海螺溝冰川退縮區不同演替階段植被碳貯量與分配格局
2019-08-30 07:07:58何詠梅楊丹荔
西南林業大學學報 2019年5期
何詠梅 李 偉 楊丹荔,3 羅 輯
( 1. 中國科學院、水利部成都山地災害與環境研究所,四川 成都 610041;2. 中國科學院大學,北京 100049;3. 成都理工大學地球科學學院,四川 成都 610101)
全球變化研究是當今地球科學研究的重要議題之一。氣候變化對陸地生態系統的影響是多尺度、多層次的,主要表現在生態系統的結構和功能方面[1],生態系統碳循環研究是全球變化科學中的重要組成部分,也是研究生態系統對氣候變化貢獻和響應的重點之一[2]。森林生態系統是陸地生態系統的主體,世界的森林生態系統儲存了陸地生態系統大部分的有機碳,在陸地碳循環中發揮著重要作用[3-4]。《京都議定書》正式生效后,對森林生態系統碳貯量的研究顯得更為迫切。目前對森林碳貯量的估計,主要是通過直接或間接測算森林生物量或現存量再乘以含碳率推算而得。因此,森林群落的生物量及其組成樹種的含碳率是研究森林碳貯量的兩個關鍵因子[5]。森林生態系統由于其自身的多樣性和復雜性,其中的碳循環過程存在很大的不確定性,隨著氣候、土壤等生態因子的地帶性變化,生態系統碳循環和碳貯量都呈現出相應的空間變化規律[3,6]。貢嘎山海螺溝冰川在過去150多a的時間里,后退幅度十分顯著,原生裸地近地表環境變化迅速,土壤有機質積累過程明顯。退縮區發育著完整的植被演替序列,呈現著植物群落演替在時空上的連續性和生態環境變化的規律性[7-8]。演替序列每個階段碳循環密切相關,由此可以準確測定演替序列的碳動態,有助于把握演替序列生態系統中營養元素的循環和利用方式,進而深入研究演替序列生態系統的格局和過程,指導植被生態恢復建設等。本研究通過測定海螺溝冰川退縮區植被演替序列各個階段生物量及不同樹種含碳率,構建了原生演替不同階段的碳分配和碳貯量格局,對研究原生演替過程中的碳循環有積極意義。
1 研究區概況
貢嘎山位于青藏高原東部邊緣,地處東經101°30′~102°15′,北緯 29°20′~30°20′,面積約10 000 km2,主峰海拔7 556 m,是橫斷山主要的季風海洋性冰川作用中心,區內新構造運動強烈,地貌為高山峽谷類型,自然條件復雜。本研究選擇貢嘎山海螺溝冰川近150 a形成的冰川退縮區為研究區(29°34′21″N,102°59′42″E)(圖1),該區域海拔2 800~2 950 m,氣候濕冷,屬于山地寒溫帶氣候類型,年平均氣溫4.2 ℃,年降水日數達260 d以上,年平均降水量約1 947 mm。海螺溝是貢嘎山東坡最主要的冰川河,水熱條件配合好,冰川消融速度快,近百年來沒有冰進過程,其冰川退縮區內發育著自小冰期以來形成的完整植被演替序列,并且該區域土壤有連續成土過程,生境變化具有周期短、更替速度快的特點,是進行全球變化研究的理想場所。在該研究區長達2 km的范圍內,人為干擾很小,有利于測定該區域植被演替過程中的碳動態。

