撫育擇伐對馬尾松林主要樹種空間分布格局及其關聯性的短期影響
2019-08-20 10:58:02王曉榮曾立雄肖文發
生態學報 2019年12期
王曉榮,曾立雄,雷 蕾,潘 磊,肖文發,*
1 中國林業科學研究院森林生態環境與保護研究所,國家林業局森林環境重點實驗室, 北京 100091 2 湖北省林業科學研究院, 武漢 430075 3 南京林業大學南方現代林業協同創新中心, 南京 210037
群落結構由植物種類組成及其空間分布格局所決定,物種間不同的空間關系會導致群落結構存在差異[1],直接影響不同層次樹種的空間分布格局和更新演替機制[2]。關于植物種群空間結構的研究多集中在未受干擾的自然林分,重點研究自然群落結構形成和動態變化[3]、組配方式[4- 5]、物種共存或競爭[6- 7]、生物多樣性維持等[8- 9],但對人為采伐干擾下的群落空間結構變化和種間相關性等方面研究明顯不足[10- 11]。
撫育擇伐作為森林經營中重要的人為干擾方式,往往會改變森林的物種組成和群落空間結構[12],造成林內溫度、光照、物種競爭、更新狀況、特定生境條件和土壤等生態系統結構要素的改變[13- 14]。已有研究發現,適度擇伐可促進森林結構優化,對植物群落生長和更新演替產生有利影響[12,15],但以往經營管理更多關注擇伐密度、擇伐強度、次數以及采伐周期,對采伐物種類型、種間關系等林分結構的改變考慮較少[10- 11,16]。同時,不同擇伐方式對森林結構的影響會隨研究區域或樹種差異而不同[17],使得撫育擇伐方式對森林結構變化的影響一直未有定論[14]。因此,明確典型森林群落物種空間分布格局和種間關系對撫育擇伐干擾的響應,有助于揭示人為經營干擾對森林群落特性、穩定性和演替規律等的影響機制[4,18-19],為合理開展森林經營管理提供科學依據。
目前,我國人工林面積全球最大,根據第八次全國森林資源清查,面積已達0.693億hm2[20]。馬尾松(Pinusmassoniana)是我國亞熱帶特有鄉土樹種,適應性強、耐干旱與瘠薄,廣泛分布于秦嶺、淮河以南[21],也是三峽庫區分布面積最大和最主要的林分類型,占庫區森林面積34%[22]。以往馬尾松林主要利用方式和管理措施包括采伐大徑材馬尾松、砍伐雜木以及林下植被撫育等3種模式,長期受經營水平粗放和人為干擾強烈等多種因素的影響,庫區馬尾松林分質量、穩定性及生態防護功能低下等問題十分突出。監測常規撫育擇伐措施下馬尾松林分結構組成特征及種間功能聯系,追蹤撫育擇伐后森林空間格局形成的生態學過程,以確定合理的撫育擇伐策略,可促進林分結構優化發展。