2,4-D處理對大百合種子休眠解除及內(nèi)源激素變化的影響
2019-08-30 07:07:54李鳳榮李葉芳馬詩雨關(guān)文靈
西南林業(yè)大學(xué)學(xué)報 2019年5期
關(guān)鍵詞:差異
李鳳榮 李葉芳 馬詩雨 關(guān)文靈
( 云南農(nóng)業(yè)大學(xué)園林園藝學(xué)院,云南 昆明 650201)
大百合(Cardiocrinum giganteum)是百合科(Liliaceae)大百合屬(Cardiocrinum)多年生鱗莖類草本植物[1],該屬植物因植株巨大并具有網(wǎng)狀脈的葉片而顯著區(qū)別于百合屬(Lilium)植物[2]。大百合植株粗壯挺拔,葉片碩大,花朵潔白芳香,具有極高的觀賞價值,其鱗莖富含多種營養(yǎng)物質(zhì),可供食用[3],也可藥用[4];果實可入藥,作為馬兜鈴(Aristolochia debilis)的代用品[5],是優(yōu)良的野生植物資源。
國內(nèi)關(guān)于大百合的研究主要集中在其引種栽培[6]、組織培養(yǎng)[7]、細(xì)胞水平[8]以及雜交[9]等方面。而國外鮮有對大百合屬的研究,主要在其種子生物學(xué)[10]、生活史[11]、花粉發(fā)育[12]等方面。關(guān)文靈等[13]的研究表明大百合種子存在形態(tài)和生理后熟現(xiàn)象,其休眠類型屬于復(fù)雜的形態(tài)生理休眠。蔡薇等[14]研究變溫層積過程中加赤霉素(GA3)、6-芐氨基腺嘌呤(6-BA)、硝普鈉(SNP)處理對大百合種胚后熟的影響。2,4-二氯苯氧乙酸(2,4-D),是一種人工合成的植物生長素,具有促進(jìn)細(xì)胞生長與分化,加速根、芽伸長,促進(jìn)果實形成與種子萌發(fā)等作用[15]。目前,鮮見2,4-D浸種處理對大百合種子休眠解除的研究,也未見內(nèi)源激素在大百合種子變溫層積過程中變化的研究。本研究采用2,4-D浸種處理結(jié)合變溫層積的方法,探究對大百合種子休眠破除的影響,篩選出適宜的濃度,在最佳處理濃度的基礎(chǔ)上,研究大百合種子在不同時期內(nèi)源激素的動態(tài)變化,為揭示內(nèi)源激素在大百合種子休眠解除過程中的作用提供參考,為種子育苗生產(chǎn)實踐提供科學(xué)依據(jù)。
1 材料與方法
1.1 試驗材料
試驗所用大百合種子,于2015年11月采自云南省哀牢山國家級自然保護(hù)區(qū)(東經(jīng)100°44′~101°30′,北緯 23°36′~24°56′),在 4 ℃ 低溫環(huán)境下儲存?zhèn)溆谩T谧儨貙臃e結(jié)合激素處理的不同時期,對大百合種子取樣,取樣時間為層積后 0、30、60、90、120、130 d,在各時間點定期取樣,樣品經(jīng)液氮冷凍后,置于-80 ℃超低溫冰箱保存,用于測定種胚發(fā)育過程中植物內(nèi)源激素。
1.2 試驗方法
1.2.1 預(yù)處理方法
