三峽庫區消落帶立柳(Salix matsudana)生長及營養元素分配特征
2019-09-05 04:50:56吳科君馬文超陳紅純何欣芮
生態學報 2019年14期
吳科君,馬文超,李 瑞,陳紅純,黃 超,何欣芮,魏 虹,*
1 三峽庫區生態環境教育部重點實驗室,重慶市三峽庫區植物生態與資源重點實驗室,西南大學生命科學學院, 重慶 400715 2 攀枝花市旅游局, 攀枝花 617000
三峽水庫自建成運行后,實行“冬蓄夏排”的反季節性水位調度模式,在每年的10月底到達最高水位175 m,次年6至9月維持在最低水位145 m[1],形成了庫岸線長2000 km、消漲幅度為30 m、面積約350 km2的消落帶[2]。庫區消落帶內大量原生植物因不能適應反季節長時間水淹而大面積消亡,造成消落帶生物多樣性降低,生態屏障功能減退以及景觀惡化等環境問題[3]。為恢復庫區消落帶生態系統正常的結構和功能,利用適生物種進行植被重建被認為是最有效的手段之一[4- 6]。近年來,研究者逐漸將前期模擬研究篩選出來的耐水淹物種用于消落帶植被重建示范工作中。立柳(Salixmatsudana)作為庫區本土植物,因其耐水淹、生長迅速和成活率高等特性被用于庫區165 m以上消落帶植被重建[7- 8]。
前期模擬實驗研究發現,在水淹脅迫下立柳可通過改變形態特征、光合效率以及提高抗氧化防御系統中酶的活性和增強次生代謝等來提升對水淹環境的耐受能力[7- 10]。隨著庫區消落帶植被重建與恢復工作的展開,基于原位的試驗研究逐漸增多,研究者通過對植物的生長情況、分配位置以及生理生態機制等方面的研究[11- 13],豐富了立柳等重建物種對庫區“冬蓄夏排”的水文節律的響應機制。已有研究表明植物體內各器官中的營養元素對植物生長和各種生理功能起著重要的作用[14]。營養元素的不足會限制植物生長、發育和代謝,進而影響庫區消落帶植被修復與重建的可持續性[10,15- 16]。而高強度的水淹會顯著影響植物各器官中營養元素的積累與分配方式,并影響到植株的生長[17- 21]。因此,通過原位試驗探究在三峽庫區消落帶立柳的營養元素積累特征,對了解該物種對庫區水位變化的適應及可持續性生長是必要的。
2012年3月,本課題組在位于重慶忠縣石寶鎮共和村汝溪河流域的三峽庫區消落帶植被修復示范基地165—175 m海拔區域內種植生長均勻的2年生立柳植株。經過3個水淹周期后,植株生長良好,為進一步探討立柳對庫區消落帶冬季水淹的適應機理,于2015年5月中旬展開實地采樣與研究,分析在經歷了冬季水淹后的退水初期立柳各器官中營養元素的積累和分配特征,以期為消落帶植被重建工作提供理論和實踐指導。
