TALEN介導(dǎo)水稻OsTZF9基因定點(diǎn)突變研究
2019-09-10 07:22:44周淑芬施惠芳
福建農(nóng)業(yè)科技 2019年2期
關(guān)鍵詞:水稻
周淑芬 施惠芳


摘?要:RRTZF鋅指基因在植物生長發(fā)育和脅迫反應(yīng)中起著重要作用。利用TALEN基因組編輯技術(shù)定點(diǎn)突變水稻RRTZF基因OsTZF9,結(jié)果表明:根據(jù)日本晴基因組中的OsTZF9基因序列,設(shè)計(jì)3對(duì)TALEN靶序列,并構(gòu)建相應(yīng)的3個(gè)TALEN植物敲除表達(dá)載體;通過農(nóng)桿菌介導(dǎo)法分別將3個(gè)TALEN敲除載體導(dǎo)入水稻明恢86和秀水134的胚性愈傷組織中,獲得了100個(gè)轉(zhuǎn)化再生植株,PCR檢測(cè)獲得87個(gè)陽性植株;PCR擴(kuò)增轉(zhuǎn)基因植株靶突變區(qū)并測(cè)序列,發(fā)現(xiàn)所有轉(zhuǎn)基因植株均未出現(xiàn)靶突變。
關(guān)鍵詞:水稻;OsTZF9基因;TALEN;定點(diǎn)突變
DOI: 10.13651/j.cnki.fjnykj.2019.02.004
Abstract: RRTZF zinc finger gene plays an important role in plant growth, development and stress response. Target mutation of RRTZF gene OsTZF9 in rice was carried out by TALEN genome editing technique. The results showed that three pairs of TALEN target sequences were designed according to the sequence of OsTZF9 gene in Nipponbare, and three plant knockout expression vectors were constructed and introduced into embryogenic callus of Minghui 86 and Xiushui 134 by Agrobacteriummediated transformation, obtaining 100 transformants. Among them, 87 positive plants were obtained through PCR detection. The target mutation regions of the transgenic plants were detected and sequenced, to find no target mutation.
