臨界脅迫貯藏條件對水稻種子活力的影響
2019-07-25 08:22:04曹高燚華澤田
湖北農業科學 2019年13期
蔣 薇 ,李 瑤 ,李 娜 ,趙 飛 ,,亓 娜 ,蔡 卓 ,曹高燚 ,,華澤田
(1.天津農學院農學與資源環境學院,天津 300384;2.中國農業科學院甘薯研究所,江蘇徐州 221000;3.國家粳稻工程技術研究中心,天津 300357)
種子是農業的根本,種子質量的高低與農業的發展息息相關。種子活力(Seed vigor)是衡量種子在田間出苗情況的重要指標,實驗室可以通過監測種子發芽勢、發芽率、發芽指數及活力指數來評價種子活力狀態。種子萌發由遺傳和環境條件等多種因素調節,環境條件是制約水稻種子活力的重要因素,其中包括收獲時期、溫度、種子含水量等[1]。張玉蘭等[2]以粳稻和秈稻種子為試驗材料,研究5年后種子不同含水量對貯藏壽命的影響,表明水稻種子適度超干的含水量對種子活力的影響最小,最利于種子儲藏。 在此基礎上,胡群文等[3,4]進一步研究得出,若含水量保持在5.5%左右,種子保存最為得當,即此時的種子活力相較其他含水量種子最高。陳新紅等[5]以當地水稻品種為試材,研究得出逆境不利于種子的保存,且逆境條件對種子的損害越大,種子活力下降得越明顯。與此同時,種子發育得越完整,結構越齊全,種子萌發得越好。徐軍等[6]的研究表明在種子完熟期之后收獲,對種子活力越有利。關于貯藏溫度對種子活力的影響,繆麗霞等[7]采用8個水稻品種探討得出貯藏溫度越低,對種子發芽力影響越小。低溫和適宜含水量能較好地降低種子貯藏過程中活力的損耗。
水稻是產業化程度較高的作物,每年都面臨較大的貯藏損失,影響種子質量。種子貯藏時間的長短往往受內部和外部兩個方面的因素影響,外部因素主要是儲藏環境和儲藏時間,內部因素常以含水量衡量。
本研究設置了不同的儲藏條件對不同品種水稻種子進行整體活力研究,探求水稻保存的極限條件,了解水稻種子耐貯性生理生化機制,明確不同含水量、貯藏溫度與種子活力的關系,對不同品種水稻種子的耐貯性做出客觀評價,確定最優的雜交粳稻種子貯藏環境,為水稻種子耐貯性生理生化機制提供理論依據,對降低企業的貯藏成本和生產風險具有重要應用價值。
1 材料與方法
1.1 試驗材料
2個常規粳稻品種津原E5和津原11。
1.2 試驗方法
1.2.1 種子貯藏水分設置 將水稻種子吸水晾干,實時監測種子含水量,再配合硅膠干燥劑,將符合試驗要求的種子塑封后放入不同溫度的冰箱貯藏。津原 E5設低含水量(9.6%)、中含水量(14.8%)、高含水量(25.1%)3個處理;津原11設低含水量(9.6%)、中含水量(14.3%)、高含水量(22.7%)3個處理。
1.2.2 種子貯藏溫度設置 試驗設置3種不同貯藏溫度,分別為4、-10和-20℃ 3個處理,分別在貯藏1個月(4月份)、6個月(9月份)及9個月(12月份)時測定其種子活力指標。
1.2.3 鑒定水稻種子活力相關參數 水稻種子可溶性蛋白含量、丙二醛含量、過氧化物酶活性參照成廣雷等[8]的方法測定。