以蒸餾水為對照(CK),將大百合種子置于質(zhì)量濃度分別為1、5、10 mg/L的2,4-D溶液中浸泡48 h;每個處理種子150粒,設(shè)置3個重復(fù)。將上述浸種的種子裝入尼龍網(wǎng)袋中,再用潮濕的苔蘚覆蓋網(wǎng)袋,最后放入自封袋中,并在自封袋上用牙簽戳 3~4個小孔以保持透氣性。在恒溫箱中進(jìn)行模擬夏季—秋季—冬季的高溫到低溫的變溫 25 ℃/15 ℃( 60 d)、15 ℃/5 ℃( 60 d)、4 ℃(50 d)層積處理,周期性光照(光照12 h,黑暗12 h)。
1.2.2 指標(biāo)測定方法
1)胚長、胚生長率和發(fā)芽率的測定 在變溫層積90、105、120、135 d時,每個處理中隨機抽取10粒種子,在體視顯微鏡(徠卡公司,德國)下沿著與胚平行的方向,測量胚的長度。在變溫層積150 d時,隨機選取100粒,按公式(1)計算胚生長率,共重復(fù)3次。當(dāng)胚根突出珠孔1 mm時,種子視為已萌發(fā),觀察并記錄種子發(fā)芽起始時間。每隔5 d,統(tǒng)計100粒種子中的發(fā)芽率,并按公式(2)計算。

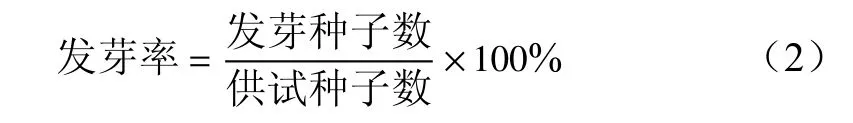
2)內(nèi)源激素含量的測定 在變溫層積期間,定期對處理組(5 mg/L 2,4-D)和CK浸泡48 h的種子分別取樣,經(jīng)液氮冷凍后,保存于-80 ℃超低溫冰箱中;用液-質(zhì)聯(lián)用色譜儀(安捷倫,美國)測定GA3、生長素(IAA)及脫落酸(ABA)的內(nèi)源激素含量,具體方法參照參考文獻(xiàn)[16]。
1.3 數(shù)據(jù)分析
采用SPSS 19.0對種胚長度、胚生長率進(jìn)行單因素ANOVA的Duncan多重比較方法進(jìn)行顯著性差異分析,對內(nèi)源激素ABA、IAA和GA3含量進(jìn)行單樣本T檢驗分析,使用Excel 2013進(jìn)行數(shù)據(jù)統(tǒng)計和圖表制作。
2 結(jié)果與分析
2.1 不同濃度2,4-D浸種對大百合種胚長度的影響
由圖1可知,隨著層積時間的增加,不同處理大百合種胚的長度均增長,但在層積90 d時,不同處理間胚長差異不顯著;層積105 d時,CK與5 mg/L 2,4-D處理差異顯著(P<0.05),與10 mg/L 2,4-D處理差異極顯著(P<0.01),CK的胚長分別高出5 mg/L 2,4-D和10 mg/L 2,4-D的胚長8.38%、14.74%。層積120 d時,CK與其他處理差異顯著(P<0.05),與5 mg/L和10 mg/L 2,4-D顯著極差異(P<0.01),CK的胚長分別高出5 mg/L 2,4-D和10 mg/L 2,4-D的胚長15.28%、15.88%。層積135 d時,CK與5 mg/L 2,4-D處理顯著差異(P<0.05),5 mg/L 2,4-D處理的胚長高出CK的胚長9.03%。