因此,本研究以三峽庫區馬尾松林為研究對象,模擬以往馬尾松利用和管理措施,采用未擇伐、伐除優勢馬尾松、伐除非馬尾松、除灌等方式[23]建立固定監測樣地,利用點格局法分析撫育擇伐3年后馬尾松林中主要樹種結構、空間分布和種間關聯性,探討不同撫育擇伐方式對馬尾松林分組成和結構的短期影響,為三峽庫區馬尾松森林結構優化和林分質量提升以及制定合理的森林經營方案提供參考。
1 研究區概況
研究地點位于湖北省宜昌市秭歸縣九嶺頭林場(30°59′—31°05′N,110°45′—110°48′E),海拔156—2030.6 m,森林覆蓋率95.2%。屬于亞熱帶大陸性季風氣候,年均氣溫17.9℃,年降水量1000—1250 mm,多集中在4—9月份,年日照時數1619.6 h,無霜天260 d。土壤類型主要以黃壤、黃棕壤為主。研究區內馬尾松林為20世紀70年代飛播造林,長期以來人為干擾極少,經過自然更新演替,形成以馬尾松為優勢樹種,伴生有少量光皮樺(Betulaluminifera)、漆樹(Toxicodendronvernicifluum)、茅栗(Castaneaseguinii)、鹽膚木(Rhuschinensis)等的松闊混交林。灌木樹種主要有綠葉胡枝子(Lespedezabuergeri)、樺葉莢蒾(Viburnumbetulifolium)、宜昌莢蒾(Viburnumerosum)等,草本以中日金星蕨(Parathelypterisnipponica)、狗脊(Cibotiumbarometz)、魚腥草(HeartleafHouttuynia)、野青茅(Deyeuxiaarundinacea)、細梗苔草(Carexteinogyna)為主。
2 研究方法
2.1 樣地設置
2013年9月,選擇生長在同一坡面且林相完整的馬尾松林,按照未擇伐(Non-cutting, NC)、伐除優勢馬尾松(DominantPinusmassoniana-cutting, DPMC)、伐除非馬尾松(NoPinusmassoniana-cutting, NPMC)、除灌(Shrubs-cutting, SC)等4種管理措施,各設置1塊面積為50 m×50 m的正方形樣地,樣地間相距20—30 m(當樣地面積≥2500 m2時能夠較真實的反映林木的空間分布格局[24])。伐除優勢馬尾松措施為采伐樣地內平均胸徑以上的馬尾松,采伐強度為70%(按胸高斷面積計算);伐除非馬尾松措施為采伐樣地內胸徑在4 cm以上的非馬尾松(包括光皮樺、漆樹等喬木,以及部分木姜子、火棘、樺葉莢蒾等),強度為15%;除灌措施為清除樣地內林下灌木層,但保留更新馬尾松小樹。采伐作業采用人力油鋸伐木,對采伐的樹干進行移除[23]。
