植物染色質重塑復合體研究進展
2019-08-27 04:27:26郭明欣何亞莉許可可趙旭升
安徽農業科學 2019年13期
郭明欣 何亞莉 許可可 趙旭升
摘要 染色質重塑復合體在基因的轉錄調控方面發揮重要作用。綜述了近年來國內外學者在染色質重塑復合體的組成、結構、分類及功能分析方面的最新研究概況。重點介紹了植物領域染色質重塑復合體核心亞基的研究進展,并分析了目前影響該領域發展的原因及解決對策,同時還對應用前景進行了展望。
關鍵詞 染色質重塑復合體;轉錄調控;核心亞基
中圖分類號 Q943文獻標識碼 A
文章編號 0517-6611(2019)13-0016-04
doi:10.3969/j.issn.0517-6611.2019.13.005
開放科學(資源服務)標識碼(OSID):
Research Advance on Chromatin Remodeling Complex in Plants
Abstract Chromatin remodeling complex play a vital role in transcriptional regulation. In this paper, we reviewed the components, structure, classification, functional analysis of chromatin remodeling complex. Furthermore, we focus on the core subunits of chromatin remodeling in plants. Moreover, the new trends and prospects of chromatin remodeling complex were also discussed.
Key words Chromatin remodeling complex;Transcriptional regulation;Core subunits
染色質的基本結構單元是核小體,核小體包括4種組蛋白即H2A、H2B、H3、H4,每一種組蛋白各有2個分子,形成為一個組蛋白八聚體。146 bp的DNA圍繞組蛋白八聚體形成核小體的核心結構。連接核小體之間的DNA鏈上由組蛋白H1 結合,由此形成了11 nm的核小體串珠狀結構。核小體經過進一步的折疊及組裝形成染色質的高級結構[1]。染色質的折疊和包裝使染色質可以合適的大小存在于真核細胞核里。然而,染色質的高度緊密使得一些基因的正常轉錄受到抑制。現在已知有2類染色質的修飾復合物:一類是ATP依賴的染色質重塑復合體(ATP-dependent chromatin remodeling complex),其依靠核心亞基ATPase水解ATP為整個復合物提供能量。該復合體可以移動核小體的位置,改變DNA和組蛋白之間的互作,使轉錄起始因子可以結合在DNA上[2-4]。另一類是染色質的共價化學修飾,一般是對組蛋白末端“尾巴”進行乙酰化、磷酸化、甲基化和泛素化等修飾[5]。已有的研究表明,染色質重塑復合體通過調控相關基因的表達參與了植物的生長發育以及對外界逆境響應等眾多過程。目前對于染色質重塑復合體的研究已逐漸成為研究基因表達調控的熱點。基于此,筆者綜述了染色質重塑復合體的組成、結構、分類以及功能,為后續的研究提供參考,并在核心亞基的功能解析方面進行了展望。
1 染色質重塑復合體的組成和結構
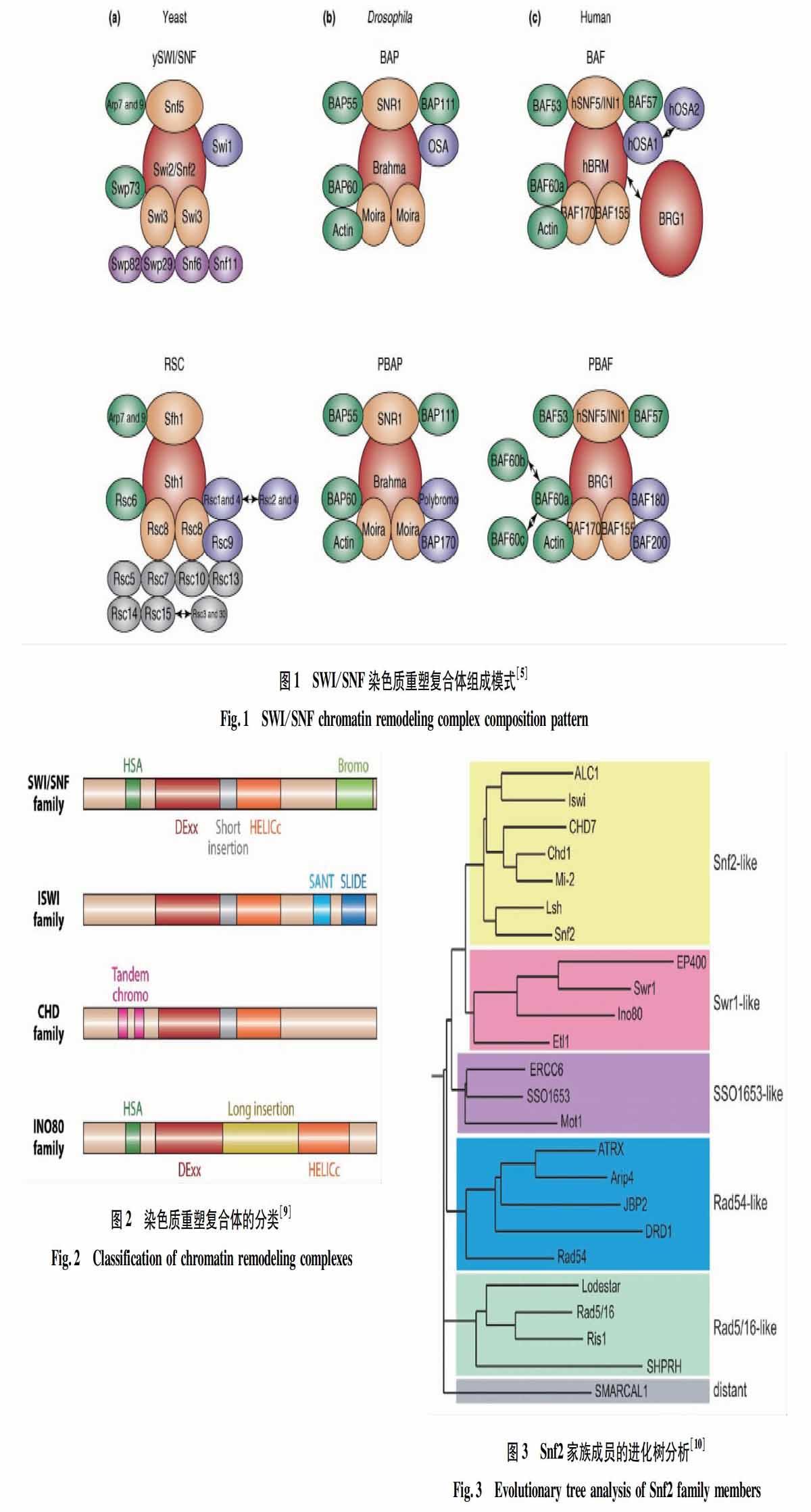
ATP 依賴的染色質重塑復合體是由多個亞基組成的復合體。核心亞基是屬于 Snf2家族的ATPase。由于復合體有很多的亞基組成,而且有些亞基分子量很小。另外,有些核心亞基,比如酵母的 Sth1、Isw1 等參與了幾個結構相似、功能不同的重塑復合體[6-7],因此直接純化染色質重塑復合體變得非常困難,也阻礙了對該類復合體功能的深入研究。目前,利用低分辨率的電子顯微鏡(low resolution electron microscopy)已經確定了少數染色質重塑復合體的組成。如圖1所示[5],復合體的結構像一個大的碗,碗的中央部位收縮以使復合體能夠結合在核小體上[8]。