圖 1 不同濃度2,4-D處理下大百合種胚的長度變化Fig. 1 Length changes of C. giganteum embryos under different concentrations of 2,4-D
2.2 不同濃度2,4-D對大百合種子發(fā)芽的影響
由表1可知,CK的胚生長率與5 mg/L 2,4-D和 10 mg/L 2,4-D差異極顯著(P<0.01);5 mg/L 2,4-D處理的胚生長率最高,達(dá)到82%,比CK高出10%;10 mg/L 2,4-D處理的胚生長率最低,為52.33%,比CK低19.67%。5 mg/L 2,4-D的發(fā)芽起始時間最早,為4 ℃下14 d,比CK早11 d。

表 1 大百合種子胚生長率和發(fā)芽時間Table 1 Seed embryo growth rate and germination time of C. giganteum
由圖2可知,1 mg/L 2,4-D處理下,種子發(fā)芽率高于CK 3%,5 mg/L 2,4-D的發(fā)芽率最高,達(dá)75.33%,高于CK 15.33%,可促進(jìn)大百合種子的萌發(fā);10 mg/L 2,4-D的發(fā)芽率最低,為36.33%,明顯低于CK,低了30%。結(jié)合來看,5 mg/L 2,4-D處理的胚生長百分比最高,發(fā)芽時間最早,發(fā)芽率最高,利于大百合種胚后熟,可以縮短發(fā)芽時間,提高種子萌發(fā)。
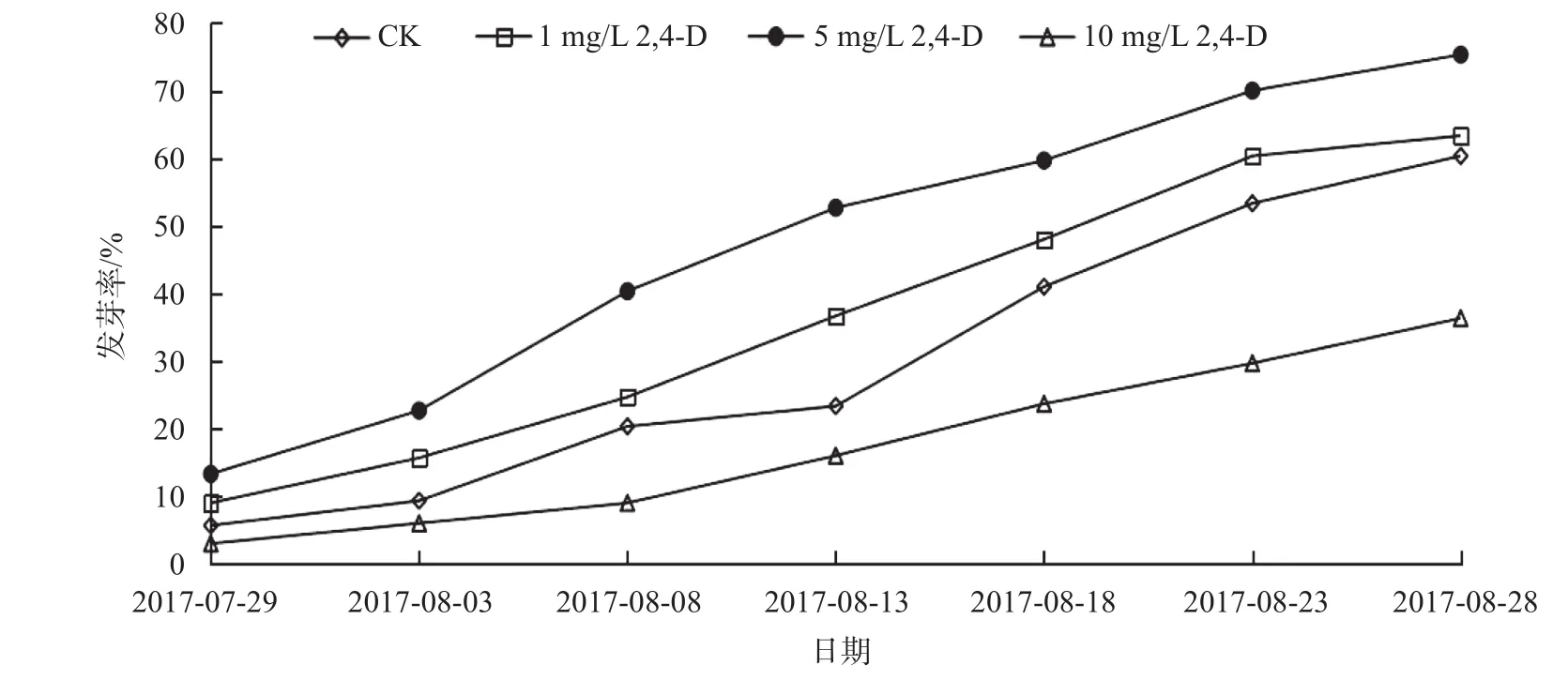
圖 2 不同濃度2,4-D處理下大百合種子的發(fā)芽率Fig. 2 Germination rate of C. giganteum seeds treated with 2,4-D with different concentrations
2.3 大百合種子層積過程中內(nèi)源激素的動態(tài)變化
隨著層積時間的增加,在層積過程中,大百合種子ABA、IAA和GA3含量的變化表現(xiàn)出不同的規(guī)律,見圖3。

圖 3 大百合種子層積過程中內(nèi)源激素含量的變化Fig. 3 Variation of endogenous hormones content in the process of C.giganteum seeds stratification
由圖3a可知,在大百合種子層積過程中,CK和2,4-D處理在ABA含量的變化總體呈下降趨勢。在未變溫層積0 d時差異極顯著(P<0.01),2,4-D處理比 CK高了 14.53 ng/g;在層積30 d時差異極顯著(P<0.01),2,4-D處理的比CK低了9.55 ng/g;在層積120 d時差異極顯著(P<0.01),CK比2,4-D低了5.59 ng/g;在層積60、90、130 d時,處理差異不顯著。CK中ABA含量在層積60 d時,下降速度較快,層積30 d相比,降了14.1 ng/g,在層積120 d時達(dá)到最低水平9.82 ng/g;而2,4-D處理的ABA含量在層積30 d時,急速下降,降了31.69 ng/g,在層積60 d時達(dá)到最低水平14.14 ng/g。表明2,4-D 5 mg/L處理的ABA含量下降速度的時間比CK早了30 d,最低水平時間比CK早60 d。