2.2 調查方法
于2016年8月,利用相鄰格子調查方法,將樣地劃分為25個10 m×10 m的樣方,對各樣地內對所有胸徑≥1 cm的喬木進行編號、掛牌并進行每木檢尺,記錄樣方號、樹種、胸徑。以每個樣方為單元,左下角為坐標原點測量所有胸徑≥1 cm的喬木樹種相對于坐標原點的平面直角坐標系的相對坐標(以X,Y的形式表示)。同時,測定各樣地的經緯度、海拔、坡度、坡向等立地因子。樣地基本情況具體見表1。
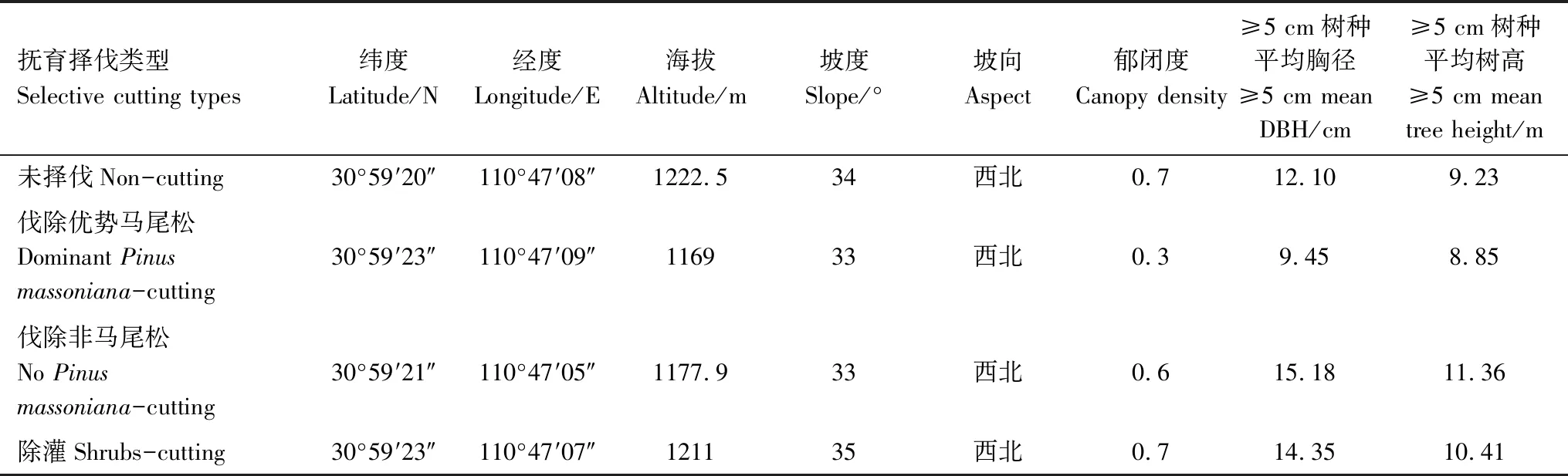
表1 擇伐撫育3 a后樣地基本概況
2.3 數據分析
2.3.1 重要值計算方法
重要值=(相對密度+相對優勢度+相對頻度)/3
相對密度=(某個種的個體數/所有種的個體總數)×100
相對優勢度=(某個種的胸高斷面積和/所有種個體胸高斷面總和)×100
相對頻度=(某個種的頻度/所有種頻度總和)×100
2.3.2 物種多樣性測度
Margalef豐富度指數(dM)
dM=(S-1)/lnN
Simpson優勢度指數(λ)λ=∑ni(ni-1)/N(N-1)
Pielou均勻度指數(Jsh)Jsh=H/lnS
式中,S為物種數;N為所有物種的個體數之和;Pi=ni/N代表第i個物種的相對多度,ni為第i個物種的個體數。
2.3.3 喬木個體空間點格局分析
首先,將每個樣方實測坐標進行全局化處理后得到林木個體坐標。然后,采用Ripley′s K函數的改進型Ripley′s L函數[3,10]分析不同擇伐撫育措施主要樹種的空間分布格局與種間空間關聯性。具體公式如下:
式中,A為研究區面積;uij為點i與j之間的距離;Ir(uij)為指示函數,當uij≤r時,Ir(uij)=1,當uij>r時,Ir(uij)=0;wij為權重值,用于邊緣校正。
式中,L(r)=0時,表示隨機分布;L(r)>0時,表示聚集分布;L(r)<0時,表示均勻分布。
利用Monte-Carlo函數來擬合檢驗計算上下包跡線即置信區間,并以空間尺度r為橫坐標,L(r)實際值和擬合值的上下包跡線為縱坐標作圖。當分析單個種群時,若種群實測值在包跡線以內,則符合隨機分布;若在包跡線上,則為集群分布;若在包跡線下,則為均勻分布。當分析兩個種群間關聯性時,種群實際分布數據L12(r)高于上包跡線,表示顯著正相關,在上下包跡線之間,表示相關性不顯著,低于下包跡線則為顯著負相關。