圖 1 海螺溝冰川退縮區樣地分布Fig. 1 Map of sampling plots in Hailuogou glacier retreat area
2 研究方法
2.1 樣品采集與測定
冰川退縮后形成原生裸地,3~5 a就有先鋒植物不斷侵入。在柳樹(Salix rehderana)、冬瓜楊(Populus purdomii)、沙棘(Hippophae rhamnoides)主導的演替前期階段,由于它們的生態位相近,競爭加劇。隨著演替進行,各階段優勢種不斷更替,最終形成以峨眉冷杉(Abies fabri)和麥吊云杉(Picea brachytyla)為主的頂極群落。基于前人對海螺溝冰川退縮區研究基礎[9-10],選取了8個典型樣地,各樣地冰川退縮時間、林分密度、優勢物種組成狀況見表1。
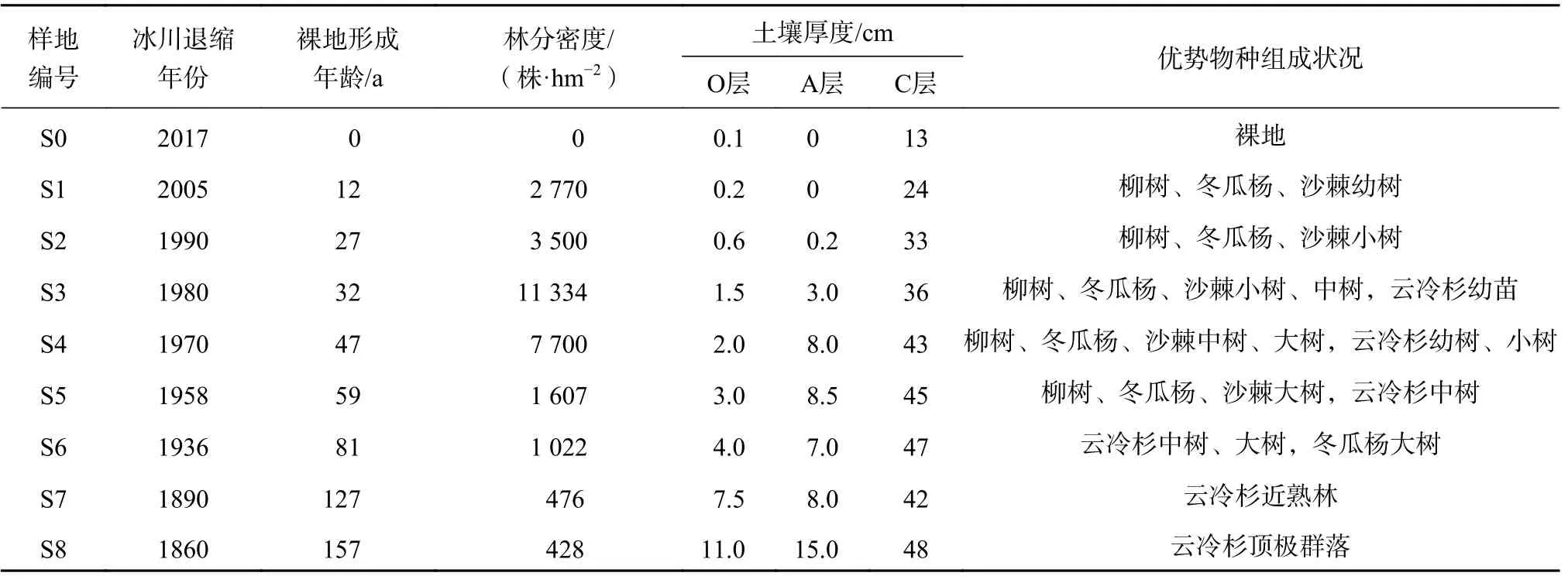
表 1 不同演替階段群落特征Table 1 Species characteristics of plots at different stand ages
于2017年4月和8月在各個樣地隨機布設3個10 m×10 m的樣方,記錄樣方內喬木的名稱、基徑、胸徑、樹高、冠幅、存活狀態等,并沿樣方對角線布設3個1 m×1 m的小樣方,調查樣方內所有灌木、草本、地被物,記錄植物名稱,數量等。同時,在樣地內隨機布設3個0.5 m×0.5 m的小樣方,用于收集凋落物。樣地內不同喬木各器官除單獨采集樣品外,還將在計算得出生物量后根據生物量比例將不同樹種相同器官樣品混合取樣。采用收獲法收獲樣方內所有灌木、草本及地被物,記錄總重后采集樣品。粗木質殘體包括立枯、倒木和站樁,在樣方內采集立枯、倒木和站樁的皮和木質部,組成混合樣。在每個樣地挖掘3個土壤剖面采集土壤樣,按照野外土壤發育性狀,將土壤按以下層次采集:O層,完全分解的腐殖質層;A層,腐殖質積累的礦質土表層;C層,母質層。記錄每層厚度,挑出土壤混合樣中的根系和石礫,采用環刀法測土壤容重。
2.2 碳儲量計算方法
所有植物樣品帶回實驗室,放入85 ℃的恒溫箱中烘至恒重,土壤樣做風干處理,磨碎待用。本研究中樣品含碳率的測定采用Elementar Vario Macro Cube C測定儀測定(德國,檢測下限≤10 μg/g)。
喬木層生物量根據羅天祥等[11-13]的生物量計算方式計算得出;粗木質殘體生物量根據張修玉等[14-15]的計算方法得出,再采集樣地內具代表性的立枯、倒木作為標準參照物帶回實驗室稱量其干質量后進行校正;土壤碳貯量依據以下公式計算:
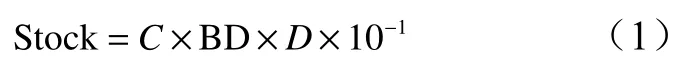
式中:Stock表示碳元素在各土層里的貯量(kg/hm2);C表示碳元素在土層中的濃度(mg/kg);BD表示土層的容重(g/cm3);D表示土層厚度(cm)。
2.3 分析方法