2 染色質重塑復合體的分類
由于很多染色質重塑復合體的組成和生化功能是未知的,因此很難根據生化功能對其進行分類。傳統的分類方法,是根據Snf2蛋白所包含的2個主要功能域DEXDc和HELICc之間插入氨基酸的長度,把染色質重塑復合體分為4類,包括SWI/SNF、 Mi-2/CHD、ISWI、INO80/SWR1(圖2)[9]。這是一種過于簡化的、比較模糊的分類方法。
Snf2家族的蛋白都含有DEXDc和HELICc這2個結構域(Domain),這2個結構域組合在一起發揮ATPase的作用。Flaus等[10]利用來自不同物種的1 300個Snf2家族的蛋白,通過對蛋白的DEXDc和HELICc所包含的及2個功能域之間的氨基酸序列進行多重序列比對建立了進化樹,把Snf2家族分為5個類群(group)、23 個亞家族(subfamily)(圖3)。
3 染色質重塑復合體的功能
目前對染色質重塑復合體的功能研究主要集中在動物和微生物領域,比如人類、斑馬魚、果蠅、小鼠、或線蟲。在模式植物擬南芥、水稻等有少量的相關研究報道,但總體研究進展較其他領域遠為落后。
染色質重塑復合體在生物體中的功能主要體現在3個方面。第一,調節基因的轉錄,起始或抑制基因的轉錄。大部分ATP依賴的染色質重塑復合體能夠改變染色質結構激活基因的轉錄[11-14],但是Mi-2/NuRD亞家族的染色質重塑復合體卻抑制基因的轉錄。Mi-2/NuRD不僅具有改變核小體位置的功能,還具有組蛋白去乙酰化的功能,使處于轉錄激活狀態的基因中的組蛋白發生去乙酰化,從而導致轉錄受到抑制[15]。第二,染色質重塑復合體參與DNA修復和減數分裂時姐妹染色體的聯會和分離。酵母的RSC、SWI/SNF 和擬南芥的BRM參與了DNA的損傷修復[16-17];酵母的RSC參與了減數分裂時姐妹染色體的聯會和分離[18]。第三,當生物面對內在或外在條件改變時,需要染色質重塑復合體來完成轉錄重組(transcriptional reprogramming)。
酵母中的SWI/SNF是第一個被發現的染色質重塑復合體,其核心亞基Swi2/Snf2的相關突變體是通過2個獨立的遺傳途徑篩選到的。swi2突變體表現交配缺陷的表型(mating type switching defective phenotypes);而snf2突變體表現蔗糖不發酵的表型(sucrose nonfermenting phenotypes),2個突變體是酵母中同一個基因 YOR290C突變所致。后續研究表明,Swi2/Snf2與其他9個亞基組成一個大的復合體發揮作用(圖1)[5]。之后,又陸續在酵母、人類、果蠅等生物體中發現了一些染色質重塑復合體,如酵母的RSC、ISW1a、ISW1b、ISW2、CHD1、INO80、SWR1[10,19-20];人類的BAF、PBAF、NURF、CHRAC、ACF、CHD1、NuRD、INO80、SRCAP、TRRAP[10];果蠅的BAP、PBAP、NURF、CHRAC、ACF、CHD1、Mi-2、Tip60、Pho-dINO80[10]。這些染色質重塑復合體參與了生物體的很多過程,包括交配、合子分裂及胚胎的早期發育和器官的發生(神經系統、心臟系統、表皮系統、肌肉系統等[5,9]。
在植物中,到目前為止還沒有一個完整的染色質重塑復合體的組成被發現。但是有一些核心亞基的功能已經被研究,比如擬南芥的AtBRM[21-24]、AtSYD[25-28]、AtCHR12[29-30]、AtCHR23[29],以及水稻的OsCHR4[31]、OsALT1[32]。這些核心亞基參與了植物的很多發育過程,包括營養生長、生殖生長以及植物對生物和非生物脅迫的響應。
3.1 AtBRM