本研究隨機模擬1000次,達到99%的置信水平,最大距離尺度為12.5 m,為樣地邊長的1/4[14]。數據分析與作圖利用R語言軟件R- 3.2.4中相關程序包以及Excel 13.0完成。
3 結果與分析
3.1 撫育擇伐對馬尾松林物種組成特征及物種多樣性的影響
撫育擇伐3 a后,NC樣地中喬木物種22種,DPMC 17種,NPMC 21種,SC17種。進一步統計出各措施下重要值≥5的喬木樹種優勢程度(表2),NC樣地中優勢樹種包括馬尾松、茅栗、光皮樺、漆樹、宜昌木姜子、鹽膚木,累積重要值占總體的82.34%;DPMC樣地優勢樹種包括馬尾松、香椿、光皮樺、漆樹、鹽膚木、宜昌木姜子、檫木、火棘,累積重要值占總體的90.14%;NPMC樣地優勢樹種包括馬尾松、漆樹、宜昌木姜子、鹽膚木、香椿、火棘、檫木,累積重要值占總體的87.23%;SC樣地優勢樹種包括馬尾松、漆樹、光皮樺、香椿,累積重要值占總體的73.3%。不同撫育擇伐措施均在一定程度上降低了喬木層物種多樣性(表3),與NC相比,DPMC、NPMC和SC的Margalef豐富度指數和Shannon-Wiener多樣性指數分別降低28.43%、6.71%、12.79%和5.22%、2.17%、7.39%,Simpson優勢度指數具有輕微增加,Pielou均勻度指數則無顯著變化。不同撫育擇伐措施均在一定程度上改變了物種組成以及部分樹種的優勢程度,以DPMC和SC對林分物種組成影響最大。
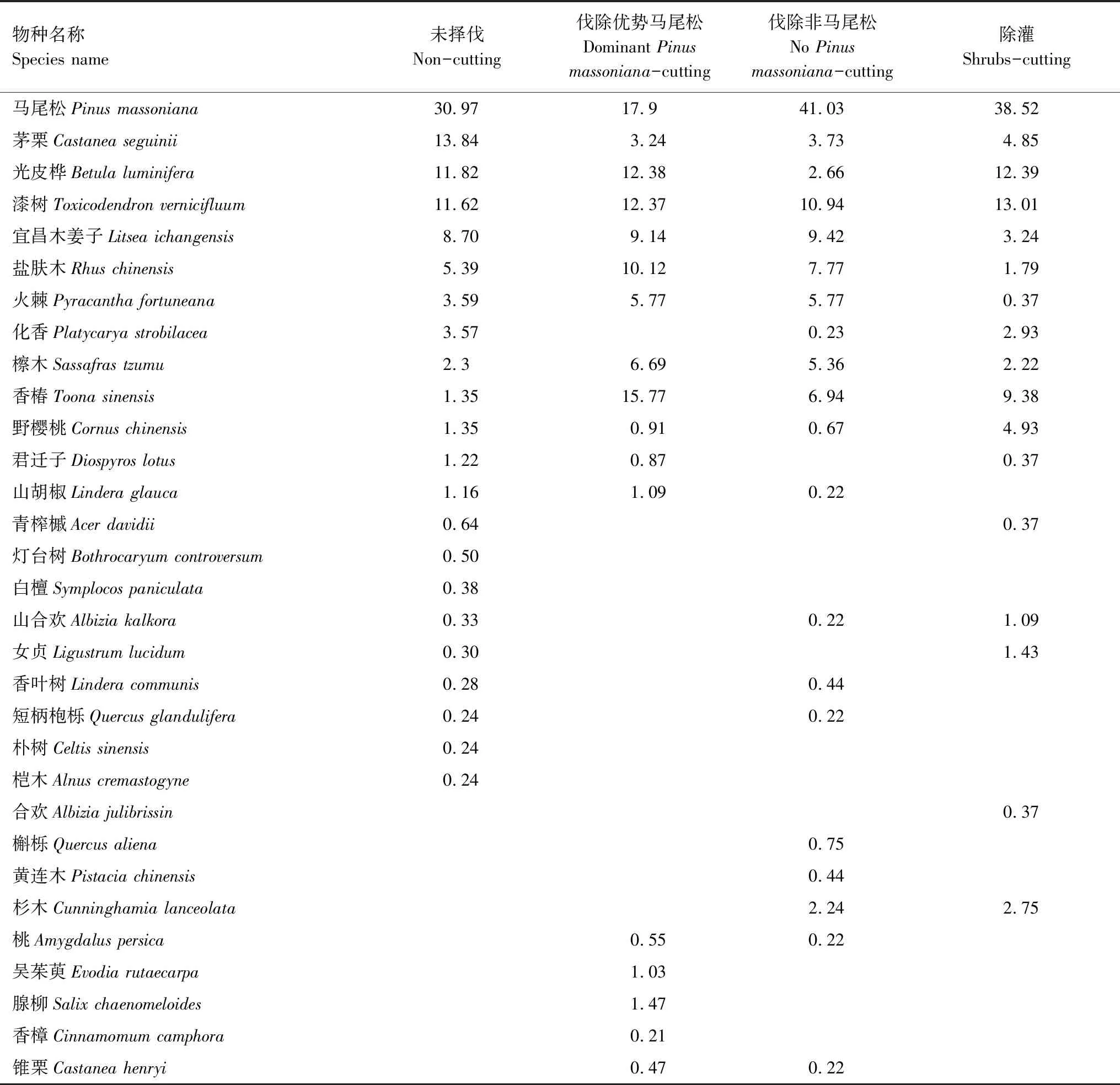
表2 各撫育擇伐措施中主要樹種重要值
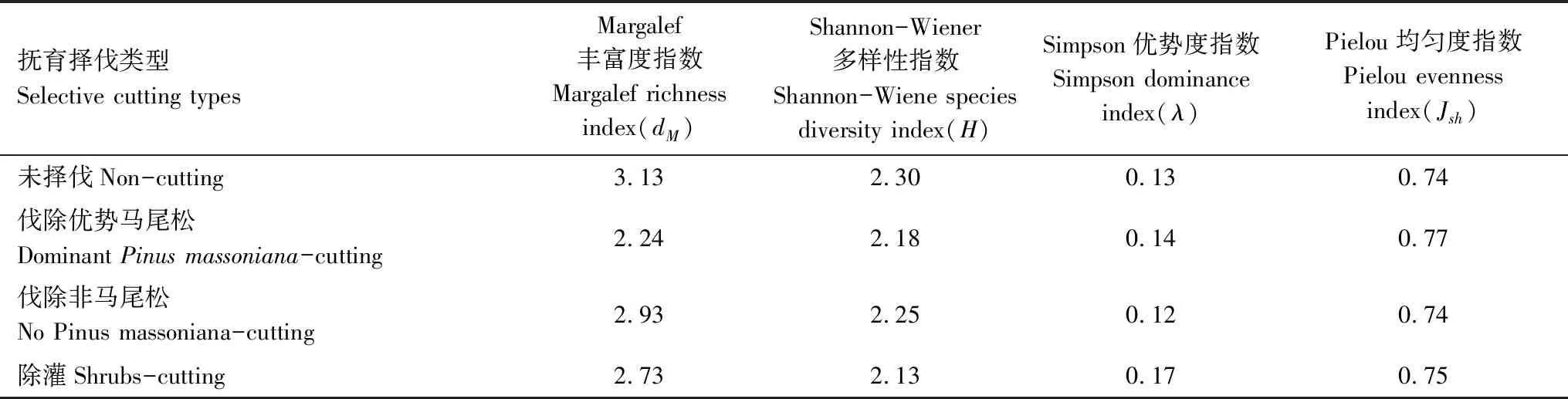
表3 撫育擇伐3 a后喬木樹種物種多樣性指數
3.2 撫育擇伐方式對馬尾松林主要樹種空間分布格局的影響
各樣地內種群分布點圖可以直觀反映出各樹種個體的空間分布狀況。不同撫育擇伐措施明顯改變了各優勢樹種的空間格局,且擇伐強度和擇伐對象選擇的不同對優勢樹種空間分布格局的影響程度也存在差異(圖1)。
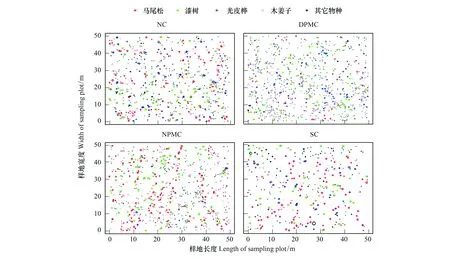
圖1 不同撫育擇伐樣地中主要喬木物種的空間分布散點圖Fig.1 Spatial distribution pattern of major tree species in different selective cutting after three yearsNC:未擇伐,Non-cutting;DPMC:伐除優勢馬尾松,Dominant Pinus massoniana-cutting;NPMC:伐除非馬尾松,No Pinus massoniana-cutting;SC:除灌,Shrubs-cutting
