山東省水稻惡苗病病原菌的研究
2019-09-03 16:19:47張洪瑞楊軍袁守江李廣賢周學標徐建第陳博聰朱其松
山東農業科學 2019年7期
張洪瑞 楊軍 袁守江 李廣賢 周學標 徐建第 陳博聰 朱其松


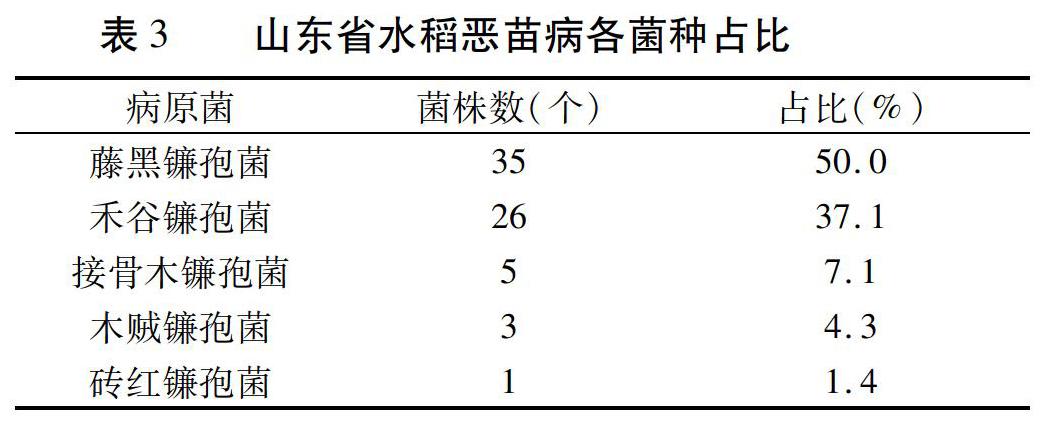
摘要:從山東省18個水稻種植區采集水稻惡苗病樣本,共分離獲得70個單孢菌株。參照Booth分類系統,這些菌株歸屬于5個鐮孢菌種,其中,藤黑鐮孢菌(Fusarium fujikuroi)占50.0%,禾谷鐮孢菌(Fusarium graminearum)占37.1%,接骨木鐮孢菌(Fusarium sambucinum)占7.1%,木賊鐮孢菌(Fusarium equiseti)占4.3%,磚紅鐮孢菌(Fusarium lateritium)占1.4%。本研究明確了山東省水稻惡苗病病原菌的菌種及優勢菌。
關鍵詞:山東省;水稻;惡苗病;病原鑒定;串珠鐮孢菌
中圖分類號:S435.111.4+4文獻標識號:A文章編號:1001-4942(2019)07-0080-03
水稻惡苗病(Fusarium moniliforme Sheld)是常見的水稻種傳真菌性病害,由無性態的串珠鐮孢菌、有性態的藤侖赤霉菌侵染引起,俗稱白稈病、搶先稻,又稱徒長病,從苗期至抽穗期均可發生[1,2],主要靠帶菌種子傳播[3-5]。水稻受惡苗病菌侵染后,病株在2 ~?4葉期即表現徒長,植株比健株高,葉片、葉鞘狹長,根系發育異常,生長細弱。病株一般不能抽穗或不能完全抽穗,即使抽穗,穗小而不實[6,7]。近年,隨著大面積稻草還田及水稻集中育秧的推廣,惡苗病發生越來越重,我國不少稻區普遍發生[8-10]。該病害發病率高,對水稻穩產、高產威脅大[6,11,12],一般減產10%~20%,嚴重的可減產50%以上[13,14]。
水稻惡苗病主要由串珠鐮孢菌引起,但一些研究者發現其他鐮孢菌也能致病[6,11],且不同地區病菌種類及其比例有所差異。王拱辰等[6]從300個單孢菌株中共分離出8種惡苗病菌,其中串珠鐮孢菌占總標本數的60.2%,是浙江水稻惡苗病的主要致病菌,木賊鐮孢菌、禾谷鐮孢菌、磚紅鐮孢菌、雪腐鐮孢菌、尖鐮孢菌、半裸鐮孢菌、腐皮鐮孢菌均有不同程度的弱致病性,其優勢種和不同種占比因地區和水稻品種的差異而有所變化。陜西漢中地區以串珠鐮孢菌為主[15]。湖北22個縣的水稻惡苗病菌,88.6%的菌株為串珠鐮孢浙江變種[11]。陳夕軍等[16]研究發現,江蘇省惡苗病以藤黑鐮孢菌為優勢菌(60.8%),雪腐鐮孢菌、半裸鐮孢菌、禾谷鐮孢菌、磚紅鐮孢菌和尖孢鐮孢菌分別占14.8%、6.9%、6.8%、6.2%和4.5%。除上述研究結果外,鄭鎬燮等[17]研究還表明,惡苗病有生理分化現象。
