。
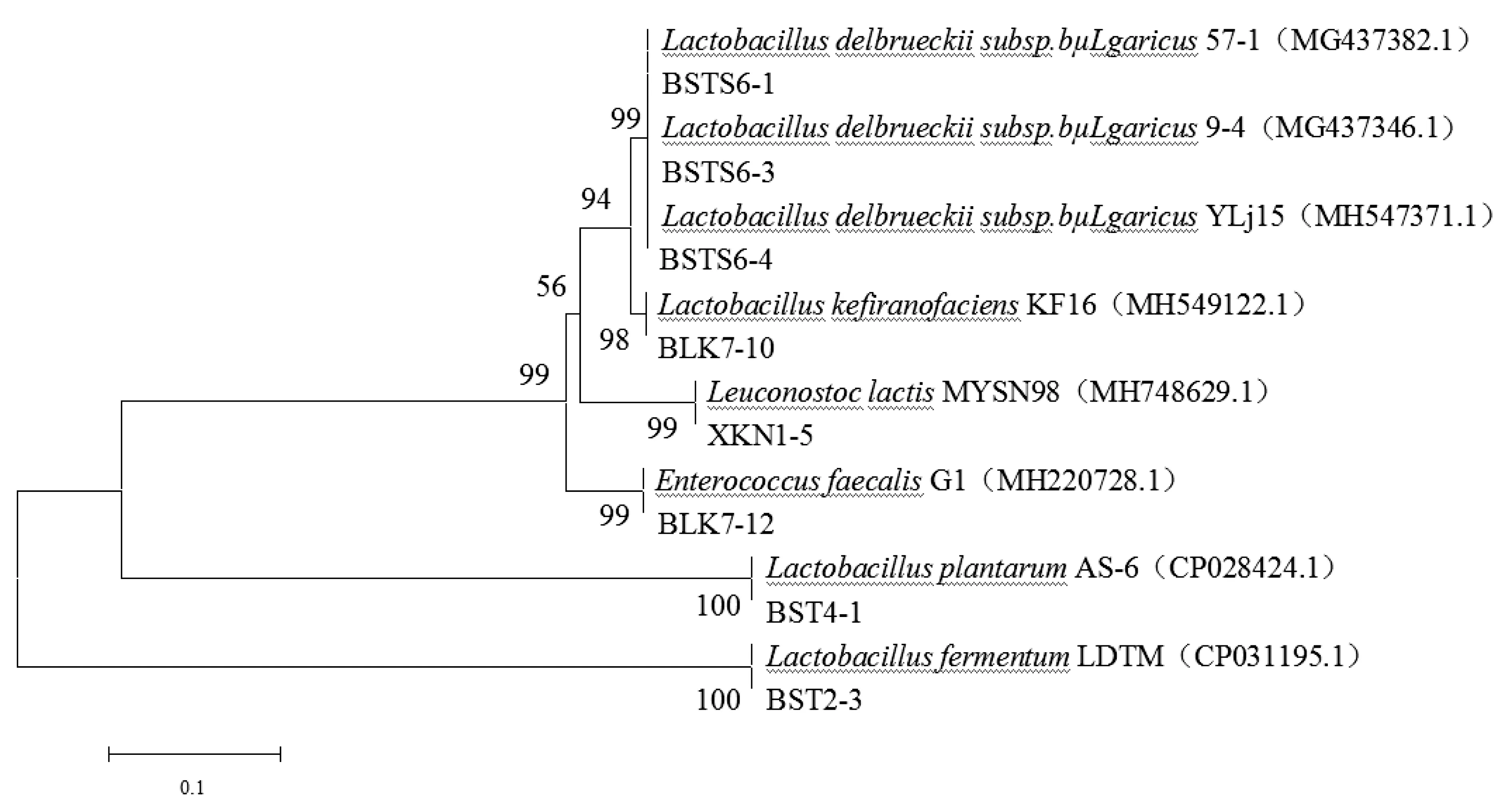
圖3 菌株16SrRNA基因序列的系統發育樹

表3 菌株系統發育相似性分析
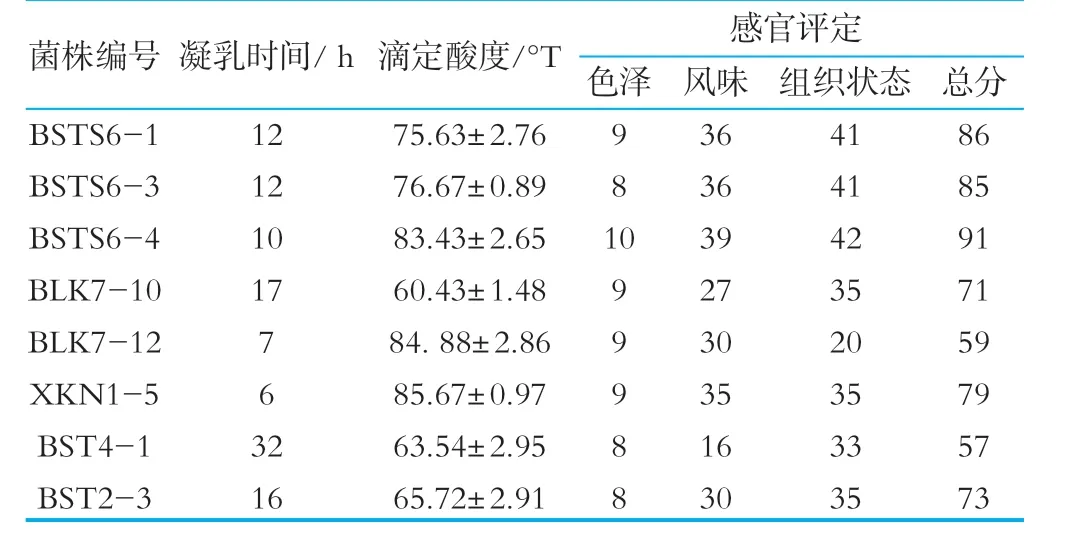
表4 發酵菌種篩選結果
3.3 菌株產酸性能研究
3.3.1 菌株生長曲線
由圖4可以看出,大部分菌株在培養4 h后進入對數生長期,18 h左右進入穩定期。菌株BLK7-10培養至24 h后才有緩慢生長,生長周期過長,不符合工業生產需求。除菌株BLK7-10外,其他5株乳酸菌在培養18 h后菌懸液OD 600值均能達到1.3以上,生長性能 良 好,菌 株 BSTS6-1,BSTS6-3,BSTS6-4,XKN 1-5,BST2-3的最佳收獲時間為18~20 h。
3.3.2 菌株產酸能力
菌株產酸速率是評價酸乳發酵劑的一項重要指標,快速的酸化不僅對成品的香氣、質構和感官特性有重要影響,也能有效防止酸乳污染[15]。