2 結果與分析
2.1 種子發芽指數的變化
種子發芽指數是種子發芽率的細化和深入,是種子活力指標的重要參數。津原E5和津原11兩個品種從4—12月貯藏期間發芽指數見圖1,并未達到顯著性變化。津原E5在12月時,不同貯藏條件下發芽指數呈現一定的趨勢(圖1A),在4℃時,隨著種子含水量的下降,發芽指數略微上升。在-10℃時,種子發芽指數與種子含水量的關系更為密切。在-20℃時,可明顯地觀察到隨著種子含水量的下降,發芽指數大幅度上升。在-20℃時,貯藏至12月時津原E5(含水量25.1%)和津原11(含水量22.7%)的高含水量種子較貯藏初期(4月)種子發芽指數呈明顯下降趨勢。在低溫(4℃)及超低溫(-10、-20℃)貯藏條件下,較低的種子含水量能在一定程度上保護種子,有助于種子的萌發。
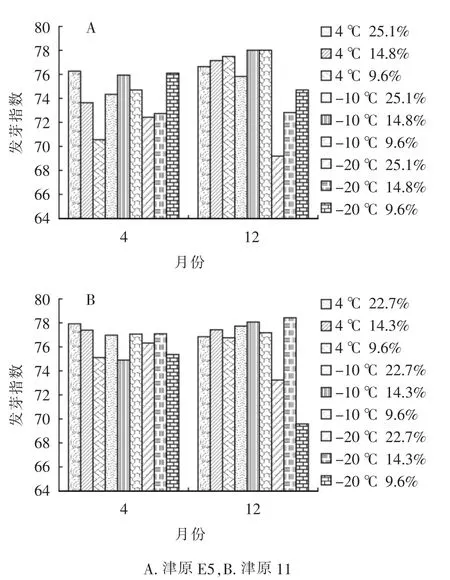
圖1 種子貯藏過程中發芽指數變化
2.2 種子活力指數的變化
種子活力指數是種子發芽速率和生長量的綜合反應,可以更準確地反映種子活力。津原E5和津原11兩個品種的活力指數,在貯藏過程中無顯著性變化。低含水量處理的種子在-20℃相較于其他含水量種子有較高的活力指數(圖2)。由此可見,在較短的貯藏時間內,種子活力指數基本不變,同時,在超低溫(-20℃)貯藏條件下,較低的種子含水量有助于種子的萌發,且溫度越低(-20℃)效果越顯著。

圖2 種子貯藏過程中活力指數變化
2.3 可溶性蛋白含量的變化
種子可溶性蛋白參與各種代謝,可以反映種子的代謝狀況。經過9個月的臨界貯藏,兩個品種的可溶性蛋白含量均有所下降。種子在-20℃貯藏期間,中含水量和低含水量種子可溶性蛋白變化不顯著,但并不改變整體的下降趨勢。可溶性蛋白含量與貯藏時間呈負相關,且高含水量種子下降最顯著(圖3)。

圖3 種子貯藏過程中可溶性蛋白含量變化
2.4 丙二醛含量的變化
丙二醛是植物體內脂質過氧化的有害代謝物,它能與細胞各成分發生強烈反應的過氧化產物,會損傷多種酶和膜系統。測定丙二醛(MDA)含量可以反映種子脂質過氧化的程度。隨著貯藏時間的增加,MDA含量不斷積累,津原E5在貯藏期間,中含水量和低含水量處理的種子MDA含量均顯著上升,且在不同貯藏溫度下,隨含水量的下降,MDA含量上升(圖4)。津原11在貯藏期間,4℃和-10℃處理的種子MDA含量隨種子含水量下降而逐漸上升。臨界貯藏條件下種子MDA含量隨含水量下降而上升。
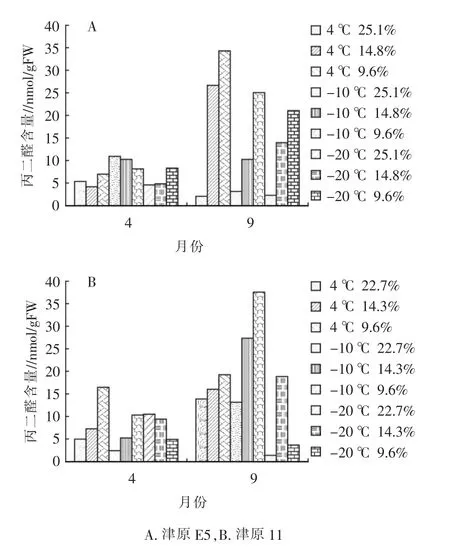
圖4 種子貯藏過程中丙二醛含量變化
3 小結與討論