所有數據分析和圖形制作均使用SPSS和Excel軟件完成。采用單因素方差分析法(one-way ANOVA)分析不同演替階段植被含碳率的差異,并采用最小顯著差法(LSD)進行多重比較。
3 結果與分析
3.1 喬木層主要樹種碳庫構成
在海螺溝冰川退縮區植被演替的各個階段,喬木層優勢種含碳率存在差異,各器官間含碳率也存在差異。在演替前期(S1~S3),群落內植被含碳率表現為沙棘(47.46%)>柳樹(46.45%)>冬瓜楊(45.63%),在演替中期(S4~S5),群落內植被含碳率表現為云冷杉(48.37%)>沙棘(46.27%)>冬瓜楊(45.53%)>柳樹(45.26%),在演替后期(S6~S8),群落內云冷杉含碳率(48.43%)顯著大于冬瓜楊(45.85%)(P<0.05)。在整個演替序列上,針葉樹種的平均含碳率(48.4%)顯著高于闊葉樹種(46.15%)(P<0.05)。
海螺溝冰川退縮區原生演替序列上喬木層主要樹種碳貯量見表2。由表2可知,隨著演替的進行,喬木層碳貯量呈顯著增加的趨勢。從S1到S8,樣方內喬木層碳貯量分別為3.07、40.52、86.09、94.90、 103.07、 129.61、 167.16、 180.52 t/hm2,平均增長率為 1.22 t/(hm2·a);其中 S2 到 S3 階段增長最快,平均增長率為4.6 t/(hm2·a),S7階段到S8階段增長速率最慢,平均增長率為0.45 t/(hm2·a)。除 S3 和 S4 2 個階段間不存在顯著差異外,其余各階段間均存在顯著性差異(P<0.05)。在演替前期S1階段,群落內沙棘碳貯量最大,其次是柳樹,冬瓜楊碳貯量最小,在S2階段和S3階段則呈現出相反的趨勢,冬瓜楊碳貯量最大,且顯著高于沙棘和柳樹(P<0.05)。在演替中期,云冷杉碳貯量極低,冬瓜楊碳貯量在該時期達到最大,沙棘和柳樹碳貯量迅速降低,在S5階段,沙棘和柳樹總碳貯量只占群落碳貯量的5%。演替后期,云冷杉占據群落內主導地位,碳貯量迅速增加,占群落內碳貯量的85%以上。從植物各器官來看,各階段碳貯量表現為樹干>樹根>樹枝>樹皮>樹葉。
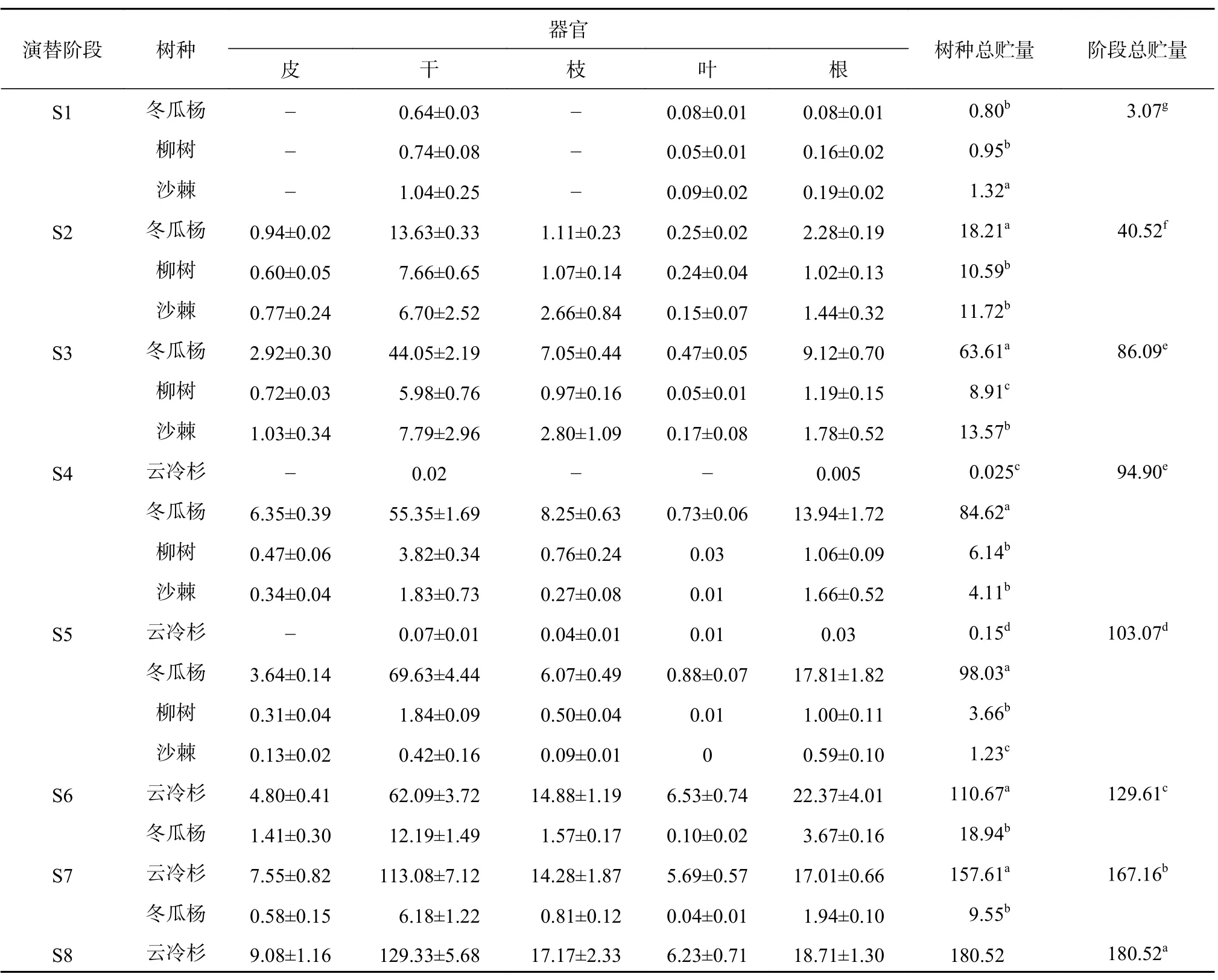
表 2 原生演替序列喬木樹種碳貯量Table 2 Carbon storage of arbor species of primary succession sequence t/hm2
3.2 灌木、草本、地被層碳庫構成
在演替的各個階段,林下的灌木、草本、地被層碳貯量都較小,所占比例較低(圖2)。根據相關研究[16],以含碳率45%計算灌木層碳貯量,以含碳率41%計算草本和地被層含碳率。隨著演替的進行,總碳貯量由最初的0.12 t/hm2逐漸增加到頂級群落的12.41 t/hm2,其中灌木層所占比例最高,為63.21%,草本層所占比例最小,不足3%。在整個演替序列上,灌木層貯量表現為持續增加,除演替前期各階段間無顯著性差異外,其余各階段間均存在顯著性差異(P<0.05);草本層碳貯量表現為先增加后減小再增加的趨勢,在S5階段達到最大,S6階段碳貯量最低,S5階段草本碳貯量與S4、S8階段無顯著性差異,但顯著大于其余各階段碳貯量(P<0.05);地被層碳貯量在各階段波動較大,但總體上碳貯量呈增加趨勢,S6階段碳貯量最大,顯著大于其余階段碳貯量(P<0.05)。