AtBRM是果蠅DmBRM的同源基因。DmBRM是Snf2類型的染色質重塑復合體BAP、PBAP的核心亞基。AtBRM編碼的蛋白由 2 193個氨基酸組成,含有 DEXDc、HELICc、BROMO 3個功能域。對AtBRM的功能研究表明,AtBRM調控著擬南芥的營養生長、生殖生長及對非生物脅迫的響應。AtBRM的RNAi轉基因植株表現為株高變矮、葉片卷曲、葉片與花序分生組織變小、開花期提前,并導致了花器官形態的改變如花瓣和雄蕊變小、花藥不能成熟、結實率降低[21]。brm突變體則表現了更嚴重的發育缺陷,純合突變體完全不育[22]。
合成和積累種子儲藏蛋白(seed storage proteins)是種子成熟過程中的一個重要方面。編碼種子儲藏蛋白相關的基因主要在成熟的種子中表達,而在營養器官如葉片中沒有轉錄產物。但是,在brm突變體的葉片中,能夠檢測到與種子儲藏蛋白相關基因如At2S2、At2S3、At2S5、At7S1等的表達[23]。進一步研究表明,AtBRM與AtSWI3C呈現較強互作,與AtSWI3B微弱互作,而與AtSWI3A和AtSWI3D沒有互作[22]。而且,swi3c突變體與brm有相似的表型,而且在swi3c突變體葉片中At2S2、At2S3、At7S1等基因的表達明顯上調。SWI/SNF復合體的組成亞基Snf5在擬南芥中的只有一個同源基因AtBSH,bsh突變體與brm有相似的表型,At7S1的表達在bsh突變體葉片中也明顯上調[23]。另外,brm、swi3c、bsh突變體在苗期對植物激素ABA都表現高度敏感,其中AtBRM的功能喪失導致AtABI5的表達上調,使brm突變體的耐旱能力明顯增強[24]。
3.2 AtSYD
AtSYD 是Snf2類型的染色質重塑復合體的核心亞基,編碼的蛋白由3 583個氨基酸組成,含有DEXDc和HELICc 2個結構域。對AtSYD的功能研究表明,AtSYD 調控著擬南芥全生育期的生長發育以及對生物脅迫的響應,該基因的功能喪失造成syd突變體株高變矮、生長緩慢、葉片極性缺失、次級花序減少。同時,AtSYD與AtLFY共同調控莖端分生組織(shoot apical meristem,SAM)的識別[25],并通過直接調控WUS的表達而影響頂端分生組織的大小[26]。
植物花器官的發育由A、B、C、D、E這5類基因控制,其中B、C類基因控制著雄性和雌性生殖器官的發育,因而顯得尤為重要。在擬南芥中存在一類多梳蛋白,它們會抑制B、C類基因的表達。最近的研究表明,AtSYD和AtBRM能夠克服多梳蛋白的抑制而激活B類基因AP3和C類基因AG的表達,而且AtBRM和AtSYD可以直接分別結合在AP3和AG的啟動子區域[27]。
另外,AtSYD 還參與了生物脅迫的信號轉導。AtSYD調控乙烯(ethylene)和茉莉酸(jasmonic acid,JA)信號通路中一些基因如PDF1.2a、MYC2、VSP2的表達。在syd突變體中,PDF1.2a、MYC2、VSP2的表達明顯下調。與野生型相比,syd 對真菌病原物番茄灰霉病菌(Botrytis cinerea)敏感,但是對細菌病原物假單胞桿菌(Pseudomonas syringae)表現不敏感,表明AtSYD對病原菌具有選擇抗性[28]。
3.3 AtCHR12、AtCHR23
AtCHR12、AtCHR23又分別叫做MINU1、MINU2。AtCHR12編碼的蛋白由1 102 個氨基酸組成,而AtCHR23編碼的蛋白由1 064個氨基酸組成。AtCHR12與AtCHR23都含有DEXDc和HELICc這2個結構域,具有81.7% 的同源性。由此,AtCHR12和AtCHR23表現功能冗余,單個基因功能喪失的chr12或chr23突變體在表型上沒有明顯變化。與此相比,chr12、chr23雙突變體則表現為株高變矮、根系變短、生長緩慢、生育期延長、花藥數目減少、結實率降低[29]。
AtCHR12負向調控擬南芥對非生物脅迫的耐性。在正常生長條件下,chr12突變體的表型與野生型沒有差異,但在干旱、高溫、鹽脅迫的情況下,突變體則表現了明顯的耐性。在干旱和高溫脅迫的情況下,chr12的株高高于野生型,而過量表達該基因的轉基因植株的株高則較野生型降低。另外,在25、50 mM NaCl脅迫時,chr12突變體的根長較野生型明顯增加[30]。