1 材料與方法
1.1 研究材料
立柳(S.matsudana)是三峽庫區原有的多年生庫岸樹種,又稱龍爪柳,是旱柳的一個變種,屬楊柳科,柳屬落葉喬木。本研究選擇三峽庫區汝溪河流域消落帶植被修復示范基地(107°32′—108°14′E,30°03′—30°35′N)作為試驗樣地,其中在基地內海拔為165—175 m之間以1 m×1 m的密度種植了立柳純林,該林地面積約4 hm2。2012年3月種植時平均株高為1.613±0.13 m,基徑0.288±0.045 cm,冠幅0.492±0.034 m2。試驗樣地3個采樣帶中土壤元素含量見表1。
表中數值為平均值±標準誤(n=10);同列不同小寫字母分別表示處理之間有顯著差異(P<0.05)
1.2 試驗設計及取樣
于2015年5月中旬進行實地取樣,取樣時整個立柳純林中林木保存率約為95%。根據每個淹水周期水淹時長和深度的差異將示范基地劃分為3個采樣帶:175 m的對照組(CK)、170 m的中度水淹組(MS)以及165 m的深度水淹組(DS)。各采樣帶3個水淹周期內水淹情況見表2。
為盡可能減小陽光照射等因素對立柳的影響,分別在3個樣帶內隨機選取長勢均一的立柳樣木,使用高枝剪于樹冠中上層的東、南、西、北四個方位隨機采集葉和枝條,然后將各方位的葉片、枝條單獨混合后裝入自封袋;用根鉆以植株基部為圓心,0.5 m半徑等距離鉆取植株根樣(直徑2—5 mm),混合裝于自封袋,每個采樣帶設置5個重復。樣品帶回實驗室后,先用超純水將其清洗干凈,然后置于105℃的烘箱內殺青5分鐘,再置于60℃的烘箱中烘干至恒重,最后將烘干后的樣品放入球磨儀中粉碎后,封裝待測。
在采樣的同時,在3個樣帶隨機選取立柳各10株,原位記錄立柳的株高、基徑和冠幅生長指標。使用卷尺測定每株立柳的株高和冠輻,用數顯游標卡尺測量基徑。
1.3 營養元素含量測定
將待測的植物樣品粉碎、過篩后,稱取0.005 g,采用Vario EL cube CHNOS元素分析儀(Elementar,德國)進行全N含量測定;另外稱取0.05 g使用SpeedWaveMWS- 4微波消解儀(Berghof,德國)消解待測樣后,采用ICAP 6000電感耦合等離子體發射光譜儀(Thermo,美國)測定立柳根、枝條及葉片P、K、Ca、Mg、Fe、Mn、Cu、Zn含量。