目前有關山東省水稻惡苗病的研究尚未見報道,因此,本研究通過采集、分離山東省各地區水稻惡苗病菌,明確其種類及優勢菌,以期為進一步研究該病菌對多菌靈、咪鮮胺、浸種靈等水稻處理劑的抗性以及抗性水稻種質資源篩選提供理論基礎,為該病害防治提供參考。
1 材料與方法
1.1 材料
2017年10月,從山東省濟寧、菏澤、臨沂、日照、東營等市的18個縣區采集水稻惡苗病樣本,經組織分離、純化再單孢分離,獲得70個菌株。
1.2 方法
1.2.1 病菌分離和保存
病菌分離:剪取10 cm長病葉鞘,去離子水沖洗去表面泥塵及病菌孢子,然后于無菌水中浸泡12 h,取出置于管底裝有吸水脫脂棉球的試管中,于25℃恒溫培養箱中進行培養,至葉鞘表面出現霉層后進行單孢分離。
病菌保存:取活化的菌餅(Φ 6 mm)于1.5 mL離心管中,加50%無菌甘油1 mL,-70℃保存。
1.2.2 種類鑒定 主要參照文獻[18]~文獻[20]的分類系統和方法進行鑒定。
2 結果與分析
2.1 水稻惡苗菌株分離純化
從山東省18個地區的病樣中共分離獲得惡苗病菌單孢菌株70個(表1)。其中,濟寧市魚臺縣清河鎮試驗基地、東營市墾利區和東營市東營區三村因保濕后病莖上未產生孢子或病樣污染嚴重,經多次分離、純化,均未能獲得純培養的鐮孢菌菌株。
2.2 菌株保存
所有菌株樣品均已保存在山東省水稻研究所超低溫冰箱中(-70℃)。
2.3 各分離菌株歸屬種類
根據參考文獻[18]~[20]的分類系統及相關性狀描述方法,將來自山東省18個縣區的水稻惡苗病菌鑒定為鐮孢菌屬的5個種,其中藤黑鐮孢菌為優勢種,共35個菌株,占50.0 %;其次是禾谷鐮孢菌,占37.1 %;其他3種菌分別是接骨木鐮孢菌、木賊鐮孢菌和磚紅鐮孢菌,占比分別為7.1%、4.3%和1.4%(表2、表3)。
3 討論與結論
江蘇、浙江、陜西漢中、湖北22個縣的水稻惡苗病菌只有藤黑鐮孢菌一種優勢菌[6,11,15,16],發病情況較為單一,比較好防治。從表2和表3可以看出,山東省各稻區水稻惡苗病菌共歸屬5個種,分別為藤黑鐮孢菌(即原來的串珠鐮孢菌)、禾谷鐮孢菌、接骨木鐮孢菌、木賊鐮孢菌和磚紅鐮孢菌,其中藤黑鐮孢菌和禾谷鐮孢菌為優勢菌,田間發病情況、發病癥狀較為復雜多變,防治較為困難。
本試驗結果可為進一步研究山東省水稻惡苗病田間發病、惡苗病菌對水稻處理劑的抗性以及抗惡苗病水稻種質資源篩選提供理論基礎。
參 考 文 獻:
[1]傅華欣, 朱龍粉, 顧文, 等. 16%惡線清WP浸種防治水稻惡苗病的效果[J]. 安徽農業科學, 2000, 28(6): 765-766.