3.4 OsCHR4
OsCHR4屬于Mi-2亞家族的核心亞基,編碼的蛋白由2 259個氨基酸組成,含有1個PHD功能域、2個CHROMO功能域、1個DEXDc功能域和1個HELICc功能域。OsCHR4在水稻近軸段葉肉細胞葉綠體的發育中發揮重要作用。chr4突變體葉片的遠軸端生長正常,而近軸端葉色變黃。進一步的研究表明,chr4突變體葉片近軸端葉肉細胞的葉綠體存在缺陷,其質體的生長和類囊體膜的形成被阻礙。OsCHR4定位在細胞核內,其轉錄產物主要分布在根端分生組織、花、維管束以及葉肉細胞中[31]。
3.5 OsALT1
OsALT1屬于推測的Snf2家族的核心亞基,編碼的蛋白由1 228個氨基酸組成,含有1個DEXDc功能域、1個HELICc功能域和1個RING finger結構域。OsALT1在水稻苗期的耐堿性及生長發育方面發揮重要的調節作用。alt1突變體在苗期表現非常明顯的耐堿性,同時,在正常栽培條件下,alt1突變體的最典型特征是根系較短,同時地上部有一定程度的矮化。
系統進化分析結果表明,ALT1屬于Snf2家族中Ris1亞家族。亞細胞定位顯示ALT1是一個核定位蛋白。GUS染色和定量RT-PCR結果顯示,ALT1在根、莖、葉片、葉鞘、幼穗中均有表達。與ALT1的耐堿負向調節功能相一致,ALT1的表達受到堿脅迫顯著抑制。對ALT1耐堿機理的研究表明,ALT1主要通過調控氧化脅迫的損傷防御在水稻耐堿中發揮作用,揭示了植物耐堿的一個新的分子機制[32]。
4 展望
經過10多年的研究,生物中染色質重塑復合體的研究取得了較大的進展,部分染色質重塑復合體的結構、組成及核心亞基和附屬亞基的功能被解析,其作用機制也越來越清楚。然而在植物領域,染色質重塑復合體的研究卻相對緩慢,以模式植物擬南芥和水稻為例,擬南芥基因組中有41個Snf2家族的成員,目前僅有4個成員的功能被解析;水稻基因組中有42個Snf2家族的成員,目前僅有2個成員的功能被解析。導致這種現象的原因,一方面可能是Snf2家族基因內部存在功能冗余的現象,比如擬南芥AtCHR12和AtCHR23,因此不太容易篩選到表型發生明顯改變的突變體;另一方面,Snf2家族成員編碼的蛋白分子量均較大(大部分基因編碼的蛋白長度在1 000 aa以上),這在一定程度上限制了通過反向遺傳學的手段研究基因的功能。
近年來,基因組編輯(Genome editing)技術的出現,特別是CRISPR/Cas9技術在基因敲除中的廣泛應用,為研究Snf2家族的基因功能提供了一個非常好的途徑。CRISPR/Cas9實現了同時對多個基因的敲除[33],對于研究存在功能冗余的基因家族非常有用。此外,隨著部分Snf2家族耐逆功能的解析[32,34],應用這些基因進行分子改良與新品種選育,將會是一個很好的研究方向。
參考文獻
[1] TREMETHICK D J.Higherorder structures of chromatin:The elusive 30 nm fiber[J].Cell,2007,128(4):651-654.
[2]? HAVAS K,WHITEHOUSE I,OWENHUGHES T.ATPdependent chromatin remodeling activities[J].Cellular and molecular life sciences,2001,58(5/6):673-682.
[3]? HARGREAVES D C,CRABTREE G R.ATPdependent chromatin remodeling:Genetics,genomics and mechanisms[J].Cell research,2011,21(3):396-420.
[4]? CLAPIER C R,CAIRNS B R.The biology of chromatin remodeling complexes[J].Annual review of biochemistry,2009,78:273-304.