Key words: Rice; OsTZF9 gene; TALEN; Sitedirected mutation
RRTZF鋅指基因是一類植物特有的CCCH型鋅指基因,因其編碼蛋白含有1個(gè)TZF模體及其上游50個(gè)氨基酸為富含精氨酸區(qū)(RR)而得名,該TZF模體由中間間隔16個(gè)氨基酸的2個(gè)不同CCCH結(jié)構(gòu)域組成[1-2]。通過全基因組學(xué)分析,目前已在多種單雙子葉植物中鑒定了許多RRTZF成員。除了擬南芥中所有RRTZF基因的功能得到一定程度的解析,其他植物中僅有少部分TZF基因功能被研究。
OsTZF9是一個(gè)水稻胚乳中特異性表達(dá)的RRTZF基因,酵母單雜分析發(fā)現(xiàn),OsTZF9蛋白與谷蛋白基因啟動(dòng)子區(qū)結(jié)合,RNAi技術(shù)分析發(fā)現(xiàn)RNAi轉(zhuǎn)基因植株與對(duì)照表型沒有區(qū)別,但部分轉(zhuǎn)基因植株種子中的谷蛋白含量比對(duì)照低,表明OsTZF9基因具有下調(diào)谷蛋白基因表達(dá)的功能[3-5]。在擬南芥的RRTZF基因功能研究中,發(fā)現(xiàn)不同RRTZF基因過表達(dá)植株具有一因多效的作用,且存在許多相似表型[6],暗示植物RRTZF基因具有一定程度的功能冗余。由于這類基因存在著功能冗余,許多RRTZF基因單突變體出現(xiàn)的表型很微小,難以觀察到或者根本就沒有任何表型,因此通過RNAi或反義RNA等擬敲除技術(shù)獲得的擬突變體常因抑制不徹底更難觀察到表型。
基因組編輯是一種在基因組水平上對(duì)DNA序列進(jìn)行定向改造的遺傳操作技術(shù),克服了RNAi技術(shù)抑制不徹底的缺陷。TALEN技術(shù)是2011 年發(fā)展起來的非常有效的基因組編輯技術(shù),由黃單胞桿菌轉(zhuǎn)錄激活類效應(yīng)子TALE和核酸內(nèi)切酶FokⅠ的DNA切割域組成,前者能特異性識(shí)別DNA序列,后者為DNA切割域[7]。本研究構(gòu)建了OsTZF9基因的TALEN敲除載體并獲得其轉(zhuǎn)基因植株,為進(jìn)一步研究OsTZF9基因的功能奠定基礎(chǔ)。
1?材料與方法
1.1?植物材料
轉(zhuǎn)基因受體材料為秈稻明恢86和粳稻秀水134。
1.2?載體與菌株
TALEN左右臂骨架載體pTALENL20和pTALENR16、改造過的pCAMBIA1301載體、大腸桿菌D18由上海斯丹賽公司提供,大腸桿菌DH5a和根癌農(nóng)桿菌LBA4404由福建省農(nóng)業(yè)遺傳工程重點(diǎn)實(shí)驗(yàn)室保存。
1.3?主要試劑