圖 2 原生演替序列草本層、地被層、灌木層碳貯量Fig. 2 Carbon storage of herb layer, ground cover and shrub layer of primary succession sequence
3.3 凋落物、粗木質殘體碳庫構成
森林中凋落物和粗木質殘體中的碳是森林生態系統碳庫的重要組成部分,二者含碳率以陳有超[17]研究中的0.46為準,計算得出演替各階段的碳貯量,見圖3。由圖3可知,凋落物碳貯量隨著演替的進行呈現上升趨勢,由S1階段的0.44 t/hm2增加到S8階段的1.29 t/hm2,其中S7階段略有下降,但波動幅度較小,S1階段凋落物碳貯顯著低于其余各階段,S2~S8階段間均無顯著性差異。演替過程中群落的葉量和小枝量都是逐步增加,但是在S3階段的種間競爭產生大量枯枝,S1~S5階段是落葉闊葉林階段,每年落葉較多,從S7階段開始發展到常綠的暗針葉林階段,落葉較少,由此形成凋落物碳貯量隨著演替進程的波動。

圖 3 凋落物和粗木質殘體碳貯量Fig. 3 Carbon storage of litters and coarse wood residues
粗木質殘體的碳貯量表現為先增加后減小,在S6階段達到最大值15.68 t/hm2,之后有所下降,但總體上碳貯量是增加的。除S5、S6、S7階段間碳貯量無顯著性差異外,其余各階段碳貯量均存在顯著性差異(P<0.05)。S1階段沒有粗木質殘體,隨后的競爭產生了枯立木和倒木,S4、S5階段主要是沙棘和柳樹死亡后產生的倒木,S6、S7主要是冬瓜楊死亡產生的倒木。
演替最初階段,凋落物碳貯量在二者中所占比例高達95%,隨著演替的進行,組成群落的種群種內、種間競爭加劇,產生大量死樹,粗木質殘體快速增加,凋落物碳貯量增幅較低,所占比例下降到不足1%。各階段中二者總貯量的變化也表現為先增加后減小,和粗木質殘體變化趨勢一致,這主要是由于演替中后期有大量的柳樹和冬瓜楊大樹死亡,粗木質殘體迅速上升所致。
3.4 原生演替序列土壤碳庫構成
各演替階段土壤碳貯量隨著演替的進行逐漸增加,從S1階段的0.98 t/hm2增加到S8階段的144.88 t/hm2,增長近150倍,各階段間土壤碳貯量均存在顯著性差異(P<0.05)。S1到S2階段不僅凋落物量少,而且凋落物分解還在進行,土壤碳貯量增加緩慢,年增長速率為0.54 t/(hm2·a);S2到S3階段增長速率最快,平均年增長率為2.22 t/(hm2·a),且柳樹、冬瓜楊、沙棘競爭劇烈,產生大量枯枝、倒木,輸入土壤的碳較多。土壤碳貯量一直呈現逐漸增加趨勢,到頂級群落土壤碳貯量還遠低于貢嘎山峨眉冷杉成熟林的土壤碳貯量(291.8 t/hm2)[18],與我國森林土壤和世界土壤平均碳貯量(193.6、189 t/hm2)[3,19-20]相比,土壤碳貯量存在一定差距,土壤還沒有形成真正的淀積層,說明海螺溝冰川退縮區土壤發育還沒有達到成熟狀態,土壤發育還將持續進行[21]。