由圖5可以看出,菌株XKN 1-5產酸速率最快,20 h內酸度可達到112.86°T,菌株BSTS6-4在20 h內酸度由24.44°T上升到99.48°T,產酸速率較快。菌株BSTS6-1在20 h內,酸度由27.29°T上升到94.31°T,菌株BSTS6-3在20 h酸度由28.51°T上升至97.65°T。菌株BST2-3和BLK7-10產酸速率較慢,實際生產中易造成酸乳污染,產生不必要的經濟損失。
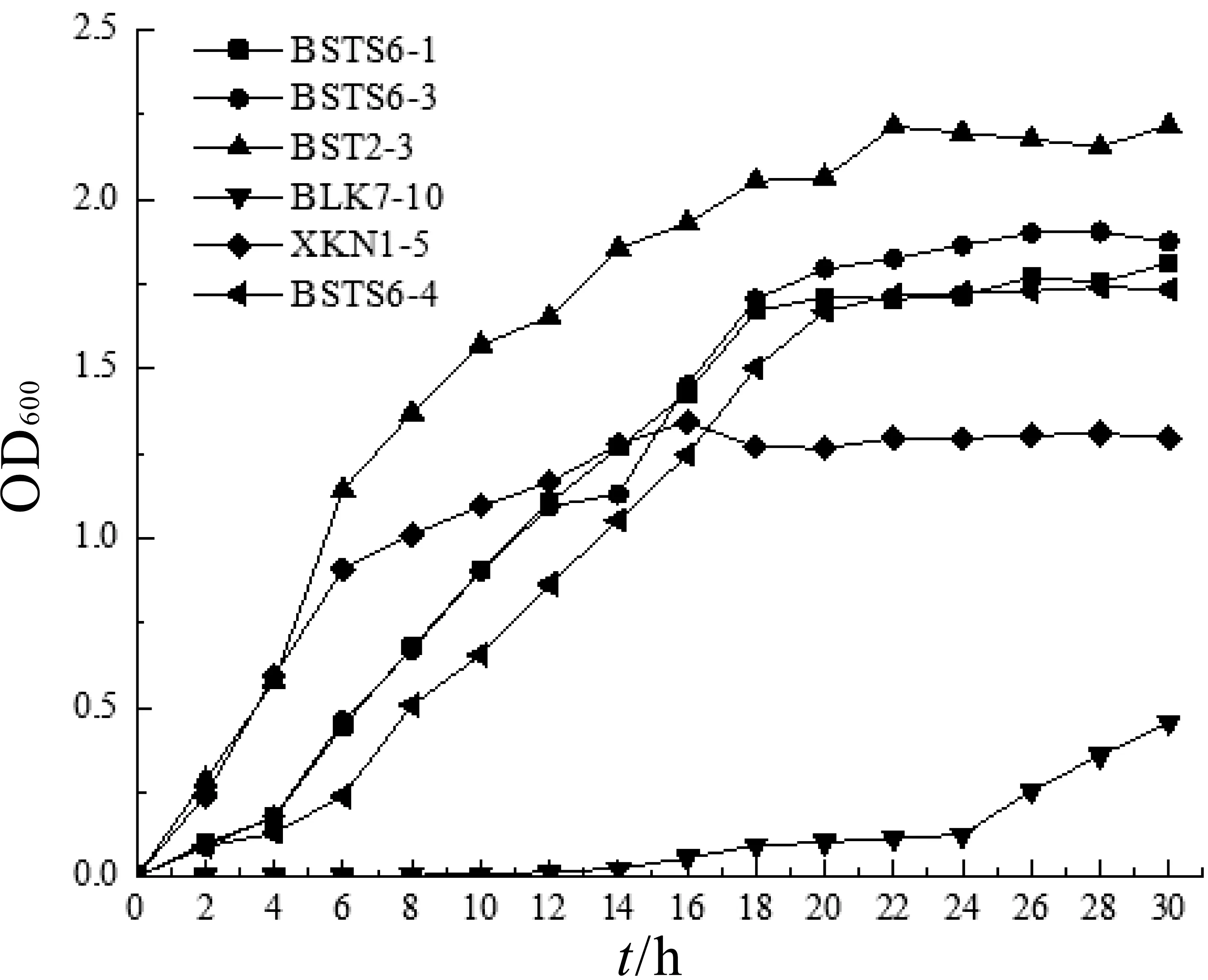
圖4 菌體密度變化曲線(OD600值)
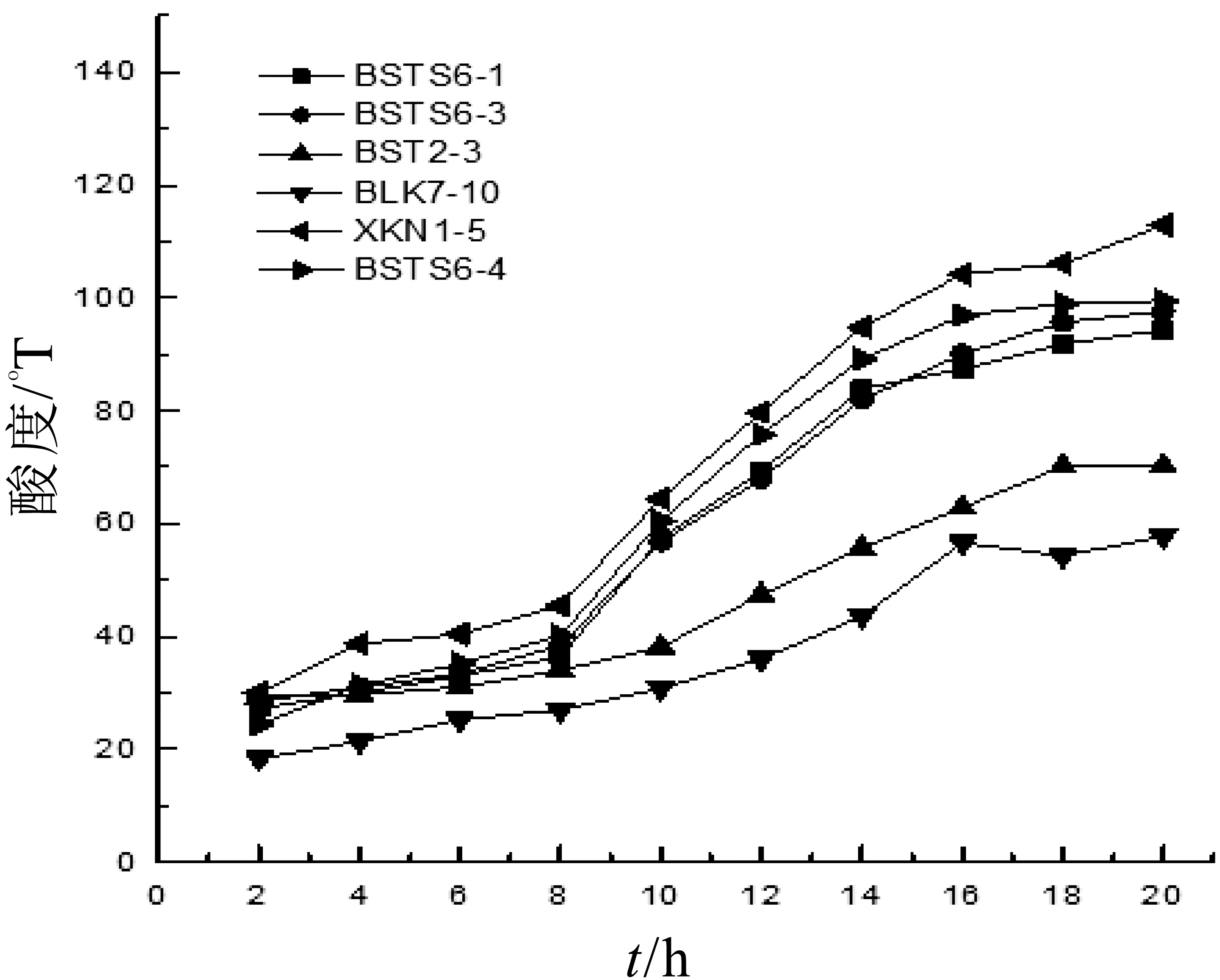
圖5 發酵過程中乳酸菌酸度的變化
3.3.3 菌株后酸化能力
后酸化使得酸乳在冷藏過程中過酸甚至產生苦味,不僅會產生讓消費者難以接受的風味,也會降低產品的貨架期。酸乳生產中常使用后酸化能力較弱的菌株來降低后酸化對酸乳的影響[16]。