TALEN試劑盒(上海斯丹賽公司),PrimeSTAR HS DNA Polymerase(TaKaRa公司),Taq DNA 聚合酶、質(zhì)粒提取試劑盒、DNA純化試劑盒(天根生化科技有限公司),限制性內(nèi)切酶、DNA Ligation Kit(Fermentas公司),引物合成、DNA測(cè)序(上海博尚生物技術(shù)有限公司),潮霉素(Roche 公司),組織培養(yǎng)試劑(Sigma公司)。
1.4?載體構(gòu)建
根據(jù)TALEN靶序列設(shè)計(jì)原則,在OsTZF9基因的編碼序列區(qū)設(shè)計(jì)TALEN左右臂識(shí)別序列。根據(jù)設(shè)計(jì)的TALEN左右臂識(shí)別序列,按照上海斯丹賽生物技術(shù)有限公司提供的模塊合成方法(FastTALETMTALEN試劑盒),合成TALEN左右臂模塊并分別構(gòu)建于骨架載體pTALENL20和pTALENR16中,經(jīng)酶切和測(cè)序驗(yàn)證后,獲得構(gòu)建正確的TALEN左臂和右臂基因表達(dá)盒。用HindⅢ和AscⅠ酶從質(zhì)粒中切取TALEN左臂基因表達(dá)盒,膠回收5 kb條帶;用SacⅠ和AscⅠ酶從質(zhì)粒中切取TALEN右臂基因表達(dá)盒,膠回收4 kb條帶;用HindⅢ和SacⅠ酶切改造過的pCAMBIA1301植物表達(dá)載體,膠回收8.5 kb條帶。將上述3個(gè)片段一步法連接獲得含有TALEN左右臂基因表達(dá)盒的植物雙元表達(dá)載體,并電擊轉(zhuǎn)化農(nóng)桿菌LBA 4404,于-80℃下保存?zhèn)溆谩?/p>
1.5?農(nóng)桿菌介導(dǎo)的水稻遺傳轉(zhuǎn)化
以日本晴的幼胚誘導(dǎo)愈傷組織,并用于隨后的轉(zhuǎn)化。水稻愈傷組織的誘導(dǎo)、繼代及與農(nóng)桿菌的共培養(yǎng),抗性愈傷組織的篩選及再生等相關(guān)的培養(yǎng)基和具體的試驗(yàn)步驟參照文獻(xiàn)[8]進(jìn)行。
1.6?轉(zhuǎn)基因植株檢測(cè)
CTAB法[9]提取水稻植株葉片基因組DNA,在25 μL體系中分別擴(kuò)增TALEN表達(dá)框內(nèi)的基因片段和OsTZF9靶突變區(qū)。前者引物為FOKF(5′TGACCGAGTTCAAGTTCC 3′)和NOSR(5′TTGCGGGACTCTAATCA3′);后者引物為77GF(5′CAAAGCACATTGCAAACACAG 3′)和77GR(5′GTAACACACCCATCTACACGAAG3′)。PCR反應(yīng)條件為:94℃預(yù)變性5 min;94℃變性45 s,56℃退火45 s (溫度依據(jù)GC含量高低而定),72℃延伸45 s(時(shí)間根據(jù)擴(kuò)增長度而定,1 kb·min-1),共35個(gè)循環(huán);72℃延伸10 min。取PCR擴(kuò)增產(chǎn)物(5 μL)于瓊脂糖凝膠上進(jìn)行電泳并用凝膠成像儀拍照分析。