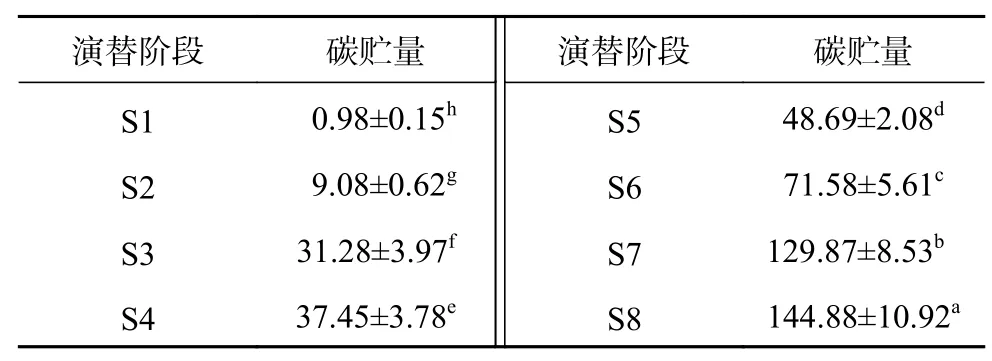
表 3 原生演替序列土壤碳貯量Table 3 Soil carbon storage of primary succession sequence
3.5 原生演替序列碳貯量動態變化
由各階段各層碳貯量得出原生演替森林生態系統各階段的總碳貯量,見圖4。隨著演替進行總碳貯量逐漸增加,由S1階段的4.63 t/hm2增加到頂級群落的346.10 t/hm2,除S7與S8無顯著性差異外,其余各階段間均存在顯著性差異,且顯著小于S7、S8(P<0.05)。其中喬木層和土壤碳貯量在整個演替序列上所占比例均超過85%,其中S2、S3、S7、S8階段所占比例超過90%。在S2階段二者碳貯量達到該階段總貯量的96%,其余部分只占4%。其中土壤所占比例持續增加,由S1的21.20%增加到S8的41.86%,而喬木所占比例則呈現先上升后下降的趨勢,由S1的66.37%下降到S8的52.16%。S7、S8階段的總貯量分別為321.02、346.1 t/hm2,遠高于我國森林生態系統的平均碳貯量(258.8 t/hm2)[20-21],但與我國針葉林生態系統的平均碳貯量(408 t/hm2)相比還較低[22-24]。

圖 4 原生演替序列碳貯量動態變化Fig. 4 Dynamic changes of carbon storage of primary succession sequence
4 結論與討論
樹種之間含碳率的差異,主要是由于喬木種類之間自身結構的不同而導致的[5]。海螺溝冰川退縮區植被含碳率與唐宵等[25]、黃從德等[26]對四川主要針葉樹種含碳率的研究結果、張萍[27]對北京延慶縣9個主要森林類型含碳率的研究結果、馬欽彥等[28]對華北主要森林類型建群種含碳率的研究結果基本一致。
海螺溝冰川退縮區原生演替不同階段植物體不同部位含碳率分布為41.82%~50.9%,范圍略大于周鵬[16]在貢嘎山典型植被帶上所測得的植被含碳率(43.72%~48.81%);略小于陳有超[17]在貢嘎山東坡峨眉冷杉林所測得的含碳率(41.50%~53.4%);王金亮等[5]對滇西北4種不同樹種各器官的含碳率進行了測定,結果為45.09%~56.02%,顯著大于本研究區植被含碳率,這可能與當地的地理位置和樹種有關。在整個演替序列上,樹枝和樹葉含碳率相對較大,其余部位含碳率相對較低,主要是因為樹枝含有較多的木質素,而木質素的含碳率較高,而樹葉有豐富的葉綠體,有大量有機物存在所引起的[5]。在演替前期(S1~S3),群落植被含碳率表現為根系含碳率最低(45.72%),由于在演替前期植物根系對土壤中營養元素的吸收與利用過程是能量的消耗過程。無論是元素的獲取過程還是元素在植物體內的運輸、同化和分配過程都需要消耗能量,能量最終來源于植物體內碳水化合物的轉化消耗。其中對吸收起重要作用的細根的周轉是最大的碳消耗過程[29]。