由圖2和圖3可知,在NC樣地中,馬尾松在5.5—6 m尺度上呈聚集分布,其他尺度表現為隨機分布;茅栗在0—12.5 m尺度上呈明顯的聚集分布;漆樹在0—7 m尺度上呈隨機分布,7—12.5 m尺度上表現為聚集分布;光皮樺在0—1.5 m尺度上呈聚集分布,>1.5 m的尺度上表現為隨機分布;宜昌木姜子僅在0—0.5 m尺度上呈聚集分布,>0.5 m的尺度上表現為隨機分布。在DMPC樣地中,馬尾松在0—4.5 m尺度上呈隨機分布,4.5—12.5 m尺度上為聚集分布;漆樹在0—0.5 m尺度和9—12.5 m尺度上呈隨機分布,而在0.5—9 m尺度上則表現為聚集分布;光皮樺、香椿、鹽膚木在0—12.5 m尺度上均表現為聚集分布;宜昌木姜子和檫木在>0.5 m的尺度上表現為聚集分布;火棘在0—1.5 m尺度上呈聚集分布,而>1.5 m尺度上表現為隨機分布。在NPMC樣地中,馬尾松在0—8 m尺度上呈聚集分布,8—12.5 m尺度上表現為隨機分布;漆樹在1—12.5 m尺度上呈為聚集分布;宜昌木姜子在0—9.5 m和11.5 m尺度上呈隨機分布,9.5—11.5 m尺度上表現為聚集分布;香椿在0—8 m尺度和10.5—11.5 m尺度上呈聚集分布;鹽膚木在0—12.5 m尺度上呈聚集分布;火棘在0—0.5 m和10.5—12.5 m尺度上呈聚集分布。在SC樣地中,馬尾松在0—2 m尺度上呈隨機分布,2—8 m和11—12.5 m尺度上表現為輕微聚集分布;漆樹和光皮樺在0—12.5 m上均表現為隨機分布。
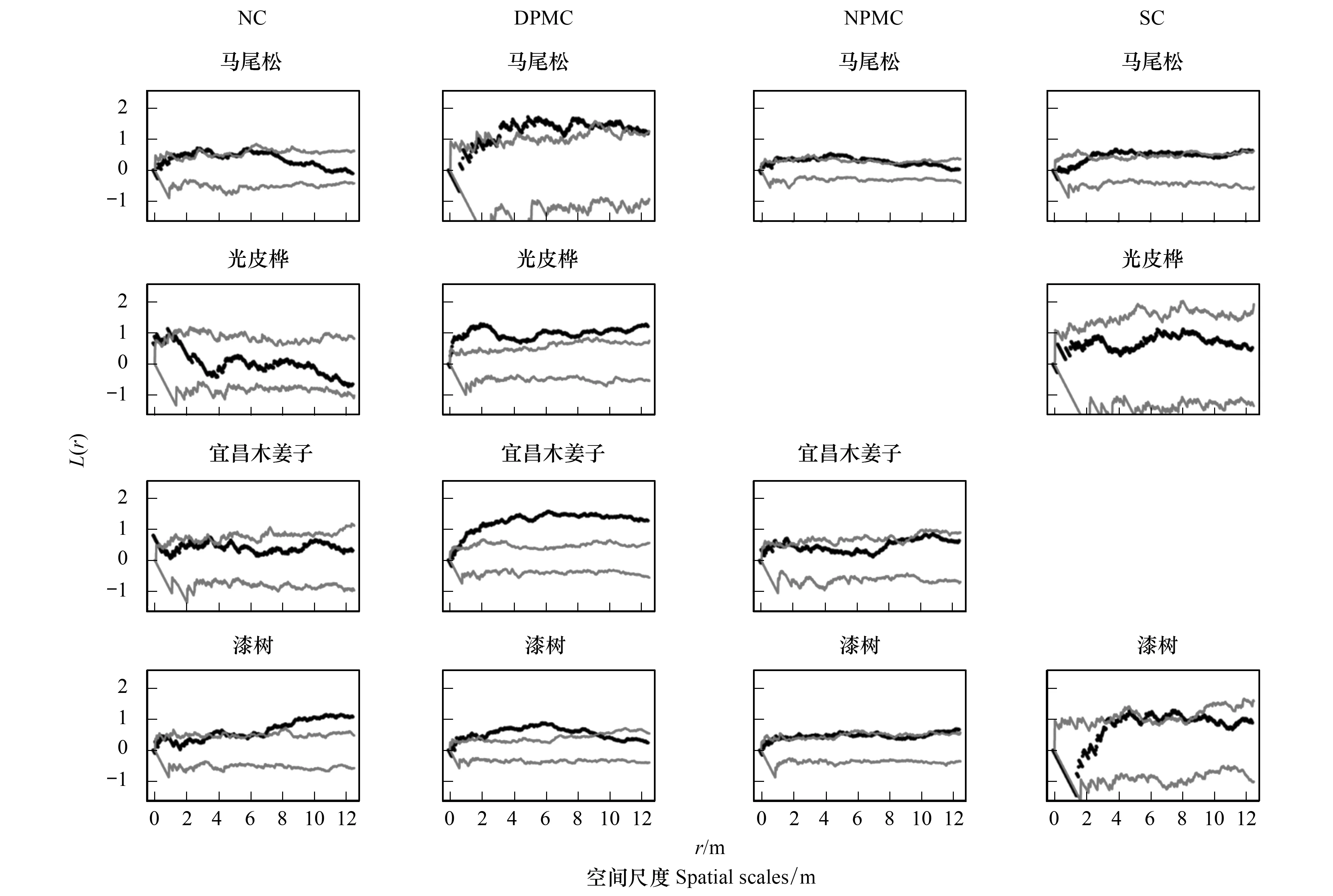
圖2 不同撫育擇伐3 a后馬尾松林4個優勢樹種的點格局分析Fig.2 Point pattern analysis of four dominant tree species in different selective cutting after three yearsNC:未擇伐,Non-cutting;DPMC:伐除優勢馬尾松,Dominant Pinus massoniana-cutting;NPMC:伐除非馬尾松,No Pinus massoniana-cutting;SC:除灌,Shrubs-cutting
比較不同撫育擇伐措施中主要樹種空間分布格局發現,NC樣地中馬尾松、漆樹、光皮樺、宜昌木姜子等主要樹種空間格局以隨機分布為主。DMPC促使馬尾松、光皮樺、香椿、鹽膚木和火棘呈現出明顯的聚集分布。NPMC導致光皮樺數量和地位嚴重下降,且香椿和鹽膚木主要以聚集分布格局為主,其他樹種空間分布格局與未擇伐樣地相似。SC進一步增加了林下更新闊葉樹種呈明顯的隨機分布格局。