2?結(jié)果與分析
2.1?設(shè)計(jì)TALEN靶序列
水稻OsTZF9基因包含1個(gè)外顯子,全長843 bp,編碼280個(gè)氨基酸,含有1個(gè)CHCH和1個(gè)TZF模體(圖1)。根據(jù)TALEN左右臂靶序列的設(shè)計(jì)原則,在CHCH和TZF模體區(qū)域共設(shè)計(jì)了3對(duì)TALEN靶序列,如表1所示。TALEN左右臂靶序列的設(shè)計(jì)原則:(1)左右臂的識(shí)別長度為12~19 bp,間隔序列長度為12~21 bp。(2)靶位點(diǎn)鄰5′端堿基是T,3′端以T結(jié)尾(3)靶位點(diǎn)盡量多含A和C,適量T,少量G。(4)盡量避開磷酸化位點(diǎn),如編碼蘇氨酸、絲氨酸、賴氨酸和精氨酸的位點(diǎn)。
2.2?構(gòu)建TALEN左右臂載體
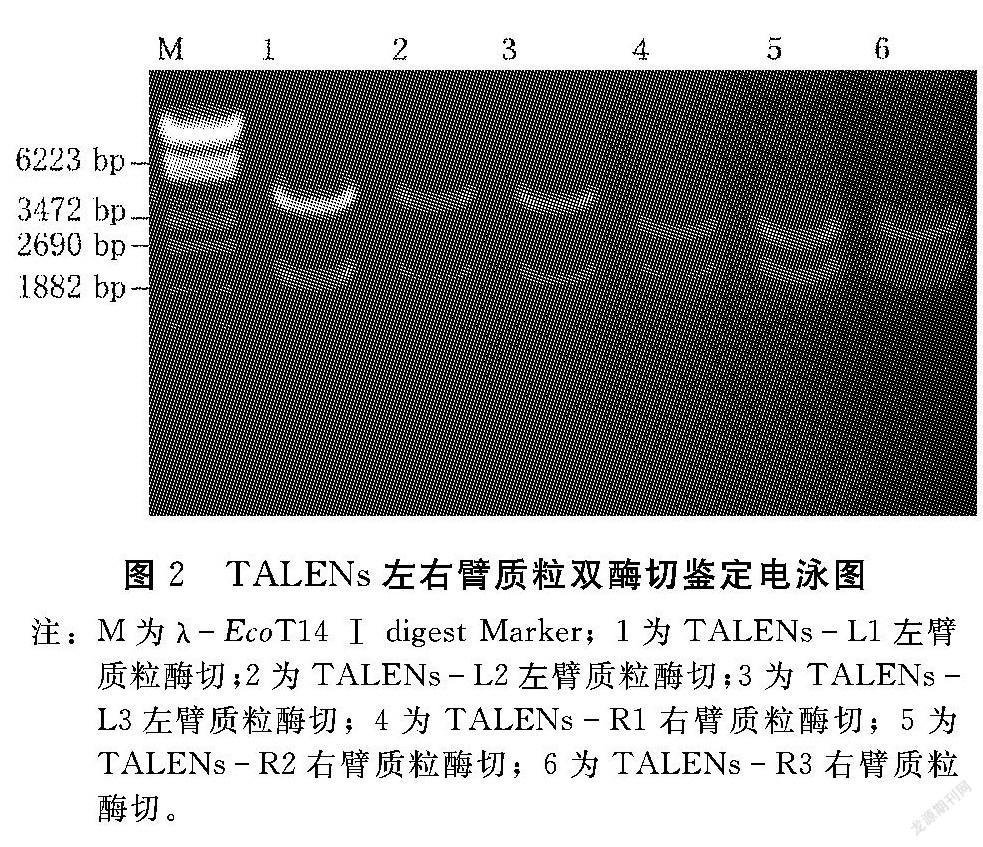
將左右臂識(shí)別序列3′端的最后一個(gè)T去除,其余堿基按1個(gè)或2個(gè)堿基組合依次進(jìn)行標(biāo)記(從1標(biāo)到9),根據(jù)標(biāo)記好的堿基在試劑盒的模塊中選擇對(duì)應(yīng)編號(hào)的模塊與TALEN左臂(pTALENL20)或右臂(pTALENR16)線性骨架載體進(jìn)行9個(gè)片段同時(shí)連接,熱激轉(zhuǎn)化D18感受態(tài)細(xì)胞,挑取單克隆菌落,經(jīng)搖菌提質(zhì)粒后使用Bam H Ⅰ和SpeⅠ酶切獲得重組載體,如圖2所示。進(jìn)一步將上述的重組載體送公司測(cè)序,與設(shè)計(jì)的標(biāo)準(zhǔn)序列在NCBI中進(jìn)行tBlastX比對(duì)。結(jié)果顯示6個(gè)重組載體均100%正確。因此,本研究成功構(gòu)建了6個(gè)正確的TALEN左右臂重組載體。
圖2?TALENs左右臂質(zhì)粒雙酶切鑒定電泳圖注:M為λEcoT14 Ⅰ digest Marker;1為TALENsL1左臂質(zhì)粒酶切;2為TALENsL2左臂質(zhì)粒酶切;3為TALENsL3左臂質(zhì)粒酶切;4為TALENsR1右臂質(zhì)粒酶切;5為TALENsR2右臂質(zhì)粒酶切;6為TALENsR3右臂質(zhì)粒酶切。
用HindⅢ和AscⅠ酶切構(gòu)建正確的TALEN左臂載體,用SacⅠ和AscⅠ酶切構(gòu)建正確的TALEN右臂載體,前者膠回收約5 kb的條帶,后者膠回收約4 K的條帶,用于后續(xù)的載體構(gòu)建實(shí)驗(yàn),圖3為膠回收后電泳檢測(cè)結(jié)果。