1.4 數據分析
使用統計分析軟件SPSS 22.0進行數據處理以及統計分析,用配對樣本t檢驗的方法(Paired sample T test)比較各處理在種植初期和種植3 a后的生長差異;采用單因素方差分析(One-way ANOVA)檢驗水淹處理對立柳生長和營養元素含量的影響,用Turkey法進行多重比較。使用Origin 8.5軟件制圖。
2 結果與分析
2.1 生長狀況
圖1所示為庫區消落帶示范基地內不同處理組立柳的生長狀況。經過3 a的生長,與種植初期相比,3個采樣帶的立柳株高、基徑和冠幅均有極顯著增加(P<0.01)。隨著海拔位的降低,水淹時長和深度增加,立柳的生長受到明顯抑制。經過3個水淹周期后,與CK組相比,MS組和DS組植株株高分別降低了18.4%和48.7%,DS組立柳的基徑和冠幅分別降低了50.3%和71.5%,但MS組的基徑和冠幅沒有顯著差異。
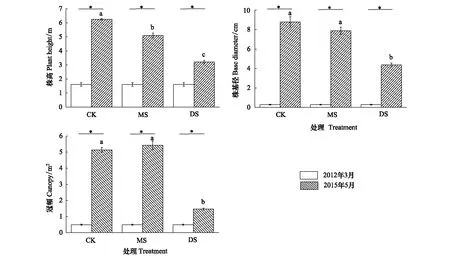
圖1 立柳植株生長狀況Fig.1 The growth situations of S. matsudana圖中數值為平均值±標準誤 (n=5);3個采樣帶中的立柳種植時間和各初始生長指標一致: *表示各處理組在2個時期有極顯著差異(P<0.01),相同字母表示2015年5月不同處理組間沒有顯著差異(P>0.05),不同小寫字母表示各處理組之間有顯著差異 (P<0.05)
2.2 水淹脅迫對立柳各器官中N、P、K含量的影響
N、P、K是植物生長發育所需要的大量元素。庫區3個采樣帶的立柳對N、P、K元素含量在植株根、枝、葉中的分配表現出了類似的特征。從總體上來看,這3種大量元素在立柳葉片中的含量均顯著高于在根和枝條中的含量(圖2)。
在經歷了不同強度的冬季水淹后,3個采樣帶內立柳的根、枝條和葉片中N、P、K元素的含量變化表現出不同的特征。水位變化對立柳葉和枝條中N、P元素含量無顯著影響,但顯著降低了水淹組根中N、P元素含量(P<0.05)。MS組和DS組N元素含量較CK組分別降低了28.2%和26.6%,P元素則分別降低了37.5%和15.2%。水位變化未對立柳葉和根中的K元素含量造成顯著影響,但DS組枝條中鉀含量顯著低于MS組和CK組(圖2)
不同處理組的立柳葉、枝條及根中N、P、K含量分別為5.2—23.7 g/kg、0.9—4.4 g/kg和5.3—13.6 g/kg,即為各器官干質量的0.52%—2.37%、0.09%—0.44%以及0.53%—1.37%,這些值均處于植物含量正常水平[22]。