[5]? KWON C S,WAGNER D.Unwinding chromatin for development and growth:A few genes at a time[J].Trends genetics,2007,23(8):403-412.
[6]? CAIRNS B R,SCHLICHTER A,ERDJUMENTBROMAGE? H,et al.Two functionally distinct forms of the RSC nucleosomeremodeling complex,containing essential AT hook,BAH and bromodomains[J].Molecular cell,1999,4(5):715-723.
[7]? VARY J C,GANGARAJU V K,QIN J,et al.Yeast Isw1p forms two separable complexes in vivo[J].Molecular cellular biology,2003,23(1):80-91.
[8]? HOTA S K,BARTHOLOMEW B.Diversity of operation in ATPdependent chromatin remodelers[J].Biochimica biophysica acta,2011,1809(9):476-487.
[9]? CLAPIER C R,CAIRNS B R.The biology of chromatin remodeling complexes[J].Annual review of biochemistry,2009,78:273-304.
[10]? FLAUS A,MARTIN D M A,BARTON G J,et al.Identification of multiple distinct Snf2 subfamilies with conserved structural motifs[J].Nucleic acids research,2006,34(10):2887-2905.
[11]? NARLIKAR G J,FAN H Y,KINGSTON R E.Cooperation between complexes that regulate chromatin structure and transcription[J].Cell,2002,108:475-487.
[12] MARTENS J A,WINSTON F.Recent advances in understanding chromatin remodeling by Swi/Snf complexes[J].Current opinion in genetics & development,2003,13:136-142.
[13]? HSIAO P W,FRYER C J,TROTTER K W,et al.BAF60a mediates critical interactions between nuclear receptors and the BRG1 chromatinremodeling complex for transactivation[J].Molecular and cellular biology,2003,23(17):6210-6220.
[14]? PRECHT P,WURSTER A L,PAZIN M J.The SNF2H chromatin remodeling enzyme has opposing effects on cytokine gene expression[J].Molecular immunology,2010,47(11/12):2038-2046.
[15]? DENSLOW S A,WADE P A.The human Mi2/NuRD complex and gene regulation[J].Oncogene,2007,26:5433-5438.