2.3?構(gòu)建TALEN植物敲除表達(dá)載體
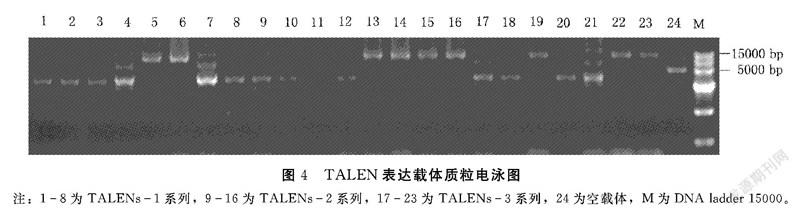
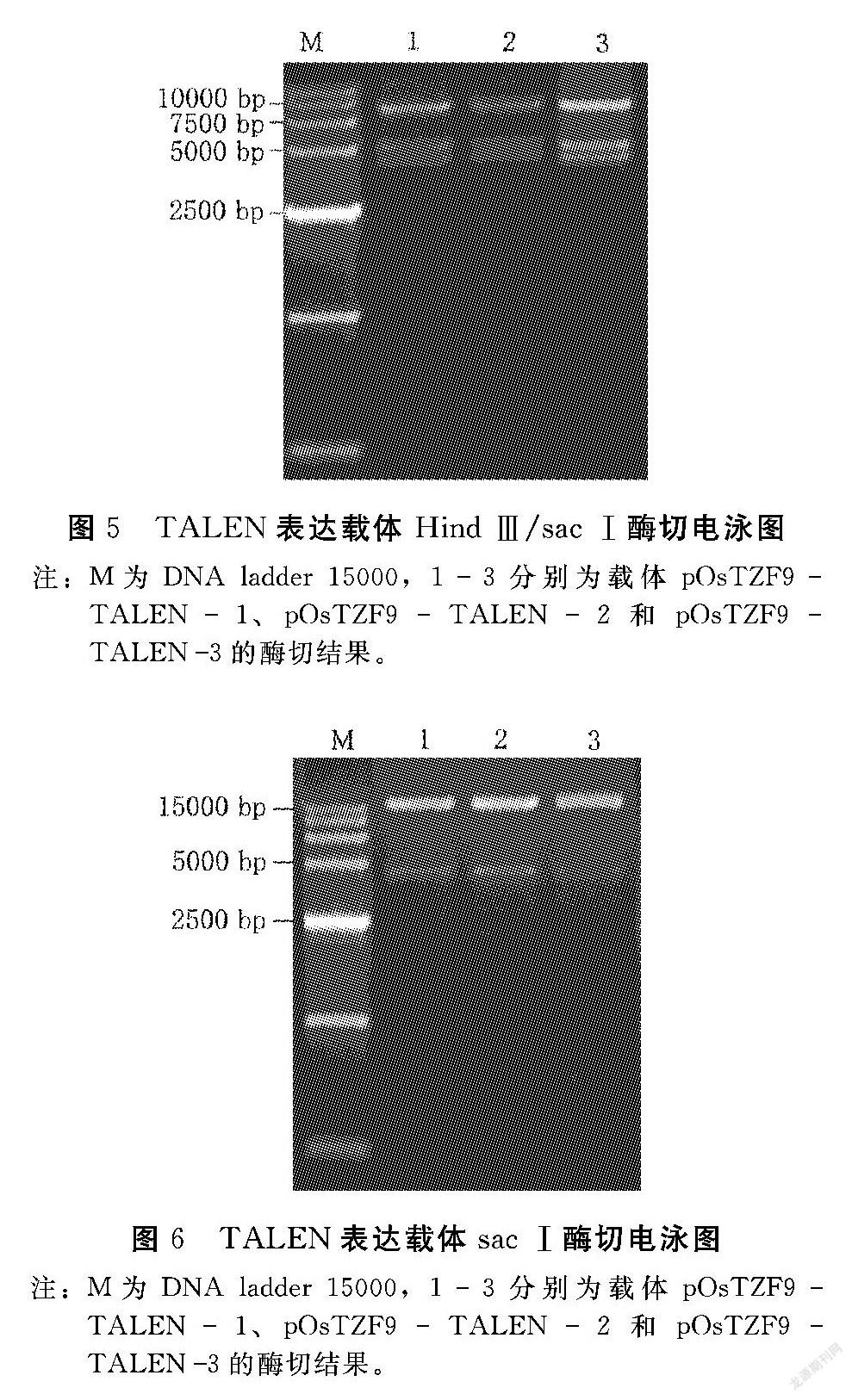
將上述膠回收的左臂(5 kb)、右臂(4 kb)和改造過的p1301線性載體(8.5 kb)于22℃連接3 h,轉(zhuǎn)化感受態(tài)D18。次日挑取單克隆,于37℃、250 r·min-1的搖床中培養(yǎng)16 h左右,提質(zhì)粒跑電泳,比對(duì)照質(zhì)粒大的質(zhì)粒初步確定為連接成功的目的載體(圖4);進(jìn)一步用HindⅢ和SacⅠ雙酶切,出現(xiàn)3個(gè)條帶約為8、5和4 kb(圖5),說明酶切結(jié)果正確;最后用SacⅠ酶切鑒定正確的質(zhì)粒,出現(xiàn)約4 kb和12 kb兩個(gè)條帶(圖6),其中前者為右臂,后者為左臂,分別膠回收這兩個(gè)片段送往公司測(cè)序,測(cè)序結(jié)果均正確,即完成OsTZF9基因3個(gè)靶位點(diǎn)的TALEN敲除載體構(gòu)建,并命名為pOsTZF9TALEN1、pOsTZF9TALEN2和pOsTZF9TALEN3。同時(shí)通過電擊法將上述3個(gè)載體分別導(dǎo)入農(nóng)桿菌中,25%甘油保存于-80℃中。
2.4?水稻遺傳轉(zhuǎn)化及檢測(cè)
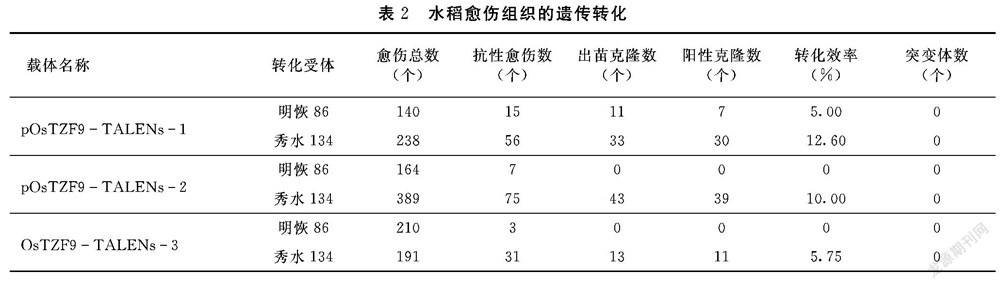
將構(gòu)建好的質(zhì)粒pOsTZF9TALEN1、pOsTZF9TALEN2和pOsTZF9TALEN3分別轉(zhuǎn)化到水稻明恢86和秀水134中,對(duì)獲得的轉(zhuǎn)化再生植株的葉片用浸泡潮霉素法和PCR方法檢測(cè),結(jié)果如表2所示,在明恢86受體材料中僅有pOsTZF9TALEN1質(zhì)粒獲得轉(zhuǎn)基因植株,其轉(zhuǎn)化率為5.00%;在秀水134受體材料中,雖然3個(gè)載體均能獲得轉(zhuǎn)基因再生植株,但其轉(zhuǎn)化率也不高。
PCR擴(kuò)增靶突變區(qū)片段,并測(cè)序分析其靶位點(diǎn)突變情況,結(jié)果發(fā)現(xiàn)所有轉(zhuǎn)基因植株均未產(chǎn)生靶突變。為了排除可能由于TALEN模塊發(fā)生重組導(dǎo)致TALEN活性喪失問題,利用引物305(5′CTCCCCTTCAGCTGGACAC 3′)和306(5′AGCTGGGCCACGATTGAC 3′)對(duì)轉(zhuǎn)基因植物基因組DNA進(jìn)行PCR擴(kuò)增TALEN左右臂序列。由于左右臂大小比較接近,因此對(duì)擴(kuò)增產(chǎn)物TA克隆并轉(zhuǎn)化到大腸桿菌DH5α中,然后對(duì)單菌落進(jìn)行菌液PCR,選擇擴(kuò)增片段2 kb左右菌液進(jìn)行測(cè)序,測(cè)序結(jié)果比對(duì)均正確,說明導(dǎo)入水稻中的TALEN模塊未發(fā)生重組。