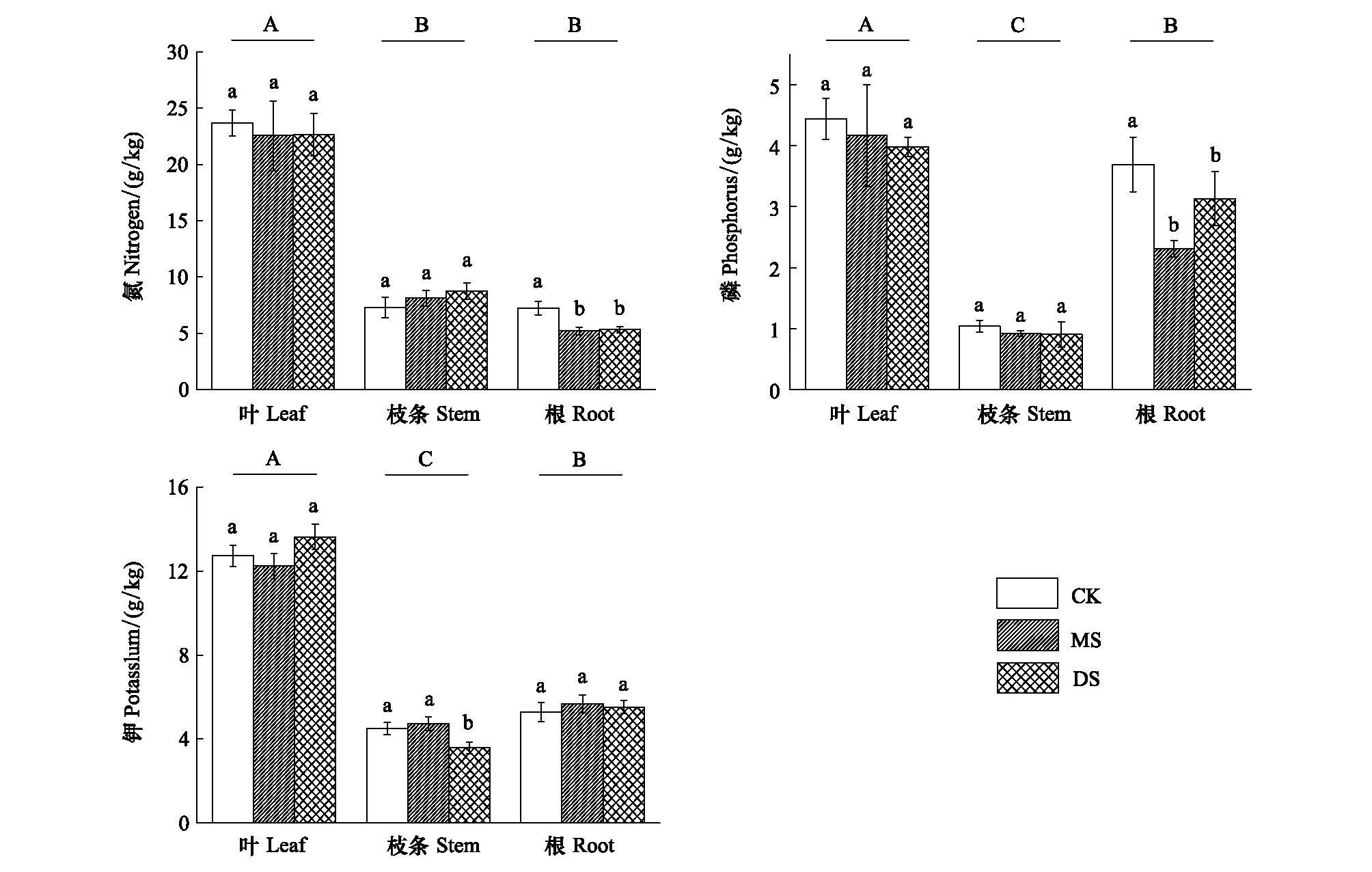
圖2 不同水淹處理下立柳N、P、K元素在葉、枝條及根中的含量變化Fig.2 Changes of N, P and K contents in leaves, stems and roots of S. matsudana under different flooding treatments圖中數值為平均值±標準誤 (n=5);不同小寫字母表示同一組織各處理之間有顯著差異 (P<0.05);不同大寫字母表示各器官間有顯著差異(P<0.05)
2.3 水淹脅迫對立柳各器官中Ca、Mg含量的影響
Ca和Mg元素是植物體內重要的中量元素。從各器官中元素分配的總體情況來看,Ca元素表現為根中的含量明顯低于葉和枝條,而Mg在各組織中基本均勻分布。
從圖3可以看出,冬季水淹對3個采樣帶中立柳植株的Ca、Mg元素含量影響顯著。在立柳的葉和枝條中,DS組中Ca元素含量受到水淹脅迫而顯著降低(P<0.05),而CK組與MS組之間無顯著差異;在立柳的根中則表現為MS組含量顯著高于CK組和DS組,水淹抑制了DS組立柳葉片對Mg的吸收和積累,促進了立柳枝條對Mg的積累,而抑制了立柳根對Mg的吸收和積累各器官不同處理組中鈣、鎂的含量分別為7.9—31.3 g/kg和1.1—2.1 g/kg,仍處于植物正常元素含量范圍[22]。
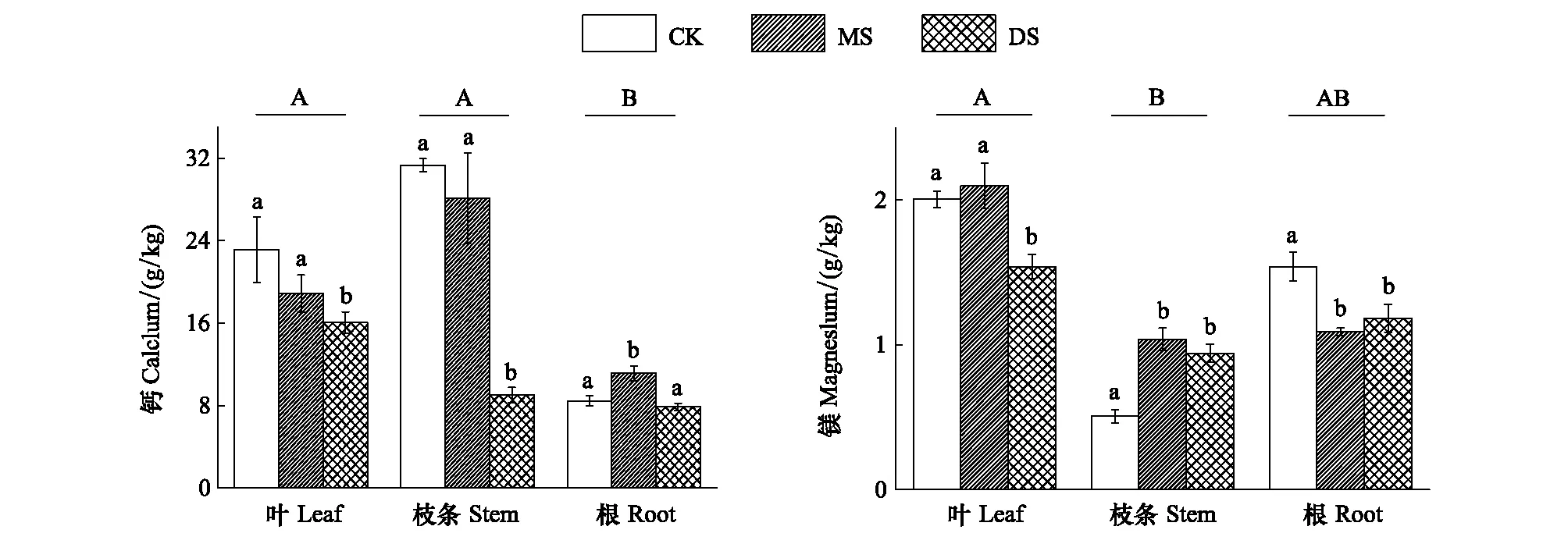
圖3 不同水淹處理下立柳Ca、Mg元素在葉、枝條及根中的含量變化Fig.3 Changes of Ca and Mg contents in leaves, stems and roots of S. matsudana under different flooding treatments
2.4 水淹脅迫對立柳各器官中Fe、Mn、Cu、Zn含量的影響
Fe、Mn、Cu、Zn是植物體內重要的微量元素,其含量變化會直接影響植株的生理功能。這4種微量元素在立柳植株各器官中受水淹脅迫影響而表現出不同的分配特征。由圖4可知,Fe元素在立柳葉、枝條和根中總體的表現為大量分配到立柳的根中;Mn元素分配情況總體表現為葉>根>枝條;Cu和Zn元素則主要分布在立柳枝條中。各處理組不同器官中營養元素Fe、Mn、Cu、Zn的含量均未低于植物體內的正常水平[22]。