種子萌發是個多因素作用的復雜過程,始于吸脹止于胚根萌發,是植物生長的第一步[1]。種子萌發受到內部因素和外界環境的共同調控,是胚胎生長發育的必要條件[9-11]。種子成功的發芽不僅對成苗至關重要,而且對作物產量也很重要。比較擬南芥、水稻和玉米等不同植物種子萌發的作用機制,其在形態學變化、細胞及其相關結構恢復、代謝激活、激素行為、轉錄和翻譯激活等都存在異同[12]。控制種子萌發和休眠的因素很多,包括由植物和土壤細菌產生的植物激素[13]。植物激素與植物基因的相互作用影響種子萌發。雖然植物激素的活性受不同水平基因表達的控制,但有一些植物基因在特定植物激素存在時被激活[14,15]。因此,調節基因表達可能是促進種子萌發的有效途徑。
除了分子機制和激素,外部環境因素如溫度光照亦影響著種子萌發[16]。在19世紀早期,人們就發現紅光能促進某些種子(如萵苣種子)萌發,而遠紅光抑制其萌發[17-19]。同樣,溫度對種子萌發和休眠也起著重要的調控作用[20]。不同植物的萌發對溫度的要求不同,低溫往往適于種子萌發,因為種子打破休眠往往需要低溫。有些植物萌發則需要一個變溫的過程[21,22]。
貯藏條件如溫度、時間、種子含水量等均對種子活力及其壽命有重要影響。張苗苗等[23]通過加拿大披堿草貯藏試驗指出,隨貯藏時間延長,種子發芽指數呈拋物線狀變化,剛收獲的種子在貯藏1~2年時利用價值較佳。由于每個品種收獲時期不甚相同,貯藏時間存在差異,導致種子休眠期存在一定程度的差異,同時不同品種水稻種子的休眠期也會存在差異,這就導致每個品種間的發芽指數、活力指數存在一定差異。同時也不排除在超低溫貯藏過程中種子發生遺傳變異而導致生理出現差異。周建明等[24]通過雜交粳稻常優3號貯藏試驗表明,種子在低溫貯藏8個月時活力指數開始下降,12個月達到極顯著。
隨著貯藏時間的延長,種子活力開始下降,導致蛋白含量下降[25-27]。丙二醛產生的根源是膜脂過氧化,主要指脂肪的過氧化,即脂肪酸在不飽和脂肪酸雙鍵上發生過氧化作用,從而使細胞膜功能衰竭。種子內油脂在酸敗過程中易產生多種有害物質,即包括丙二醛,對種子活力的降低有很大影響[28,29]。活性氧(ROS)相關線粒體功能障礙在種子退化中起著重要作用。ROS的大量產生誘導線粒體特異蛋白的改變和修飾,這些蛋白可能參與線粒體退化的過程,最終導致種子活力喪失[30,31]。 種子能否成功萌發在很大程度上取決于種子的氧化穩態。過氧化氫還原酶(APX6)在種子萌發控制和種子抗逆性方面具有新的作用。APX6替代APX1作為干種子中的主要APX,其功能喪失導致種子萌發因活性氧過度積累和氧化損傷而降低。由此可以看出種子通常會通過保護酶來降低ROS對細胞的損傷,從而在一定程度上維持種子活力[32]。本研究中丙二醛在貯藏過程中呈上升趨勢,而過氧化物酶(POD)活性呈下降趨勢,與前人研究結果相一致。
研究發現,相關活力指標如發芽指數、活力指數與貯藏時間無明顯線性關系,同時可溶性蛋白、丙二醛含量與貯藏時間呈線性關系;其中,可溶性蛋白含量與貯藏時間呈負相關,丙二醛則反之。
種子貯藏時的溫度和含水量與種子活力存在一定關聯。當溫度一定時,發芽指數隨種子含水量的降低而增加。可溶性蛋白在超低溫高含水量情況下受到明顯破壞,含量最低。在不同貯藏溫度下,隨含水量下降,丙二醛含量上升。POD活性在超低溫高水分條件下最低。