在海螺溝冰川退縮區長達2 km的植被演替序列上,植物群落與環境相互作用,生物地球化學循環逐步加強[30]。不同演替階段植被類型不同,種類組成也不同。不同種類的器官和組織含碳率不同,同一種類在不同演替也存在差異。植被演替序列生態系統的碳庫構成變化較大,碳貯量存在逐步增加的趨勢。在演替前期,喬木層樹種以冬瓜楊、柳樹、沙棘為主,林下草本、灌木等都較少。S1階段,各樹種間碳貯量差異不大,在S2、S3階段,冬瓜楊憑借其較快的高生長和較高的光合速率在群落內占據主導地位[31],碳貯量迅速增加,而柳樹和沙棘在種間競爭中不敵冬瓜楊,碳貯量遠小于冬瓜楊。該時期群落內凋落物和粗木質殘體碳貯量也較小。隨著演替持續進行,林下生境特征發生改變,云冷杉順利進入到群落中。在演替中期,云冷杉主要以幼苗及小樹為主,碳貯量較低,該時期冬瓜楊占據主林層,碳貯量達到整個演替序列的最大值,而沙棘和柳樹在該時期逐漸死亡,碳貯量持續下降。由于柳樹和沙棘在該時期大量死亡,林下粗木質殘體碳貯量與演替前期相比顯著增加。同時由于群落內大量林窗出現,林下的灌木、草本等數量有所上升。在演替后期,群落進入到以冬瓜楊和云冷杉為主的針闊混交階段,云冷杉在該時期生長較快,迅速占據主林層,碳貯量迅速增加,林下的冬瓜楊由于接受不到足夠的光照而逐漸死亡,碳貯量迅速減小。在S6階段,林下的草本層碳貯量不足0.1 t/hm2,而地被層碳貯量達到整個演替序列的最大值,這主要是由于在這一階段林冠層郁閉度增大使得林下草本層植物所能利用的光照減少,該階段林下空氣濕度較大,為苔蘚、石松等地被物的生長提供了有利條件。
海螺溝冰川退縮區植被演替是一個有序的、可觀測的連續過程,經過150多a的發展,最終由裸地演替為以云冷杉為主的頂級群落。各演替階段不同樹種各器官間的碳含量分布在41.8%~50.9%之間,總體表現為枝和葉碳含量較高,根碳含量較低。各器官含碳量的變異性較小,均不超過0.05。群落碳貯量則隨著演替的進行不斷增加,其中喬木層和土壤碳貯量在整個演替序列上所占比例均超過85%。土壤碳貯量隨著演替逐漸增加,但頂級群落林下土壤碳貯量遠低于貢嘎山峨眉冷杉成熟林的土壤碳貯量,說明海螺溝冰川退縮區土壤發育尚未完成,土壤發育還將持續進行。
原生演替開始時,在貧瘠生境上的養分積累是決定演替進程的重要因素[32]。最初的先鋒植物對氮需求不高,在群落演替初期碳貯量迅速增加,群落中的固氮植物和微生物的作用逐步增強。隨后土壤氮貯量迅速增加,植被演替動態取決于固氮植物的種類、密度、生活型以及碳氮比。演替初期圍繞著氮爭奪,冬瓜楊、柳樹、沙棘種間競爭劇烈,由于冬瓜楊光合速率高,碳積累較多,同時也可以獲得更多營養元素,生長迅速的冬瓜楊在演替前期、中期處于優勢地位;演替中期群落主要是冬瓜楊的種內斗爭,產生大量死樹和倒木,養分的生物循環加強,土壤有機質迅速增加;演替后期的暗針葉林優勢種個體碳貯量最大,喬木層和表土層貯存了大量養分,而生態系統養分的生物地球化學循環與以前明顯不同。