由圖4可知,水淹脅迫顯著增加了根系中Fe元素(P<0.05),深度水淹(DS處理組)提高了立柳枝條中Fe元素含量,但水淹脅迫對葉片Fe含量的影響不顯著。此外受水淹影響,MS組和DS組立柳葉中Mn含量較對照組顯著降低(P<0.05),而在枝條和根中,水淹組含量均高于對照組。水淹處理極顯著地降低了 DS組枝條中Cu和Zn的含量,同時降低了MS組和DS組根中的元素含量。
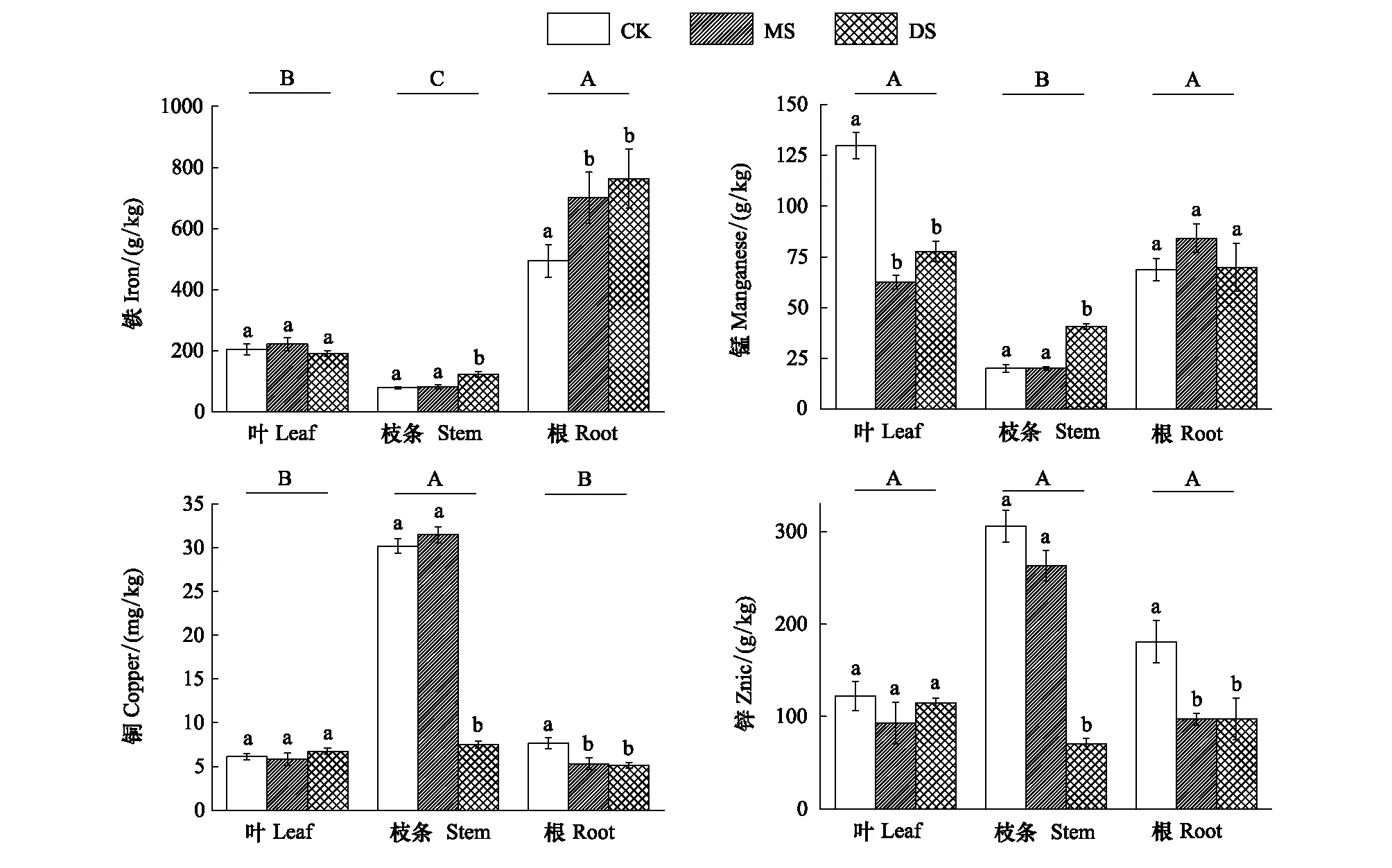
圖4 不同水淹處理下立柳Fe、Mn、Cu和Zn元素在葉、枝條及根中的含量變化Fig.4 Changes of Fe、Mn、Cu and Zn contents in leaves, steams and roots of S. matsudana under different flooding treatments
3 討論
在經歷了3個水淹周期之后,各海拔高程的立柳生長狀況和營養元素含量表現出顯著的差異性。與種植初期相比,立柳株高、基徑和冠幅均有顯著增加,表明該物種對三峽庫區消落帶冬季水淹具有良好的適應性,并在退水后具有較好的恢復生長能力。良好的生長狀態有利于植株爭取更多的生長空間和資源,從而更好地應對三峽庫區消落帶周期性水淹[23]。但立柳的株高、基徑和冠幅等生長指標均隨著海拔高度的降低即水淹脅迫強度的增加而逐漸降低。與175 m海拔(無水淹脅迫)和170 m海拔(中度水淹脅迫)區域的植株相比,經過3 a的冬季水淹后,165 m海拔(深度水淹脅迫)處立柳的生長受到了明顯抑制。生長于該海拔處的植株冬季最大水淹深度為10 m,水淹時間超過5個月,高強度的水淹導致植物光合產物的消耗明顯增加,退水后恢復生長的難度也加大,植株光合產物的凈積累量減少[24- 26]。
耐水淹植物能夠通過生理生態的變化來適應淹水脅迫,其中通過改變營養元素積累與分配來適應水淹是重要的手段之一[10]。水淹導致植物因根系缺氧而引起根際功能紊亂甚至死亡,同時還會影響土壤中營養元素的含量以及有效性,最終顯著影響植物根系對營養元素的吸收利用[27]。本研究結果顯示,庫區消落帶水位周期性的變化導致水淹組立柳根中的N、P元素含量顯著低于對照組,但各處理組葉和枝條中氮、磷元素的含量無顯著差異。植物葉中N、P元素的含量在一定范圍內與植物的光合速率成正比[28- 29],立柳葉片中較高的N、P元素含量,保證了植株正常高效的光合效率,從而保證了水淹組植株正常生長存活。