由圖3b可知,大百合種子在層積過程中,CK和2,4-D處理在IAA含量上呈先上升后下降的趨勢,不同的層積時間顯著極差異(P<0.01)。變溫層積0 d時,IAA含量最低,2,4-D處理比CK高出3.98 ng/g;在層積60 d時,IAA含量達(dá)到峰值,外源2,4-D抑制內(nèi)源IAA,CK高達(dá)1 954.52 ng/g,比2,4-D處理高出516.66 ng/g;層積120 d時,2,4-D處理的IAA含量上升較快,比 CK高出 173.82 ng/g。IAA含量在層積60 d時,上升速度較快,與30 d相比,CK增長了 1 877.49 ng/g,2,4-D處理增長了 947.2 ng/g,層積90 d時,IAA含量急速下降,與60 d相比,CK降低了1 925.39 ng/g,2,4-D處理下降了1 421.02 ng/g。
由圖3c可知,大百合種子在層積過程中,CK和2,4-D處理在GA3含量的水平變化大體上呈先上升后下降的趨勢。除層積130 d時差異顯著(P<0.05),其他的層積時間顯著極差異(P<0.01)。在變溫層積0 d時,2,4-D 5 mg/L處理的GA3含量比CK高出9.86 ng/g;GA3含量在層積120 d時達(dá)到峰值,2,4-D處理比CK高出52.17 ng/g,外源2,4-D促進(jìn)內(nèi)源GA3;層積120 d,GA3含量上升速度較快,與90 d相比,CK增長了38.4 ng/g,2,4-D處理增長了99.72 ng/g;層積130 d時,GA3含量急速下降,與120 d相比,CK降低了40.01 ng/g,2,4-D處理下降了96.34 ng/g。
2.4 大百合種子后熟過程中各激素含量之間的比值變化
由圖4可知,大百合種子后熟階段IAA/ABA、GA3/ABA之間的比值變化也存在差異,在層積過程中,IAA/ABA、GA3/ABA之間的比值整體上呈先上升后下降的趨勢。
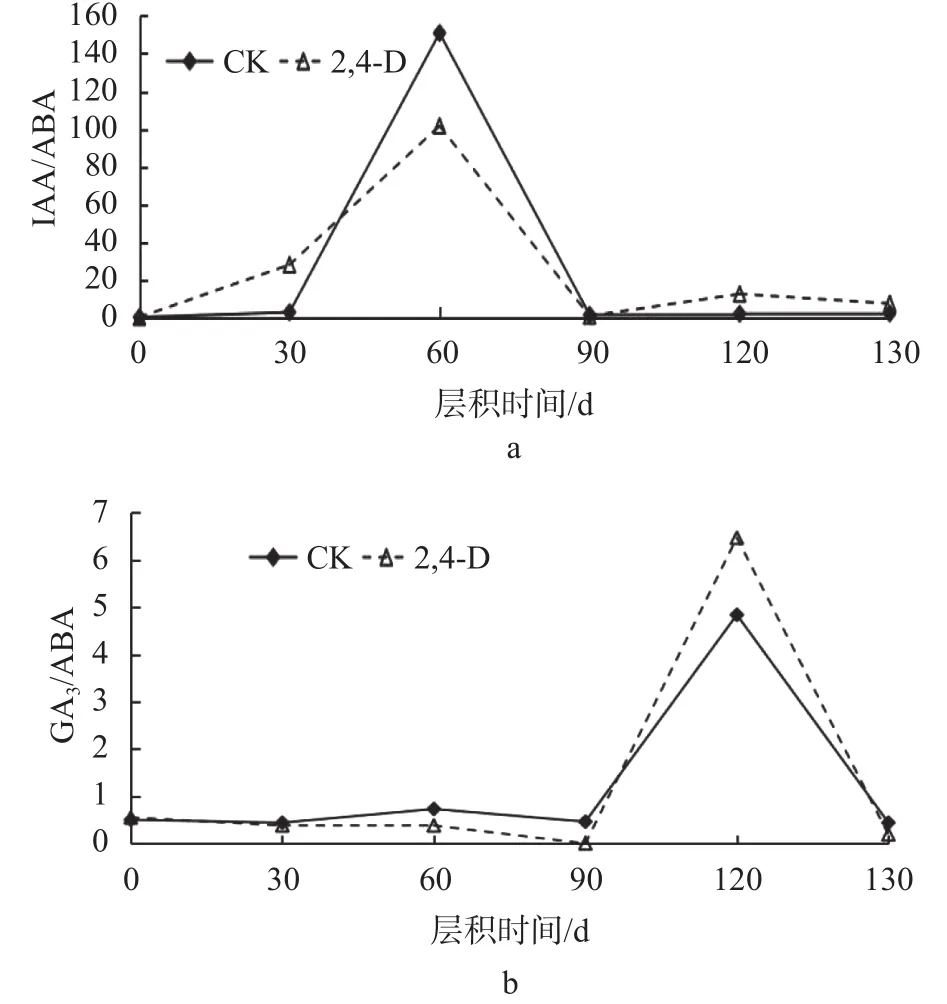
圖 4 大百合種子層積過程中各激素含量之間的比值變化Fig. 4 Changes in the ratio of hormones during stratification of C. giganteum seeds
從圖4a中可以看出,在層積60 d時,IAA/ABA達(dá)到峰值,CK比2,4-D處理高了49.24;IAA/ABA變化較大,在層積60 d時,上升速度較快,與30 d相比,CK的IAA/ABA增長了148.1,2,4-D處理增長了73.66;層積90 d時,IAA/ABA急速下降,與60 d相比,CK降低了149.47,2,4-D處理下降了100.84;層積120 d時,2,4-D處理的IAA/ABA比CK高出10.53。