由圖6可以看出,8株乳酸菌在20 d冷藏過程中酸度均有所升高,其中菌株XKN 1-5酸度變化最大,冷藏過程中酸度增加了24.18°T,后酸化能力較強,可能會造成酸乳風味的改變以及貨架期的降低;菌株BLK7-10在冷藏過程中酸度增加了3.9°T,酸度變化最小,這可能與其自身產酸速率較低有關;菌株BSTS6-1,BSTS6-3,BSTS6-4,BST 2-3在冷藏過程中酸度增加量較為適度,不會對酸乳的品質造成影響。
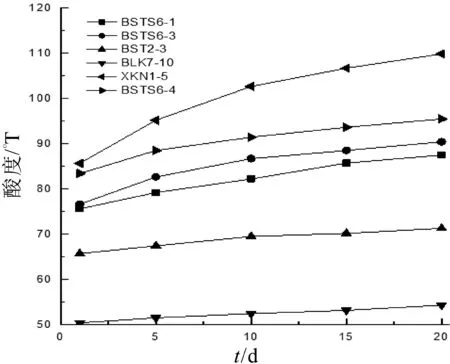
圖6 冷藏過程中酸乳酸度的變化
4 結論
從新疆伊犁地區采集的乳品中共分離出8株具有產酸凝乳性能的乳酸菌,其中菌株BSTS6-4,BSTS6-3,BSTS6-1為德氏乳桿菌保加利亞亞種(Lactobacillus delbrueckii subsp.b Lugaricus),菌株BLK7-10為開 菲 爾 乳 桿 菌(Lactobacillus kefiranofaciens),菌 株XKN 1-5為乳酸明串球菌(Leuconostoc lactis),菌株BLK7-12為 糞 腸 球 菌(Enterococcus faecalis),菌 株BST 2-3為發酵乳桿菌(Lactobacillus fermentum),菌株BST 4-1為植物乳桿菌(Lactobacillus plantarum)。經過初篩得到6株凝乳時間短,酸乳組織狀態良好、風味較佳的乳酸菌。通過對6株乳酸菌的生長性能、產酸性能進行分析對比,得到4株生長速度快、菌懸液濃度高、產酸速率高的乳酸菌XKN 1-5,BSTS6-1,BSTS6-3,BSTS6-4。后酸化實驗表明,菌株XKN 1-5在冷藏過程中酸度變化較大,可能會造成酸乳在冷藏過程中的酸敗變質,不利于酸乳的后期儲存,不符合工業生產需求。菌株BSTS6-1,BSTS6-3,BSTS6-4后酸化能力較弱,在冷藏過程中酸度變化較小,因此菌株BSTS6-1,BSTS6-3,BSTS6-4將作為備選菌株應用到后期的復合發酵實驗。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