K是維持類囊體膜上pH梯度的平衡離子[30],也是多種光合酶的活化劑,同時還參與了光合作用的CO2同化[31]; K還有助于植物組織的擴增生長,有利于植株生物量的積累。在本研究中,除165 m海拔處的立柳枝條中K元素含量有降低外,水淹未對立柳葉、枝條和根中K元素含量造成顯著影響,說明庫區內立柳在水淹脅迫條件下對該元素的積累和分配具有較好的適應,從而保證了植株株高、基徑、冠幅等組織的擴增及正常的生理功能。
Ca和Mg元素是植物體內重要的中量元素。Ca能夠感知和調節環境壓力[14],通過維持細胞壁、細胞膜及膜蛋白的穩定性,以及通過參與信號傳導來調節植物細胞對逆境反應和逆境適應性[32]。Mg是葉綠素的重要組成部分,在葉片中積累的最多,主要參與植物的光合作用[14],并涉及到大量光合酶的活化以及植物韌皮部形成[33]。本研究結果表明,165 m海拔處立柳葉和枝條中的Ca含量顯著降低,但根中Ca元素的含量相對穩定。說明在庫區消落帶高強度的水淹脅迫降低了植株自身機能,導致對Ca離子的吸收能力下降,地上部分的積累量減少。立柳葉和枝條中Ca元素含量的降低可影響葉綠體雙磷脂膜的形成,導致有機質積累量的持續減少,并影響到植物的正常生長,這也許是造成庫區165 m海拔處立柳各項生長指標顯著降低的原因之一。但立柳根中Ca元素含量的穩定,有助于對逆境脅迫信號的傳遞及并積極維持細胞膜形態,防止厭氧呼吸產生的有害物質對細胞膜的破壞。各處理組立柳Mg元素的含量和分配特征均表現為葉>根>枝條,但受到水淹影響,海拔165 m的立柳葉中的含量顯著降低,這可能會影響到植物的正常光合作用,導致立柳生長所需的光合產物供應不足。因此165 m海拔的立柳需要追蹤關注Mg含量降低對其生長造成的影響。
微量元素在植物體內的含量非常少,但對植物的生長卻至關重要[34]。Fe和Mn是光合作用功能單元中的重要組分。本研究結果顯示,水淹不同程度增加了立柳根部Fe和Mn的含量,但對立柳葉中的Fe和Mn的含量影響不明顯,均處于正常范圍內。立柳葉中Fe和Mn的含量的穩定是維持其正常的光合作用而采取的一種自我保護機制。但在水淹環境下,土壤氧化還原電位降低,Fe2+和Mn2+濃度增加,可溶性Fe2+和Mn2+很容易被植物根部過量吸收,導致立柳體內Fe、Mn的含量顯著增加[35],尤其是165 m海拔處立柳的根和枝條。植物過量吸收Fe元素會對自身產生毒害作用,不僅會影響植物的光合作用,還會對植物呼吸酶結構造成破壞;同時立柳根部Fe元素的大量積累會在根際形成一層鐵氧化合物保護膜[36],該保護膜會進一步限制立柳對其他營養元素的吸收利用,最終影響到植物的伸長生長,這可能是導致海拔165 m立柳各生長指標較170 m以上顯著降低的又一個原因。