從圖4b中可以看出,在層積120 d時,GA3/ABA達(dá)到峰值,2,4-D處理比CK高了1.63;GA3/ABA變化較小,在層積120 d時,上升速度較快,與90 d相比,CK的GA3/ABA增長了4.38,2,4-D處理增長了6.47;層積130 d時,GA3/ABA急速下降,與120 d相比,CK降低了4.4,2,4-D處理下降了6.28。IAA/ABA、GA3/ABA上升說明發(fā)芽促進(jìn)物質(zhì)占相對優(yōu)勢,種子向萌發(fā)的方向發(fā)展。
3 結(jié)論與討論
種子休眠是植物經(jīng)過長期的演化而獲得的一種對環(huán)境條件及季節(jié)性變化的生物學(xué)適應(yīng)性[17]。近年來,國際上對形態(tài)生理休眠(MPD)種子的研究已取得了一定的進(jìn)展,但仍被認(rèn)為是種子生態(tài)學(xué)領(lǐng)域中了解最少的休眠類型之一。國內(nèi)對MPD的研究也才剛剛起步,已有的研究僅限于對部分植物的種子形態(tài)和萌發(fā)處理,研究還不深入、缺乏系統(tǒng)性。姚林君等[18]系統(tǒng)總結(jié)了MPD種子的形態(tài)結(jié)構(gòu)、休眠打破的環(huán)境需求、MPD的起源與進(jìn)化以及MPD的植物類群。形態(tài)生理休眠特性在木蘭科植物中非常常見,如西康玉蘭(Magnolia wilsonii)[19]。
馮小芹等[20]研究了98%濃硫酸、高溫處理、機械處理等6種不同種子處理方法對漆樹(Toxicdendro vernicifluum)幼苗生長的影響,本實驗采用變溫層積結(jié)合激素處理對大百合種子休眠解除的影響。張家玲等[21]用2,4-D對滇重樓(Paris polyphylla)種子萌發(fā)進(jìn)行研究,結(jié)果表明90 d時,CK的萌發(fā)率為40.33%,0.4 mg/L 2,4-D浸種處理48 h滇重樓種子萌發(fā)率最高達(dá)53.33%,0.2~0.4 mg/L的 2,4-D可促進(jìn)滇重樓種子的萌發(fā),而高于0.4 mg/L的2,4-D對種子的萌發(fā)具有抑制作用。大百合種子的休眠類型屬于MPD,本研究采用不同濃度2,4-D處理大百合種子,大百合種子的胚不完全伸長,種子也會萌發(fā),在層積135 d時,5 mg/L 2,4-D處理的胚長比CK高出9.03%,胚的長度不能衡量種子休眠解除的程度。5 mg/L 2,4-D處理的胚生長百分比最高,達(dá)到82%,發(fā)芽時間最早,為4 ℃下14 d;發(fā)芽率最高,達(dá)75.33%,5 mg/L 2,4-D處理可以縮短發(fā)芽時間,提高種子萌發(fā),利于大百合種胚后熟。1 mg/L 2,4-D處理促進(jìn)作用不明顯,10 mg/L 2,4-D處理起到明顯的抑制作用。