3?結(jié)論與討論
本研究成功構(gòu)建了OsTZF9基因3個(gè)靶位點(diǎn)的TALEN敲除載體,通過農(nóng)桿菌介導(dǎo)法導(dǎo)入水稻明恢86和秀水134中,成功獲得87個(gè)轉(zhuǎn)基因植株。但在轉(zhuǎn)化過程中發(fā)現(xiàn),TALEN敲除載體明顯比其他載體的轉(zhuǎn)化率低(本實(shí)驗(yàn)室構(gòu)建的其他基因的TALEN敲除載體也有類似現(xiàn)象),原因之一可能是TALEN植物表達(dá)載體相對(duì)于其他轉(zhuǎn)化載體大,不易轉(zhuǎn)化。
本研究檢測(cè)了所有轉(zhuǎn)基因植株,但均未檢測(cè)到靶突變體。其實(shí),本試驗(yàn)在檢測(cè)其他基因的TALEN轉(zhuǎn)基因植株靶突變情況時(shí),也發(fā)現(xiàn)其突變率很低。其原因可能有3個(gè):一是TALEN載體存在著高度重復(fù),易發(fā)生同源重組, 進(jìn)而影響其活性;二是TALEN表達(dá)框插入的隨機(jī)性,當(dāng)插入到某些重要的基因內(nèi)部, 使TALEN表達(dá)框出現(xiàn)基因沉默或者表達(dá)受到抑制;三是TALEN表達(dá)框插入到植物體中有表達(dá),但是植物體把它當(dāng)成是外源物,觸發(fā)植物體的自我免疫功能,從而降低了TALEN蛋白的活性,因此就無法識(shí)別切割目的基因,導(dǎo)致無法產(chǎn)生突變體。本研究檢測(cè)3個(gè)TALEN模塊的同源重組情況,均未發(fā)生同源重組,其具體原因有待進(jìn)一步研究。
因此,為了能獲得靶基因突變體,建議多設(shè)計(jì)幾個(gè)靶位點(diǎn);同時(shí)為了減少后期的工作量,可以先在原生質(zhì)體中檢測(cè)TALEN蛋白的活性,再轉(zhuǎn)化愈傷組織。由于TALEN蛋白在受體細(xì)胞中可持續(xù)起作用,通過對(duì)抗性愈傷繼續(xù)進(jìn)行繼代培養(yǎng)可產(chǎn)生新突變,增加突變的類型。
參考文獻(xiàn):
[1]POMERANZ MC,HAH C,LIN PC,et al.The Arabidopsis tandem zinc finger protein AtTZF1 traffics between the nucleus and cytoplasmic foci and binds both DNA and RNA[J].Plant Physiol,2010,152:151-165.