[2] 方興洲, 陳莉, 產祝龍, 等. 水稻惡苗病與浸種、催芽和播種等因子的關系研究[J]. 熱帶作物學報, 2012, 33(6): 1107-1110.
[3] 沈迎春, 劉福海. 25%咪鮮胺乳油防治水稻惡苗病的效果[J]. 農藥科學與管理, 2003, 24(5): 19-21.
[4] 陳夕軍, 盧國新, 童蘊慧, 等. 水稻惡苗病菌對三種浸種劑的抗性及抗藥菌株的競爭力[J]. 植物保護學報, 2007, 34(4): 425-430.
[5] Desjardins A E, Plattner R D, Nelson P E. Production of fumonisin B (inf1) and moniliformin by Gibberella fujikuroi from rice from various geographic areas[J]. Applied and Environmental Microbiology, 1997, 63(5): 1838-1842.
[6] 王拱辰, 陳鴻逵, 徐沛生, 等. 水稻惡苗病病原菌的研究[J]. 植物病理學報, 1990, 20(2): 93-97.
[7] 產祝龍, 丁克堅, 檀根甲, 等. 水稻惡苗病發生規律的探討[J]. 安徽農業大學學報, 2004, 31(2): 139-142.
[8] 陳利峰, 徐敬友. 農業植物病理學[M]. 北京: 中國農業出版社, 2015.
[9] 季芝娟, 馬良勇, 李西明, 等. 水稻惡苗病抗性的研究進展[J]. 中國稻米, 2008(2): 24-25.
[10]趙淑元, 產祝龍, 丁克堅. 水稻惡苗病菌對使百克、多菌靈的抗性遺傳研究[J]. 安徽農業大學學報, 2008, 35(1): 46-48.
[11]羅俊國. 水稻惡苗病致病鐮孢種類及菌系研究[J]. 中國水稻科學, 1995, 9(2): 119-122.
[12]李海東, 方利民, 尤希宇, 等. 3種藥劑浸種對水稻安全性及惡苗病的防治效果[J]. 浙江農業科學, 2017, 58(12): 2235-2236, 2238.
[13]鄭鎬燮, 呂彬, 吳潤植, 等. 水稻惡苗病抗病性篩選方法的初步研究[J]. 植物保護學報, 1993, 20(4): 289-293.
[14]楊長登, 郭龍彪, 李西明, 等. 水稻抗惡苗病微效QTL的定位[J]. 中國水稻科學, 2006, 20(6): 657-659.
[15]陳嘉孚, 劉鑄德, 陸世英. 水稻惡苗病防治研究[J]. 西北農業大學學報, 1986, 14(1): 44-49.
[16]陳夕軍, 盧國新, 童蘊慧, 等. 江蘇水稻惡苗病病原菌研究[J]. 揚州大學學報(農業與生命科學版), 2008, 29(3): 88-90.
[17]鄭鎬燮, 呂彬, 吳潤植. 水稻惡苗病病原菌及其生物學特性的研究現狀[J]. 黑龍江農業科學, 1992(6): 4l-44.
[18]Booth C. The genus Fusarium[M]. UK: Commonwealth Mycological Institute, 1971.
[19]陳鴻逵, 王拱辰. 浙江省鐮刀菌志[M]. 杭州: 浙江科學技術出版社, 1992.
[20]Leslie J F, Summerell B A. The Fusarium laboratory manual[M]. USA: Wiley-Blackwell, 2006.
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
山東交通科技(2022年3期)2022-08-05 03:06:24
中國集體經濟(2022年9期)2022-04-12 01:07:35
雜技與魔術(2022年1期)2022-03-16 10:22:48
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
——山東省濟寧市老年大學之歌
老年教育(老年大學)(2021年10期)2021-11-12 09:15:48
今日農業(2021年14期)2021-10-14 08:35:40
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