劉序等[22]采用GC-MS方法分析鑒定了香榧(Torreya grandis)種子的外種皮、內(nèi)種皮和胚乳萃取液中內(nèi)源物種類及其相對含量,本研究采用LC-MS方法測定了大百合種子層積過程中3種內(nèi)源激素。Khan[23]提出種子的休眠和萌發(fā)由3個因子調(diào)節(jié),即萌發(fā)促進(jìn)物質(zhì)GA3、細(xì)胞分裂素和ABA之間的相互作用決定種子休眠與萌發(fā)。ABA和GA3是調(diào)控種子休眠與萌發(fā)的一對拮抗內(nèi)源激素,IAA對萌發(fā)沒有明顯的影響,但間接影響種胚的發(fā)育和其他激素水平[24]。大百合種子層積過程中,ABA、IAA和GA3的含量發(fā)生不同規(guī)律的變化。5 mg/L 2,4-D處理的ABA含量在層積30 d時,急速下降,下降速度的時間比CK早了30 d,在層積60 d時達(dá)到最低水平,比CK早60 d。5 mg/L 2,4-D處理的IAA含量和GA3含量在層積的0 d、120 d時,均比CK高。內(nèi)源激素的相對水平對種子休眠具有重要的調(diào)控作用,IAA/ABA在層積60 d達(dá)到峰值,CK比5 mg/L 2,4-D高了49.24,GA3/ABA的比值在層積120 d達(dá)到峰值,5 mg/L 2,4-D處理比CK高了1.63。
大百合種子的休眠并不是單一因素引起的,而是各種激素共同作用的結(jié)果。綜合來看,ABA含量的變化與大百合種胚的發(fā)育呈負(fù)相關(guān),IAA和GA3可能是促進(jìn)大百合種子休眠解除的主要調(diào)節(jié)物質(zhì),5 mg/L 2,4-D處理通過抑制ABA而促進(jìn)種胚的萌發(fā)。不同外源GA3處理天女木蘭(Magnolia sieboldii)[25]胚乳中ABA含量隨層積時間大體上呈持續(xù)下降趨勢,GA3含量均在種子休眠解除期(120 d)含量達(dá)到最大,IAA含量呈先升高后下降的趨勢,并均在層積40 d時達(dá)到峰值,隨后逐漸下降,本研究中與天女木蘭胚乳中內(nèi)源激素含量的變化相似,表明適宜濃度的外源GA3和2,4-D浸種處理有利于種子休眠解除和萌發(fā)。
[ 參 考 文 獻(xiàn) ]
[1]中國科學(xué)院昆明植物研究所. 云南植物志: 第7卷 [M]. 北京: 科學(xué)出版社, 1997.
[2]馮國楣. 中國珍稀野生花卉[M]. 北京: 中國林業(yè)出版社, 1995.
[3]關(guān)文靈, 李世峰, 李葉芳, 等. 大百合鱗莖營養(yǎng)成分分析與評價 [J]. 西部林業(yè)科學(xué), 2011, 40(1): 8-11.
[4]萬珠珠, 龍春林, 程治英, 等. 重要野生花卉大百合屬植物研究進(jìn)展 [J]. 云南農(nóng)業(yè)大學(xué)學(xué)報, 2007, 22(1):30-34.
[5]裴盛基, 龍春林. 應(yīng)用民族植物學(xué)[M]. 昆明: 云南民族出版社, 2004.