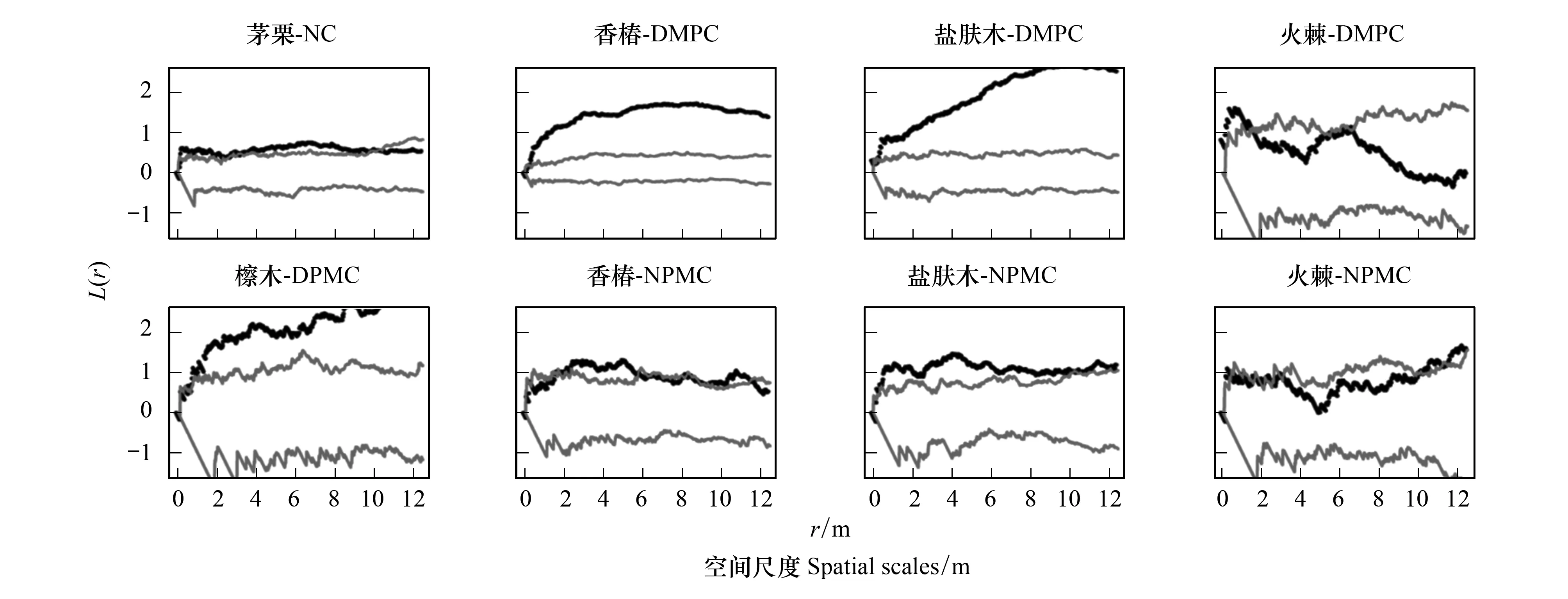
圖3 不同撫育擇伐3 a后其他優勢樹種點格局分析Fig.3 Point pattern analysis of other dominant tree species in different selective cutting after three years
3.3 撫育擇伐對馬尾松林主要樹種空間關聯性的影響
種間關聯性是用來表示一定階段內植物種群組成個體之間的空間相互關系和功能關系[25],優勢種間的關聯性可以反映其空間依賴性[18],分析研究人為撫育擇伐干擾后的主要樹種空間關聯性,有助于了解保留木在伐后群落中的作用。由圖4和圖5可以看出,在NC樣地中,馬尾松與漆樹、馬尾松與茅栗、光皮樺與漆樹在0—12.5 m尺度內無明顯空間相關性,馬尾松與光皮樺在5—11 m尺度上呈顯著正相關,宜昌木姜子與馬尾松、漆樹均在1—11 m尺度上呈顯著正相關,光皮樺與宜昌木姜子在4.5—11 m尺度上呈顯著正相關。在DPMC樣地中,馬尾松與漆樹在8—12.5 m尺度上呈顯著負相關,與光皮樺在2—9 m尺度上呈顯著負相關,與鹽膚木在2—8 m尺度上呈顯著負相關;與香椿在10—12 m尺度上呈顯著正相關;宜昌木姜子與馬尾松、漆樹在整個分析尺度上呈顯著正相關;光皮樺與漆樹在0—2.5 m尺度接近正相關,而在4—12.5 m尺度接近或達到空間負相關,與宜昌木姜子在1—9 m尺度上呈顯著負相關。在NPMC樣地中,馬尾松與漆樹在1—12.5 m尺度上接近空間正相關,與宜昌木姜子在0—5 m尺度上呈顯著正相關,與香椿在3—5.5 m尺度上呈顯著負相關;馬尾松與光皮樺、光皮樺與漆樹、宜昌木姜子均在0—12.5 m尺度內無明顯空間相關性,與鹽膚木在5—12.5 m尺度上呈顯著負相關;漆樹和宜昌木姜子在0—3.5 m尺度上呈顯著正相關。在SC樣地中,僅馬尾松與光皮樺在4—5.5 m和8—12.5 m尺度上呈接近或達到空間負相關,而其他主要樹種關聯性呈明顯的不相關。
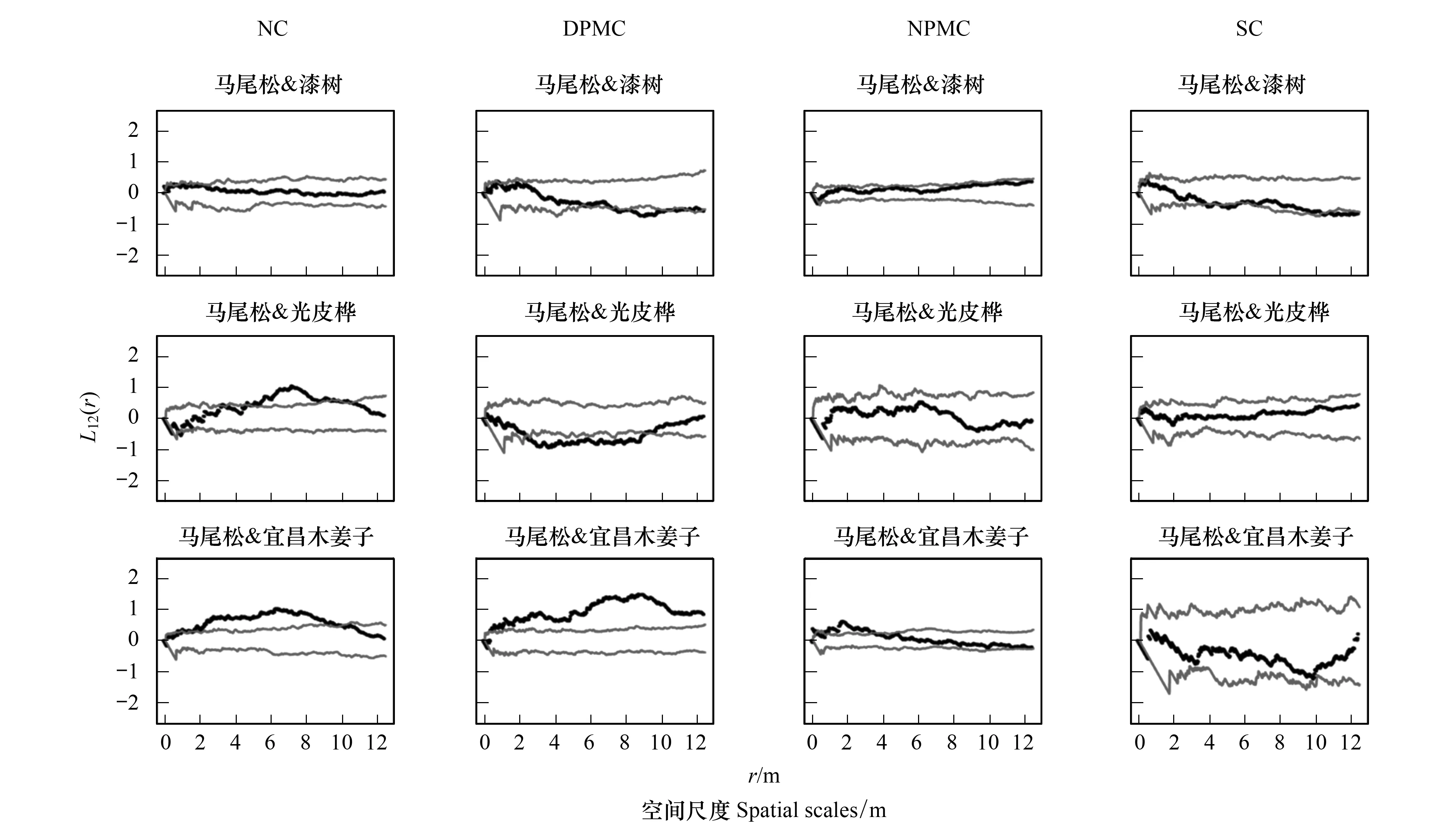
圖4 不同撫育擇伐3 a后馬尾松與其他3個優勢樹種間空間關聯性Fig.4 Interspecific spatial association between Pinus massoniana and other three dominant tree species in different selective cutting after three years