以前對原生演替的研究都沒有在一個連續完整的序列開展,這是形成這些問題的主要原因。海螺溝冰川退縮區植被演替是一個有序的、可觀測的連續完整過程,經過150多a的發展,最終由裸地演替為以云冷杉為主的頂級群落。通過碳動態可以量化這一群落變化過程中種間、種內競爭關系和競爭的結果,量化凋落物和粗木質殘體在土壤發育過程中的作用。今后需要深入研究植物營養元素循環和利用規律,揭示原生演替的碳素和養分共同驅動機制,豐富生態學理論。
[ 參 考 文 獻 ]
[1]Walther G R, Post E, Convey P, et al. Ecological responses to recent climate change [J]. Nature, 2002,416(6879): 389-395.
[2]劉紀遠, 于貴瑞, 王紹強, 等. 陸地生態系統碳循環及其機理研究的地球信息科學方法初探 [J]. 地理研究,2003, 22(4): 397-405.
[3]Dixon R K, Solomon A M, Brown S, et al. Carbon pools and flux of global forest ecosystems [J]. Science,1994, 263(5144): 185-190.
[4]程鵬飛, 王金亮, 王雪梅, 等. 森林生態系統碳儲量估算方法研究進展 [J]. 林業調查規劃, 2009, 34(6):39-45.
[5]王金亮, 王小花, 岳彩榮, 等. 滇西北香格里拉森林4個建群種的含碳率 [J]. 生態環境學報, 2012, 21(4):613-619.
[6]曹明奎, 于貴瑞, 劉紀遠, 等. 陸地生態系統碳循環的多尺度試驗觀測和跨尺度機理模擬 [J]. 中國科學(D輯: 地球科學), 2004, 34(S2): 1-14.
[7]Alexander E B, Burt R. Soil development on moraines of Mendenhall Glacier, southeast Alaska. 1. The moraines and soil morphology [J]. Geoderma, 1996,72(1/2): 1-17.
[8]Egli M, Mavris C, Mirabella A, et al. Soil organic matter formation along a chronosequence in the Morteratsch proglacial area (Upper Engadine, Switzerland) [J]. Catena, 2010, 82(2): 61-69.
[9]鐘祥浩, 羅輯, 吳寧. 貢嘎山森林生態系統研究[M].成都: 成都科技大學出版社, 1997.