Cu和Zn是氧化還原酶的成分,在植物體內主要參與氧化還原反應。植物在水淹脅迫下會增加根部對Cu和Zn的吸收量,從而保證植物根際的強氧化還原能力,將低價陽離子氧化、陰離子還原,防止長期水淹導致低價陽離子產生的毒害[22]。本研究結果顯示,水淹導致立柳根中Cu和Zn元素含量顯著降低。立柳根中Cu和Zn元素含量降低會導致立柳Fe、Mn元素含量顯著增加,而Fe元素含量增加又會限制植物對Cu和Zn的吸收。長期以往可能會影響到立柳正常的生理功能。本研究結果還表明,165 m海拔的立柳枝條中Cu、Zn含量顯著低于170 m以上的立柳,這會導致165 m海拔處立柳的呼吸作用下降,同時由于生長激素的合成不足而影響植物的伸長生長[37],最終引起海拔165 m的立柳生長狀況相交于其他兩組差一些。
除此之外,庫區冬季高強度水淹還會導致植物的生境發生改變,如土壤的酸堿度、水氣交換情況、土壤微生物群和土壤元素含量等均會對植物的生長以及營養元素的吸收造成影響[11,38]。同時相關研究表明,在水淹脅迫下,植物根系代謝受阻會使根系對土壤中營養元素的主動吸收減弱,從而導致土壤中營養元素的可利用性與植物營養元素含量之間的關系變弱[39- 40]。目前一些學者對庫區消落帶內的立柳植株各營養元素與土壤中相對應的營養元素進行相關性分析后,發現這兩者之間并無顯著性相關關系[41],這說明了在水淹環境下,造成庫區消落帶內不同海拔高度的立柳生長及營養元素積累量之間存在差異更主要是由水淹脅迫導致的。
綜上所述,在本研究中三個不同水淹強度下的立柳能主動通過調整其營養元素的積累和分配方式積極應對庫區消落帶周期性的水位變化。經過3個水淹周期后,165 m海拔處立柳營養元素的積累和分配方式與170 m海拔以上區域植株的差異較為明顯,特別是Ca、Mg、Fe、Cu和Zn元素的差異對該海拔處立柳生長的影響最為顯著。同時與170 m海拔以上區域的植株相比,經過3 a的冬季水淹后,165 m海拔處立柳的株高、基徑和冠幅受到了明顯抑制。所以持續的周期性水淹是否會進一步影響165 m海拔處植株葉、枝條和根中營養元素的含量與分配,進而影響立柳在該區域的生長乃至生存。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