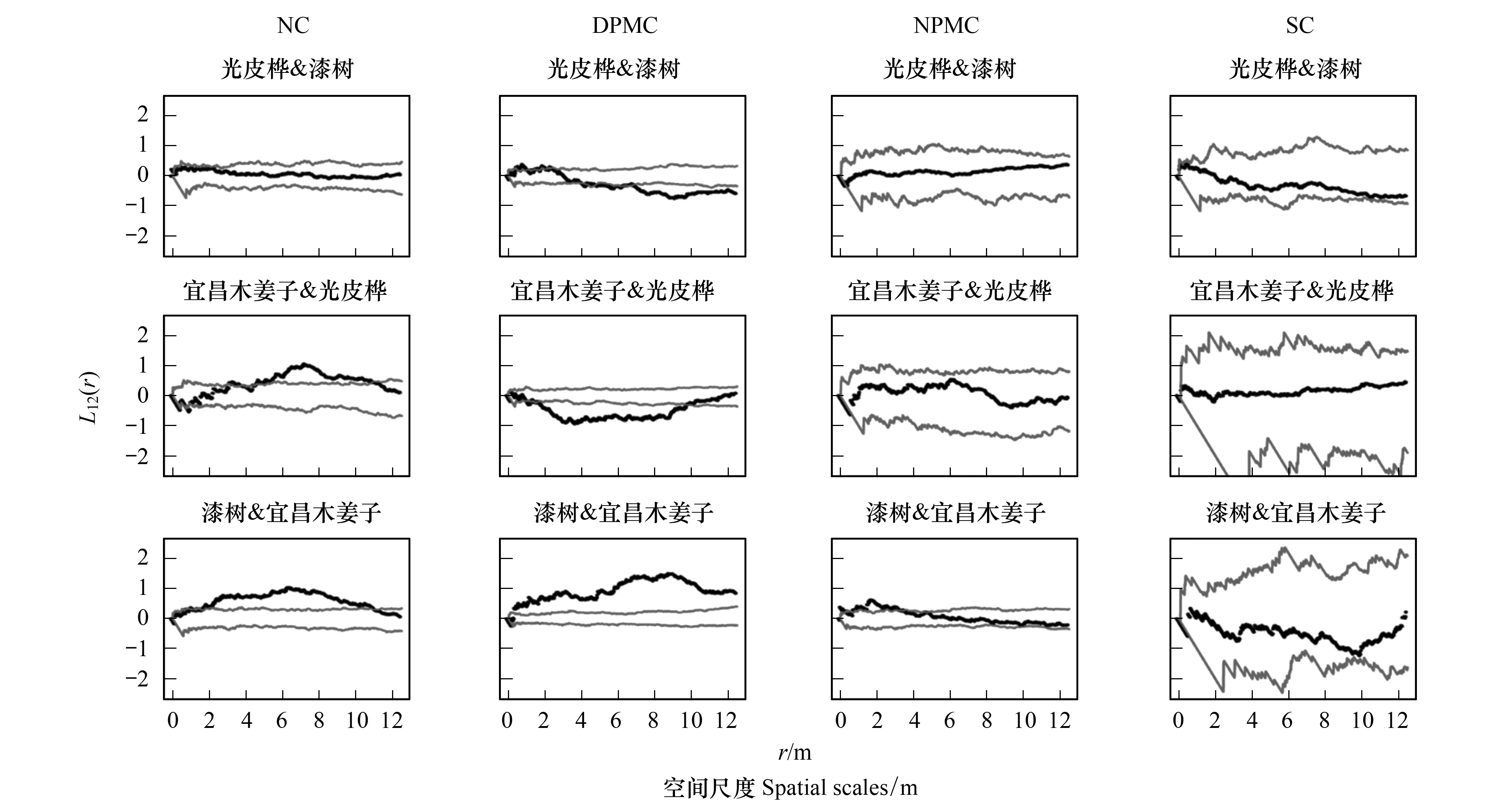
圖5 不同撫育擇伐3 a后其他優勢樹種間空間關聯性Fig.5 Interspecific spatial association of other dominant tree species in different selective cutting after three years
4 討論
4.1 擇伐對象差異對馬尾松林群落結構、物種空間關聯性的短期影響
種群分布格局受多種因素影響,如物種生物學特性、生境異質性、人為干擾因素等[1,25- 26]。撫育擇伐往往會形成光照強弱、林隙效應以及土壤營養條件不均勻等空間異質性,擇伐對象不同勢必會影響保留木的空間分布格局[11]。在本研究中,未擇伐馬尾松林分的主要樹種空間格局以隨機分布為主,優勢樹種間主要以不相關和正相關為主,說明該階段的林分樹木已逐漸占據了有效的空間生態位,種間相互競爭和排斥作用較弱,林木空間分布格局已具備一般天然林分個體隨機分布的特征[27]。然而,伐除優勢馬尾松反而明顯促使優勢樹種呈聚集分布,且樹種間的負相關性明顯增加,這是由于伐除優勢馬尾松影響了建群種和上林層結構,林冠大開導致林下更新樹種為了爭奪更多的光照、水分、土壤、養分等生存條件,加劇了樹種間的競爭性。伐除非馬尾松措施主要伐除個體較大的更新闊葉樹種,造成林內環境局部異質性增強[19],導致漆樹、香椿、鹽膚木等目標擇伐樹種呈明顯的聚集分布,保留主要樹種間以不相關為主,在一定程度上降低了種間正相關性而呈現不相關,降低了林分的穩定性。除灌措施主要去除林下植被層的灌木物種,減少了林下層植被與更新樹種的生存環境競爭,促使更新闊葉樹種隨機分布程度增強,致使更新樹種間的生長空間擴大,有利于林下樹種的生長發育[28- 29]。另外,已有研究發現,處于演替初期的群落中物種相關性主要以負相關和不相關為主,隨著植被演替的進行,物種間負相關性逐漸減弱,正相關性逐漸增強,群落結構及物種組成也更加趨于穩定[2,7]。可見,撫育擇伐導致林分發生逆向演替且對林分結構產生的影響各不相同,可能主要由于不同物種或不同層次結構的物種在群落中的生物學特性、地位、作用、相互關系以及對外界干擾響應能力存在差異所產生的。
4.2 撫育擇伐強度對馬尾松林群落結構、物種空間關聯性的短期影響