[16]? CHAI B,HUANG J,CAIRNS B R,et al.Distinct roles for the RSC and Swi/Snf ATPdependent chromatin remodelers in DNA doublestrand break repair[J].Genes development,2005,19(14):1656-1661.
[17]? SHAKED H,AVIVIRAGOLSKY N,LEVY A A.Involvement of the Arabidopsis SWI2/SNF2 chromatin remodeling gene family in DNA damage response and? recombination[J].Genetics,2006,173(2):985-994.
[18] HUANG J,HSU J M,LAURENT B C.The RSC nucleosomeremodeling complex is required for Cohesin's association with chromosome arms[J].Molcular cell,2004,13(5):739-750.
[19] VARY J C,JR,GANGARAJU V K,QIN J,et al.Yeast Isw1p forms two separable complexes in vivo[J].Molecular cellular biology,2003,23(1):80-91.
[20] JIANG Y J,WANG X,BAO S L,et al.INO80 chromatin remodeling complex promotes the removal of UV lesions by the nucleotide excision repair pathway[J].Proceedings of the national academy of sciences of the USA,2010,107(40):17274-17279.
[21]? FARRONA S,HURTADO L,BOWMAN J L,et al.The Arabidopsis thaliana SNF2 homolog AtBRM controls shoot development and flowering[J].Development,2004,131(20):4965-4975.
[22]? HURTADO L,FARRONA S,REYES J C.The putative SWI/SNF complex subunit BRAHMA activates flower homeotic genes in Arabidopsis thaliana[J].Plant molcular biology,2006,62(1/2):291-304.
[23]? TANG X R,HOU A F,BABU M,et al.The Arabidopsis BRAHMA chromatinremodeling ATPase is involved in repression of seed maturation genes in leaves[J].Plant physiology,2008,147:1143-1157.
[24]? HAN S K,SANG Y,RODRIGUES A,et al.The SWI2/SNF2 chromatin remodeling ATPase BRAHMA represses abscisic acid responses in the absence of the stress stimulus in Arabidopsis[J].Plant cell,2012,24(12):4892-4906.
[25]? WAGNER D,MEYEROWITZ E M.SPLAYED,a novel SWI/SNF ATPase homolog,controls reproductive development in Arabidopsis[J].Current biology,2002,12(2):85-94.
[26]? KWON C S,CHEN C,WAGNER D.WUSCHEL is a primary target for transcriptional regulation by SPLAYED in dynamic control of stem cell fate in Arabidopsis[J].Genes development,2005,19(8):992-1003.
[27]? WU? M F,SANG Y,BEZHANI S,et al.SWI2/SNF2 chromatin remodeling ATPases overcome polycomb repression and control floral organ identity with the LEAFY and SEPALLATA3 transcription factors[J].Proceedings of the national academy of sciences of the USA,2012,109(9):3576-3581.
[28]? WALLEY J W,ROWE H C,XIAO Y M,et al.The chromatin remodeler SPLAYED regulates specific stress signaling pathways[J].PLoS Pathogens,2008,4(2):1-8.
[29]? SANG Y,SILVAORTEGA C O,WU S,et al.Mutations in two noncanonical Arabidopsis SWI2/SNF2 chromatin remodeling ATPases cause embryogenesis and stem cell maintenance defects[J].The plant journal,2012,72(6):1000-1004.
[30]? MLYNROV L,NAP J P,BISSELING T.The SWI/SNF chromatinremodeling gene AtCHR12 mediates temporary growth arrest in Arabidopsis thaliana upon perceiving environmental stress[J].The plant journal,2007,51(5):874-885.
[31]? ZHAO C F,XU J M,CHEN Y,et al.Molecular cloning and characterization of OsCHR4,a rice chromatinremodeling factor required for early chloroplast development in adaxial mesophyll[J].Planta,2012,236(4):1165-1176.
[33]? GUO M X,WANG R C,WANG J,et al.ALT1,a Snf2 family chromatin remodeling ATPase,negatively regulates alkaline tolerance through enhanced defense against oxidative stress in rice[J].PLoS One,2014,9(12):1-25.
[33]? ZHANG Z J,MAO Y F,HA S,et al.A multiplex CRISPR/Cas9 platform for fast and efficient editing of multiple genes in Arabidopsis[J].Plant cell reports,2015,35:1519-1533.
[34]? KIM J M,TO T K,NISHIOKA T,et al.Chromatin regulation functions in plant abiotic stress responses[J].Plant,cell and environment,2010,33:604-611.