[6]張金政, 龍雅宜, 孫國峰. 大百合的生物多樣性及其引種觀察 [J]. 園藝學(xué)報, 2002, 29(5): 462-466.
[7]李守麗, 石雷, 張金政, 等. 大百合子房的離體培養(yǎng) [J]. 園藝學(xué)報, 2007, 34(1): 197-200.
[8]關(guān)文靈, 李世峰, 雷麗萍, 等. 大百合小孢子發(fā)生和雄配子體發(fā)育研究 [J]. 亞熱帶植物科學(xué), 2011, 40(4):12-15.
[9]林鳳瓊, 朱昌叁, 黎素平, 等. 大百合與百合屬優(yōu)良品種雜交試驗 [J]. 南方農(nóng)業(yè)學(xué)報, 2012, 43(11): 1733-1736.
[10]Phartyal S S, Kondo T, Baskin C C, et al. Seed dormancy and germination in the giant Himalayan lily(Cardiocrinum giganteum var. giganteum): an assessment of its potential for naturalization in northern Japan [J]. Ecological Research, 2012, 27(4): 677-690.
[11]Cao G X, Worley A C. Life history trade-offs and evidence for hierarchical resource allocation in two monocarpic perennials [J]. Plant Biology, 2013, 15(1):158-165.
[12]Chen H N, Zhao C H, Liu X R, et al. Pollen development of Cardiocrinum giganteum (Wall.) Makina in China [J]. Plant Systematics and Evolution, 2012,298(8): 1557-1565.
[13]關(guān)文靈, 李世峰, 陳賢, 等. 大百合種子休眠特性及休眠破除 [J]. 西北植物學(xué)報, 2010, 30(12): 2479-2483.
[14]蔡薇, 李葉芳, 劉春雪, 等. 變溫層積下激素對大百合種胚后熟的影響 [J]. 中國農(nóng)學(xué)通報, 2017, 33(34):103-110.
[15]劉彩紅, 李成云, 樸光一, 等. 2,4-二氯苯氧乙酸對不同羊草種子發(fā)芽特性的影響 [J]. 東北農(nóng)業(yè)大學(xué)學(xué)報,2011, 42(12): 125-129.
[16]Wu J, Hettenhausen C, Meldau S, et al. Herbivory rapidly activates MAPK signaling in attacked and unattacked leaf regions but not between leaves of nicotiana attenuata [J]. The Plant Cell Online, 2007, 19(3):1096-1122.
[17]胡晉. 種子生物學(xué)[M]. 北京: 高等教育出版社, 2006.
[18]姚林君, 張克亮, 熊作明, 等. 種子形態(tài)生理休眠研究進(jìn)展 [J]. 生態(tài)學(xué)雜志, 2019, 38(1): 247-255.
[19]韓春艷, 龍春林. 瀕危植物西康玉蘭種子休眠、萌發(fā)及貯藏特性 [J]. 云南植物研究, 2010, 32(1): 47-52.
[20]馮小芹, 胡勐鴻, 呂尋. 種子不同處理方法對漆樹幼苗生長的影響 [J]. 西南林業(yè)大學(xué)學(xué)報(自然科學(xué)),2018, 38(6): 48-57.
[21]張家玲, 馬英姿, 胡文俐. 不同處理對滇重樓種子萌發(fā)的影響 [J]. 經(jīng)濟林研究, 2018, 36(2): 100-106.
[22]劉序, 陳黎, 高捍東. 香榧種子抑制其發(fā)芽和生長的內(nèi)源物篩選研究 [J]. 西南林業(yè)大學(xué)學(xué)報(自然科學(xué)),2018, 38(1): 28-33.
[23]Khan A A. Primary, preventive and permissive roles of hormones in plant systems [J]. The Botanical Review,1975, 41(4): 391-420.
[24]趙文琦, 季蘭. 種子休眠與解除機理的研究進(jìn)展 [J].山西農(nóng)業(yè)科學(xué), 2017, 45(3): 477-481.
[25]陸秀君, 梅梅, 劉月洋, 等. GA3和變溫層積對天女木蘭種子萌發(fā)及內(nèi)源激素的影響 [J]. 西北植物學(xué)報,2014, 34(9): 1828-1835.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語文(2016年21期)2016-05-25 13:13:50