擇伐強度的大小嚴重制約了伐后林分的生長狀況[15],適度擇伐有利于優化林分空間結構,增強林分功能和穩定性[12,30]。本研究中伐除優勢馬尾松(擇伐強度為70%)屬于強度擇伐,對林分擾動過大造成林內樹種的聚集程度明顯增加,可能是由于強度擇伐降低了上林層覆蓋,造成了大面積的林窗,林窗內充足的資源促進了各個樹種保留木和幼樹的生長,但同時樹種競爭加劇又會制約優勢樹種的生長[28]。伐除非馬尾松(擇伐強度15%)屬于中度擇伐,導致部分樹種在大尺度上呈現聚集分布,種間關聯性以不相關為主,可能是緣于該措施產生了有限的林窗,促使某些樹種呈聚集化有利于發揮功能群體效應,利用有限的空間資源[10],形成適合于自身生長的微環境,從而抵御外部競爭[5],而且伐后林分結構可在短期內恢復,導致許多優勢樹種分布格局區域隨機或輕微聚集分布。雖然本研究未統計除灌措施的擇伐強度,但由于林下層植被在森林總生物量中只占很少的一部分,屬輕度擇伐。除灌進一步增強了各優勢樹種空間格局的隨機分布以及樹種空間關聯性的不相關性。短期去除林下植被層暫時有利于加速林下更新速度,但長期來看,去除林下植被對上層喬木樹種的生長不存在影響[31],低強度擇伐不能改變喬木樹種對于絕大部分資源的競爭優勢,對林分結構優化改變的程度有限。同時,森林群落組成和結構對撫育擇伐活動的響應也存在一定的時間延遲[13- 14],短期擇伐并未加速林分組成朝著演替后期的方向發展[11],未來隨著演替時間的發生,撫育擇伐后林分的群落組成結構將會進一步產生變化[7,32]。
綜上所述,伐除優勢馬尾松、伐除非馬尾松和除灌等撫育擇伐措施均不同程度的改變了林木組成、空間關系、競爭狀態。單純調整某個樹種或單個層次的結構來開展森林經營,不利于林分空間結構的整體優化。因此,未來針對三峽庫區馬尾松林經營管理時,首先要基于林分空間格局和種間關聯性判斷林分是否需要人為經營手段的干預,盡量減少人為干擾的程度和次數。其次,擇伐對象要優先選擇清理衰老樹、枯死木、生長勢較弱的馬尾松個體以及更新樹種聚集分布區域和灌木層,盡量保留有利于提高林分混交度的樹種。再次,遵循種群結構和空間格局分布規律,保證林分地表的連續覆蓋,擇伐強度應控制在15%左右,防止林分結構劇烈變化,通過產生適度的林窗,改善林分生長環境,促進馬尾松林向針闊混異齡林發展,進而實現森林結構優化和功能提升。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
趣味(語文)(2020年3期)2020-07-27 01:42:46
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