傳統民居元素是傳統文化重要組成部分,應用于陶瓷藝術設計中,自身的裝飾功能突顯,可烘托出陶瓷藝術作品的文化底蘊、幫助其創造傳統文化氛圍。對于陶瓷藝術作品的欣賞,不能僅觀察其外在表現,更應當注重其內涵、文化品位的挖掘,將傳統民居元素對陶瓷藝術作品的文化功能充分發揮。
[10]陳富斌, 羅輯. 貢嘎山高山生態環境研究: 第 2卷 [M]. 北京: 氣象出版社, 1998.
[11]羅天祥, 石培禮, 羅輯, 等. 青藏高原植被樣帶地上部分生物量的分布格局 [J]. 植物生態學報, 2002, 26(6):668-676.
[12]劉興良, 馬欽彥, 楊冬生, 等. 川西山地主要人工林種群根系生物量與生產力 [J]. 生態學報, 2006, 26(2):542-551.
[13]李登秋, 居為民, 鄭光, 等. 基于生態過程模型和森林清查數據的森林生長量估算對比研究 [J]. 生態環境學報, 2013, 22(10): 1647-1657.
[14]張修玉, 管東生, 張海東. 廣州三種森林粗死木質殘體(CWD)的儲量與分解特征 [J]. 生態學報, 2009,29(10): 5227-5236.
[15]何帆, 王得祥, 張宋智, 等. 小隴山林區主要森林群落凋落物及死木質殘體儲量 [J]. 應用與環境生物學報,2011, 17(1): 46-50.
[16]周鵬. 貢嘎山東坡垂直帶譜典型植被類型固碳分異及其影響因子[D]. 北京: 中國科學院大學, 2013.
[18]程根偉, 羅輯. 貢嘎山亞高山林地碳的積累與耗散特征 [J]. 地理學報, 2003, 58(2): 179-185.
[19]潘輝, 黃石德, 洪偉, 等. 3種相思人工林生態系統碳貯量及分配 [J]. 福建林學院學報, 2009, 29(1): 28-32.
[20]楊衛星, 李春寧, 付軍, 等. 桂西南連續年齡序列尾巨桉人工林碳儲量及其分布特征 [J]. 農業研究與應用,2017(3): 24-30.
[21]羅輯, 李偉, 佘佳, 等. 貢嘎山海螺溝冰川退縮區植被演替過程的碳動態 [J]. 山地學報, 2017, 35(5):629-635.
[22]周玉榮, 于振良, 趙士洞. 我國主要森林生態系統碳貯量和碳平衡 [J]. 植物生態學報, 2000, 24(5):518-522.
[23]呂曉濤, 唐建維, 于貴瑞, 等. 西雙版納熱帶季節雨林的C貯量及其分配格局 [J]. 山地學報, 2006, 24(3):277-283.
[24]涂潔, 劉琪璟. 亞熱帶紅壤丘陵區濕地松人工林生態系統碳素貯量與分布研究 [J]. 江西農業大學學報,2007, 29(1): 48-54.
[25]唐宵, 黃從德, 張健, 等. 四川主要針葉樹種含碳率測定分析 [J]. 四川林業科技, 2007, 28(2): 20-23.
[26]黃從德, 張健, 楊萬勤, 等. 四川省及重慶地區森林植被碳儲量動態 [J]. 生態學報, 2008, 28(3): 966-975.
[27]張萍. 北京森林碳儲量研究[D]. 北京: 北京林業大學, 2009.
[28]馬欽彥, 陳遐林, 王娟, 等. 華北主要森林類型建群種的含碳率分析 [J]. 北京林業大學學報, 2002, 24(S1):100-104.
[29]王琪, 徐程揚. 氮磷對植物光合作用及碳分配的影響 [J]. 山東林業科技, 2005, 35(5): 59-62.
[30]宋蒙亞, 余雷, 姜永雷, 等. 貢嘎山冰川退縮跡地植被原生演替驅動機理探究[C]//中國地理學會經濟地理專業委員會. 2017年中國地理學會經濟地理專業委員會學術年會論文摘要集. 成都: 中國地理學會,2017: 38.
[31]楊丹丹, 羅輯, 佘佳, 等. 貢嘎山海螺溝冰川退縮區原生演替序列植被生物量動態 [J]. 生態環境學報,2015, 24(11): 1843-1850.
[32]何磊, 唐亞, 張繼娟, 等. 原生演替及其在生態恢復中的應用 [J]. 四川師范大學學報(自然科學版), 2010,33(3): 393-402.