[2]Wang D,Guo Y,Wu C,et al.Genomewide analysis of CCCH zinc finger family in Arabidopsis and rice[J].BMC Genomics,2008,9:44-54.
[3]劉梅芳,周淑芬,徐桂磊,等.一個(gè)水稻CCCH型鋅指蛋白基因的表達(dá)模式分析[J].福建農(nóng)業(yè)學(xué)報(bào),2008,23(3):225-230.
[4]周淑芬,劉華清,徐桂磊,等.水稻種子發(fā)育期間特異性鋅指蛋白基因的篩選與分析[J].中國生物化學(xué)與分子生物學(xué)報(bào),2012,9(2):237-289.
[5]CHEN Y,SUN A,WANG M,et al.Functions of the CCCH type zinc finger protein OsGZF1 in regulation of the seed storage protein GluB?1from rice[J].Plant Mol Biol,2014,84:621-634.
[6]POMERANZ M,F(xiàn)INER J,JANG JC.Putative molecular mechanisms underlying tandem CCCH zinc finger protein mediated plant growth,stress and gene expression responses[J].Plant Signaling & Behavior,2011,6(5):647-651.
[7]MILLER JC,TAN S,QIAO G,et al.A TALE nuclease architecture for efficient genome editing[J].Nat Biotechnol,2011,29:143-148.
[8]蘇軍,胡昌泉,翟紅利,等.農(nóng)桿菌介導(dǎo)秈稻明恢86高效穩(wěn)定轉(zhuǎn)化體系的建立[J].福建農(nóng)業(yè)學(xué)報(bào),2003,18(4):209-213.
[9]MURRY MG,THOMPSON WF.Rapid isolation of high molecular weight plant DNA[J].Nucl Acid Res,1980,8(19):4321-4326.
(責(zé)任編輯:柯文輝)
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學(xué)版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農(nóng)業(yè)(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
